
Abstract
Parasitic granulomatous eosinophilic inflammation was observed in the central nervous system (CNS) of a 6-month-old Arabian colt from New York state. Inflammation was associated with eggs, larvae, and adult nematodes in the cerebellum. Nematodes had histological characteristics of the superfamily Metastrongyloidea. The presence of dorsal-spined larvae in the CNS was further indicative of infection with a nematode in the family Protostrongylidae. Infections were most compatible with Parelaphostrongylus tenuis but specific diagnosis was not possible. This is the first definitive report of a protostrongylid nematode infection in a horse.
Keywords
Parasitic migratory encephalomyelitis is a rare but important cause of neurologic disease in horses. Metazoan parasites identified from the equine central nervous system (CNS) include nematodes (Strongylus vulgaris, S. equinus, Angiostrongylus cantonensis, Halicephalobus gingivalis, Setaria spp., and Draschia megastoma) and fly larvae (Hypoderma spp.). 9 We identified parasitic nematodes in the CNS of a young horse in New York state. Nematode morphology and its occupation of the CNS were most compatible with the meningeal worm, Parelaphostrongylus tenuis. The northeastern USA has a high density of white-tailed deer (Odocoileus virginianus) that have a high prevalence of P. tenuis infection. This neurotropic nematode causes a debilitating neurologic disease in cervids, 1, 6 ovids, 1 bovids, 1, 5, 12 and camelids. 1 Recently, Van Biervliet et al. identified a Metastrongyloidea nematode in the CNS of a horse, 11 but specific identification of the parasite was not possible. Here we present the first definitive report of protostrongylid nematodes in the CNS of a horse.
A 6-month-old male Arabian horse had acute onset of an abnormal gait and held its head to the right. There was no history of pyrexia or systemic illness. The colt was born in New York state and had not traveled to other states. Clinical signs persisted following treatment with dexamethasone, and the horse was referred to the Cornell University Hospital for Animals. Clinical examination revealed marked spastic tetraparesis and ataxia in all four limbs. The head and neck were held to the right side. A C1 to C5 lesion was suspected. Cerebrospinal fluid contained mildly increased protein (116 mg/dl, normal range 5–100 mg/dl) and 1200 cells/μl (58% lymphocytes, 40% neutrophils, 2% macrophages, and a few eosinophils and erythrocytes). Cervical radiographs were suspicious for subluxation at the atlanto-occipital joint. Computed tomography (CT) examinations were normal except for a mild atlanto-occipital joint subluxation, and a light weight cast was applied as treatment for the subluxation. The foal was unable to stand on recovery from anesthesia for the CT scan and struggled violently to attempt to stand. The patient was humanely euthanized and a necropsy was performed.
Necropsy examination revealed mild subluxation of the atlanto-occipital joint on the left side with elongation of the left alar ligament. There was considerable edema of the epidural fat surrounding C1, but no hemorrhage or discoloration was identified. The calvaria and cervical vertebrae were grossly normal, and no malformation was identified. No other gross abnormalities were recognized in other organs. The entire brain and segments of spinal cord were fixed in 10% neutral-buffered formalin. On sectioning of the fixed brain, a mild, diffuse, dull discoloration of the cerebellar parenchyma was observed (Fig. 1). This was most prominent in the deeper aspect of the cerebellar vermis (Fig. 1). Tissues were embedded in paraffin, sectioned, and stained with HE.
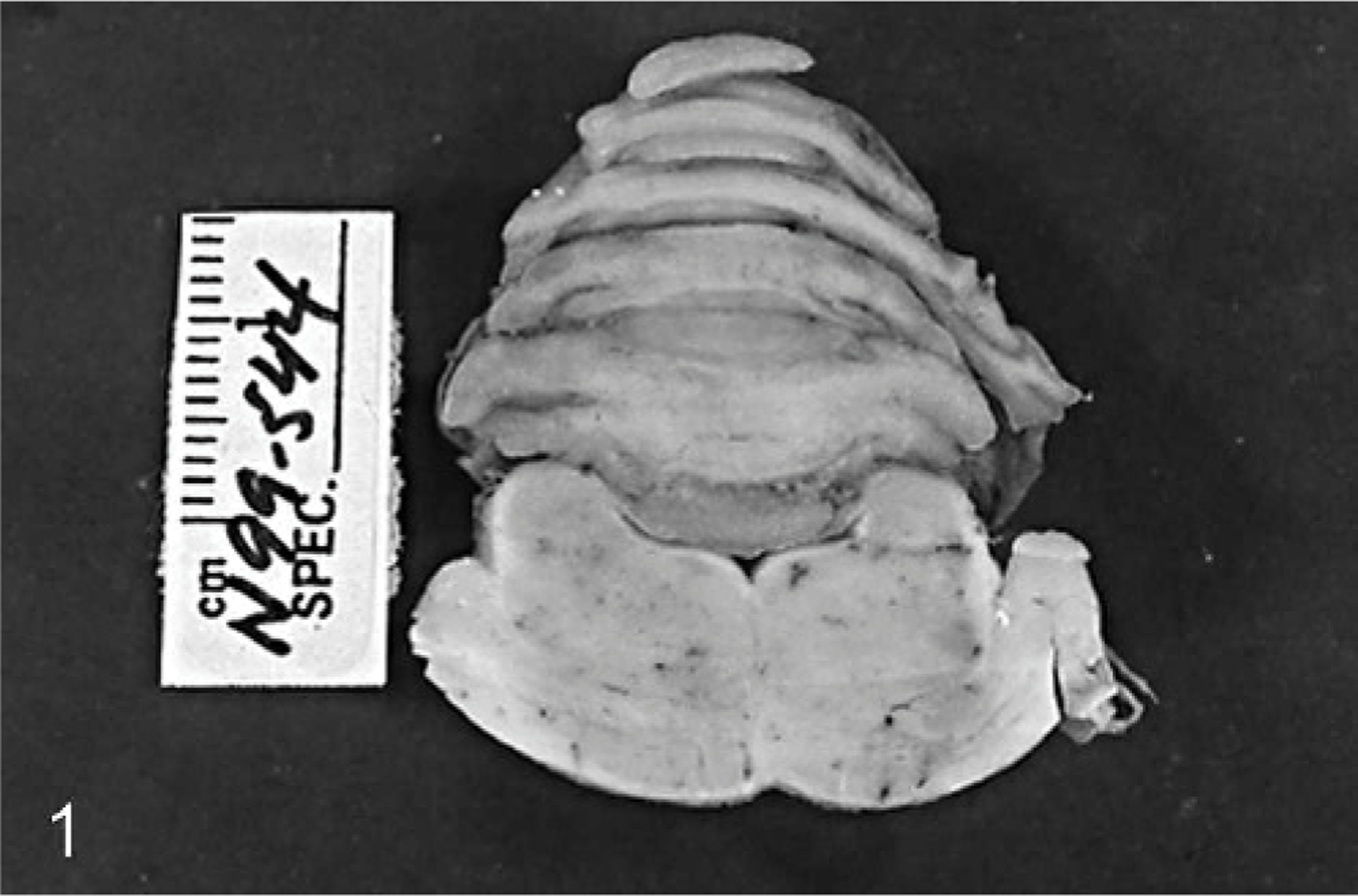
Cerebellum and pons; horse. Dull discoloration of cerebellar parenchyma.
Microscopically, the cerebellar parenchyma and the associated meninges were widely replaced by multifocal to coalescing eosinophilic granulomatous inflammation. This was most severe in the parenchyma of the cerebellar vermis and obliterated cerebellar cortex, white matter, and meninges. The exudate was composed of large numbers of epithelioid macrophages, fewer eosinophils, and multinucleated giant cells accompanying moderate astrogliosis. The affected neuropile was edematous and pale. The white matter had scattered swollen axons. In the adjacent, less affected cerebellar parenchyma, there was moderate perivascular lymphocytic and eosinophilic cuffings, expanding the Virchow-Robin spaces. The meninges were thickened with edema and lymphocytic and eosinophilic infiltration. Numerous eggs and nematode larvae were present within inflammatory exudates (Fig. 2). The eggs were approximately 21 μm in length and had a thin shell. Both developing and embryonated eggs were present (Fig. 2). The first-stage larvae (L1) were approximately 15 μm in diameter and had thin lateral alae. A posteriorly directed dorsal spine was observed anterior of the crooked tail on L1 (Fig. 3). The full length of L1 could not be determined from histologic sections. At least 1 adult female worm was present in the cerebellar parenchyma and meninges (Fig. 4). One male worm was identified in the meninges over the medulla oblongata (Fig. 5). Adult parasites had a round body with a smooth cuticle, polymyarian-coelomyarian musculature, lateral and accessory hypodermal cords, and a “strongylid” intestine composed of 3 to 5 multinucleated cells with indistinct microvilli (Fig. 4 and inset). The female parasite was 173 μm wide with paired genital tracts containing eggs and sperm (Fig. 4 inset). The male parasite was 161 μm wide and had a single genital tract containing sperm (Fig. 5).

Cerebellum; horse. Granulomatous encephalitis with numerous developing eggs (arrow) and a few embryonated eggs (arrowhead). HE. Scale bar = 100 μm.

Cerebellum; horse. First-stage larva with crooked tail and a posteriorly directed spine (arrowhead). HE. Scale bar = 10 μm.
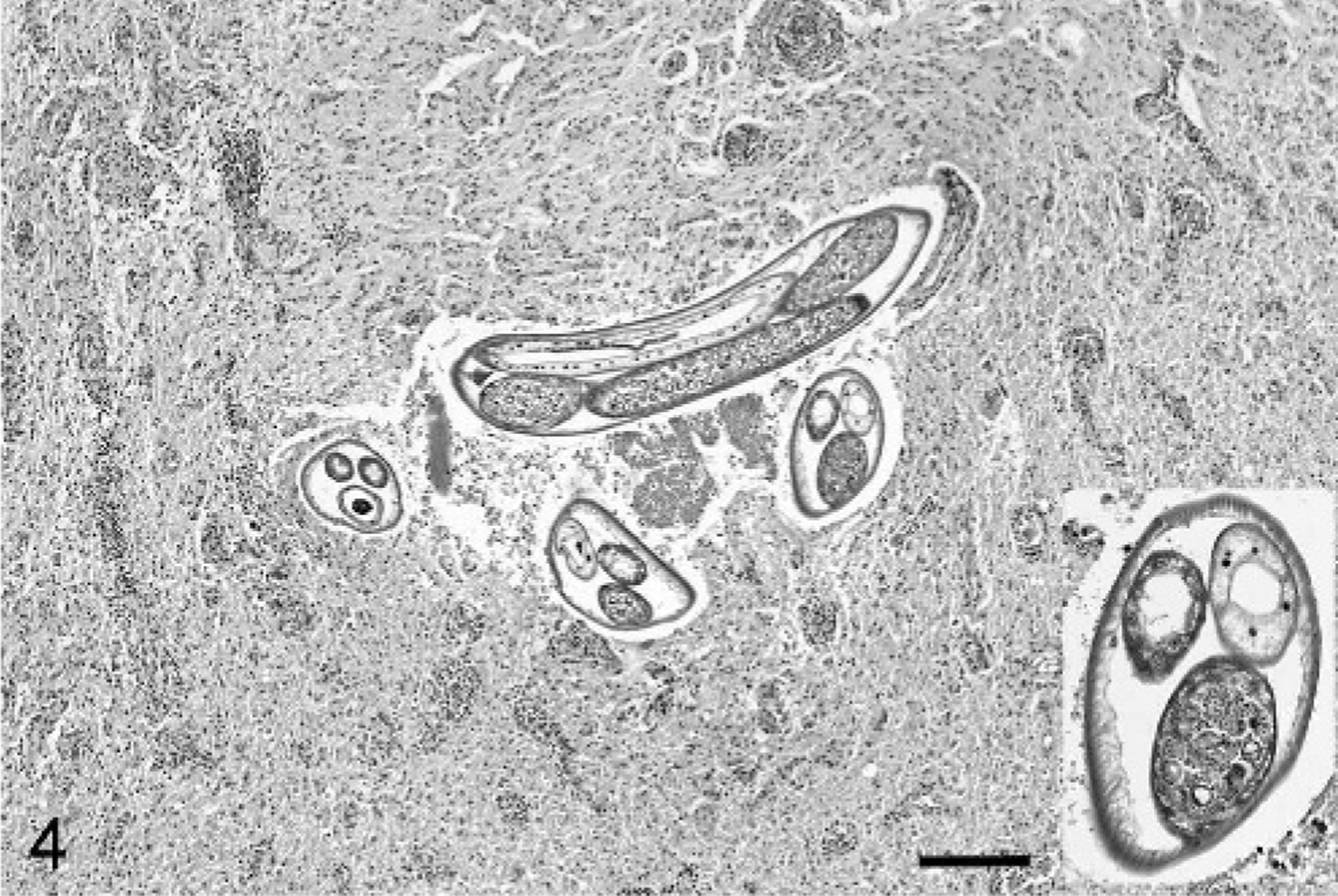
Cerebellum; horse. Granulomatous encephalitis with cross and longitudinal profiles of an adult female nematode. Inset shows higher magnification of a cross section of the worm. Note eggs and sperm in genital tracts. HE. Scale bar= 250 μm.

Medulla oblongata; horse. A cross section of male parasite in the meninges. Asterisk indicates parenchyma of medulla oblongata. HE. Scale bar = 100 μm.
Nematodes from the colt and from a previous horse infection reported by Van Biervliet et al. 11 had the characteristic “strongylid” intestine and polymyarian-coelomyarian musculature compatible with nematodes in the superfamily Metastrongyloidea. 3 Within this group, only species in the families Protostrongylidae and Angiostrongylidae demonstrate neurotropism and infection of the CNS. 1 The L1 in the CNS of the colt had lateral alae and crooked tails with a posteriorly directed dorsal spine. The spine is of diagnostic significance and is unique to protostrongylid nematodes. 1, 2 The family Protostrongylidae parasitize ruminants predominantly, and none of these species is known to infect horses. 1 This case is the first definitive report of protostrongylid nematode infection of a horse.
There are seven genera within the family Protostrongylidae that produce dorsal-spined larvae. Of these, species from both Parelaphostrongylus and Elaphostrongylus are known to infest the host CNS and musculature, whereas the other five genera occupy host lung exclusively. 1 In this case, infection with Elaphostrongylus is considered unlikely because this genus is not known to occur in North America. 8 The genus Parelaphostrongylus contains 3 species. Parelaphostrongylus odocoilei has been reported to occur in the musculature of mule deer and woodland caribou; its range is restricted to western North America. Parelaphostrongylus andersoni occurs in the musculature of white-tailed deer in the southeastern USA, New Jersey, Michigan, Wyoming, and southcentral British Columbia, and in barren-ground caribou throughout Canada. 1 Parelaphostrongylus tenuis is a common parasite of the CNS of white-tailed deer throughout eastern North America. 1 The width of the female worm in the colt CNS (173 μm) was compatible with the range of measurement reported for P. tenuis (120–250 μm) and P. odocoilei (141–179 μm) but not P. andersoni (92–130 μm). 6 The width of the male worm (161 μm) was compatible with the range of measurement reported for P. tenuis (92–200 μm) but not with P. odocoilei (138–156 μm) or P. andersoni (87–140 μm). 6 Definitive classification of protostrongylid nematodes can be made only following examination of reproductive structures from male worms. Isolation of adult nematodes from formalin-fixed tissues was unsuccessful. The etiologic diagnosis of P. tenuis, however, is supported in this case by nematode morphometry and tissue tropism, characteristic protostrongylid epizootiology, and the host's restricted travel history and geographic range.
Acquired scoliosis and dorsal gray column myelitis were observed in the previous horse with a larva of the superfamily Metastrongyloidea, and parasitic migration was proposed pathogenesis. 11 In the current case, the colt did not show clinical signs of scoliosis, and parasitic migratory lesion was not observed in the spinal cord macroscopically and microscopically.
Parelaphostrongylus tenuis, also called meningeal worm or brain worm, is a common neurotropic parasitic nematode of white-tailed deer throughout eastern North America. 1 Adult worms are found in the cranial subarachnoid space and in surrounding cranial venous sinuses. Eggs are deposited into the venous circulation and are carried as emboli to the lungs, where they lodge in small capillaries and become incorporated into granulomas. 1, 6 The eggs hatch and release L1 that escape from granulomas, penetrate lung alveoli, and enter air ducts. 1, 6 They are carried up the mucociliary escalator, are swallowed, and pass out in host feces. The L1 infect terrestrial mollusks and develop to the infective stage (L3) in this host. Deer and other hosts become infected by accidental ingestion of L3, presumably within mollusks ingested accidentally along with vegetation. 6 The L3 penetrate the abdomen and migrate to the spinal cord by 10 days after infection. 6 The larvae develop in the dorsal gray columns of the spinal cord, then migrate from the nerve parenchyma into the subarachnoid space of the cranium where they mature and copulate. 6
Parelaphostrongylus tenuis is generally clinically silent in white-tailed deer but causes neurological disease in all other North American cervids. 1, 7 Noncervid ungulates are also susceptible to P. tenuis. Debilitating neurologic disease has been reported in sheep, goats, llamas, bighorn sheep, and various types of antelopes. 1 Cattle are thought to be largely resistant to infection, 7 with disease reported only rarely. 5, 12 Horses might be similarly resistant because parasitic neurologic disease is rare in spite of extensive pasture overlap with infected white-tailed deer. For example, prevalence of P. tenuis in white-tailed deer is reported to be as high as 86%, 1 and would presumably provide ample opportunities for parasite exposure. The low incidence of infection in horses likely reflects the evolution of protostrongylid nematodes in ungulate hosts. The mechanism of resistance to P. tenuis infection in horses is unknown. It may be caused by differences associated with acidic pH around 6 of a simple stomach of grazing horses, differences in grazing habits that prohibit ingestion of L3, or differences in the horse immune response. 11 It is typically regarded that less compatible hosts result in poor or failed reproduction by P. tenuis. Excluding white-tailed deer, L1 have been observed in feces from some non-white-tailed cervids 4, 10 but not from ovids, bovids, or camelids. 7 Neither eggs nor larvae were observed in the lung of the colt in the present study. Feces were not examined for L1. Eggs occasionally are deposited on the meninges in permissive cervid hosts and hatch to release L1. However, neither eggs nor L1 have been reported from the CNS of infected ovids, camelids, or bovids. 7 Presence of protostrongylid L1 in the colt CNS is an unprecedented observation and indicates successful reproduction. It is possible that the young age of the animal played a role in establishment of infection and disease.
In summary, our observations confirm that horses are susceptible to encephalitis from infection by nematodes in the family Protostrongylidae. Specific identification of the nematodes was not possible but features of parasite morphology, their anatomical location, and epizootiology were most compatible with P. tenuis. Our observations suggest that horses might be susceptible to infection and disease caused by P. tenuis. Protostrongylid nematode infection should be included as a differential diagnosis for instance of neurologic disease in horses in endemic area of eastern North America.
Footnotes
Acknowledgements
The authors thank Alexis Wenski-Roberts and David M. Fitzgerald for assistance with photographs. We also appreciate the revision of this manuscript by Dr. Cynthia X. Bacmeister.