
Abstract
Nocardia spp. infections in mammals cause pyogranulomatous lesions in a variety of organs, most typically the lung. Members of the Nocardia asteroides complex are the most frequently recognized pathogens. Nine cases of nocardiosis in free-ranging pinnipeds and 10 cases of nocardiosis in cetaceans were evaluated. Host species included the hooded seal (Cystophora cristata, n = 8), leopard seal (Hydrurga leptonyx, n = 1), Atlantic bottlenose dolphin (Tursiops truncatus, n = 4), beluga whale (Delphinapterus leucas, n = 4), and killer whale (Orcinus orca, n = 2). The most common presentation of nocardiosis in both pinnipeds and cetaceans was the systemic form, involving 2 or more organs. Organs most frequently affected were lung and thoracic lymph nodes in 7 of 9 cases in pinnipeds and 8 of 10 cases in cetaceans. Molecular identification and bacterial isolation demonstrated a variety of pathogenic species. N. asteroides, N. farcinica, N. brasiliensis, and N. otitisdiscaviarum are pathogenic for pinnipeds. In cetaceans N. asteroides, N. farcinica, N. brasiliensis, N. cyriacigeorgica, and N. levis are pathogenic. Hematoxylin and eosin and acid fast staining failed to reveal bacteria in every case, whereas modified acid fast and Grocott's methenamine silver consistently demonstrated the characteristic organisms. In both pinnipeds and cetaceans, juvenile animals were affected more often than adults. Hooded seals demonstrated more cases of nocardiosis than other pinnipeds.
Introduction
Nocardia spp. infections are described in a variety of vertebrate and invertebrate species. 5, 44, 51, 53 In mammals, 6 basic forms can be distinguished: pulmonary; systemic; central nervous system; extrapulmonary; cutaneous, subcutaneous, or lymphocutaneous; and actinomycetoma. The pulmonary form involves only the lungs; the systemic form involves 2 or more body sites. Extrapulmonary is any primary focus without involvement of the lung and central nervous system, and actinomycetoma is a distinct subcutaneous nodule. 2, 5, 24 In humans and companion animals, nocardiosis most often results in pneumonia. Mortality is high in humans (50%) and dogs (88.5%), whereas in cattle, the fatality rate is typically much lower (5–10%). The primary presentation in cattle is mastitis, a form of nocardiosis unique to this species. Mortalities in cattle result when Nocardia sp. mastitis advances to systemic infection. 2, 5, 6 The current published reports of marine mammal nocardiosis suggest a fatality rate of 100%. 14, 15, 39, 41, 43, 45, 49, 52 This is likely biased because of the difficulty of confirming clinical cases and their resolution. All currently published marine mammal cases were diagnosed by necropsy examination.
Nocardiosis is reported in both captive and free-ranging marine mammals. Current literature suggests that the systemic form is the most common presentation in cetaceans. 15, 39, 41, 45 There is a single report of pulmonary nocardiosis in a Pacific bottlenose dolphin (Tursiops aduncus). 45 A report in a beluga whale (Delphinapterus leucas) 41 describes mastitis as part of a systemic infection. Only 1 case of nocardiosis in a pinniped has been reported. 14 That case involved a leopard seal (Hydrurga leptonyx) from an aquarium in New Zealand. The seal had a systemic infection involving skin, lung, kidney, and peritoneum. Although hundreds of pinnipeds are rehabilitated in the United States annually, no case of nocardiosis from North America has yet been reported. This low prevalence could be due to differences in exposure to the organism or biological resistance or immune status in pinnipeds compared with cetaceans.
Nocardia spp. are ubiquitous in soil and water and have been identified in marine sediment. They can become airborne, especially on dust particles. 19, 23 The distribution of pathogenic species is not clear, but some species are more common in certain geographic areas or climates. 7, 19, 23, 33, 36 Pinnipeds inhabit the interface of terrestrial and aquatic environments and opportunities for exposure to Nocardia spp. from multiple sources are abundant. Cetaceans are biologically limited to marine environments. Captive housing might enhance their exposure to terrestrial pathogens via aerosols. A comparison of nocardiosis in pinnipeds versus cetaceans and captive versus free-ranging animals might delineate differences due to species or exposure.
We report the results of postmortem examinations and describe the distribution of histologic lesions in 8 hooded seals (Cystophora cristata), a leopard seal, and 3 cetacean species with naturally occurring nocardiosis. We compare the pathogenic species and the character of infections as they relate to the various marine mammal species.
Materials and Methods
Case selection
Facilities in North America involved in diagnostic investigations of marine mammals were queried for suspected or confirmed cases of nocardiosis mortality in marine mammals. These facilities included zoos and aquaria holding marine mammals, facilities that respond to stranded free-ranging marine mammals, and diagnostic services that receive samples from these facilities.
Cases selected for inclusion and further evaluation were limited to those with complete gross and histologic results and culture or polymerase chain reaction (PCR) confirmation of nocardiosis. Cases that reported pyogranulomatous or granulomatous inflammation but lacked culture or PCR confirmation were included after molecular evaluations confirmed the presence of Nocardia sp. For each animal, the following information was obtained: species, gender, age, medical history, gross necropsy findings, histologic findings, and, if possible, species of Nocardia isolated or identified. The age class (juvenile or adult) of the wild animals was estimated on the basis of standard body length measurements or by standard dental aging methods. 27, 35, 47
Nine pinnipeds from 8 institutions met the criteria for inclusion. These consisted of 8 hooded seals and a leopard seal. All were free-ranging animals. Ten cetaceans from 7 institutions met the criteria for inclusion. These consisted of 3 species from wild and captive populations including Atlantic bottlenose dolphin (Tursiops truncatus, n = 4), beluga whale (Delphinapterus leucas, n = 4), and killer whale (Orcinus orca, n = 2). The animals included in the study were from American facilities between 1974 and 2007. The single leopard seal originated in New Zealand; tissues for this animal were sent to the United States for confirmation of nocardiosis.
Histopathology
Tissues available for histologic review varied from animal to animal and included brain, skin, heart, lung, liver, kidney, spleen, lymph nodes, adrenal, stomach, intestines, thyroid, thymus, and pancreas. Tissues were routinely embedded in paraffin, sectioned at 4–7 µm, stained with hematoxylin and eosin (HE), and examined microscopically at the facility of origin. Slides were examined by 2 of the authors (LB and JAS) to assure continuity of descriptions.
To enhance visualization of the organisms, tissue sections were stained with Brown and Brenn (BB), Ziehl-Neelsen (ZN), Fite Faraco (Fites), Coates modified Fite stain (modified Fites), and Grocott's methenamine silver (GMS) according to published protocols. 1
Culture
Diagnostic cultures were performed for 6 pinnipeds and 7 cetaceans. Material was collected at necropsy and inoculated onto blood agar plates and incubated at 37°C under aerobic or microaerophilic conditions for periods ranging from 2 to 10 days.
Polymerase chain reaction
Formalin-fixed, paraffin-embedded tissues (FFPT) of pinniped Nos. 2, 4, and 6–9 and cetacean Nos. 10–14 and 18 were processed for detection of Nocardia spp. deoxyribonucleic acid (DNA). Nine 5-µm sections were deparaffinized in xylene and rehydrated in graded alcohols. DNA was extracted from deparaffinized tissues with a commercially available kit (DNeasy Tissue Kit, Qiagen Inc., Valencia, CA) according to the manufacturer's instructions. A sample of sterile water was extracted in parallel with the tissue samples (extraction negative control). DNA was amplified by forward (CGAGCGTTGTCCGGAATTAC) and reverse (TACTG/TCCCAGAGACCCGC) primers, which amplified a 204-bp fragment of Nocardia spp. 16S rRNA. DNA extracts (2.5 µl) were added to a 25-µl reaction volume containing standard amounts of GeneAmp reagents (Applied Biosystems, Foster City, CA), 25 pmol of each primer, and 1.5 mM MgCl2. Amplification (94°C for 2 min; 35 cycles of 94°C for 1 min, 55°C for 1 min, and 72°C for 1 min; and 72°C for 5 min) was performed in a thermocycler (Model 9700, Applied Biosystems). Negative (sterile water substituted for DNA extract and the extraction negative control) and positive (Nocardia spp. clinical isolates of N. nova, N. farcinica, and N. cyriacigeorgica from terrestrial and marine mammals) controls were included in each set of reactions. PCR products were visualized by agarose gel electrophoresis, ethidium bromide staining, and ultraviolet transillumination and then purified with the use of a commercially available kit according to the manufacturer's instructions (ExoSAP-IT, USB Corporation, Cleveland, OH). Nucleotide sequences were determined on an automated sequencer (3730XL, Applied Biosystems) at the University of Chicago Cancer Research Center DNA Sequencing and Genotyping Facility. Nucleotide sequences were compared with sequences obtained from GenBank for identification. Only sequences with greater than 98% similarity to a confirmed isolate were speciated; all other sequences were recorded as Nocardia sp.
Results
Animals
Nine pinniped cases from 1974 to 2007 met the study inclusion criteria. The cases were limited to 8 hooded seals and a single leopard seal. Age, species, and sex distribution can be found in Table 1. The pinnipeds were all free-ranging animals in rehabilitation for periods ranging from 5 days to greater than 1 year before death. The affected hooded seals were stranded along the east coast of the United States—Florida (case Nos. 1 and 8), Connecticut (case No. 2), Virginia (case No. 4), North Carolina (case No. 6)—California (case No. 3), Puerto Rico (case No. 7), and the port of Montreal (case No. 5). The leopard seal (case No. 9) was stranded in Hawke's Bay on the north Island of New Zealand. The animal that was in hand only 5 days demonstrated both chronic and acute lesions, suggesting infection before entering rehabilitation. Three animals demonstrated signs only after being in rehabilitation for at least 3 weeks (case Nos. 1, 3, and 7). Six animals demonstrated signs related to nocardiosis on entry into rehabilitation (case Nos. 2, 4, 5, 6, 8, and 9) and were treated for a period of more than 1 year before death. One hooded seal was euthanatized (case No. 6); the other animals died spontaneously.
Distribution and severity of histologic Nocardia spp.-associated lesions in pinnipeds.*

- = no gross or histological lesions associated with Nocardia; + = mild; ++ = moderate; +++ = severe; nr = information not recorded; HS = hooded seal; LS = leopard seal; F = female; M = male.
Ten cetaceans from 1977 to 2007 met the criteria for inclusion. The species distribution included 3 Atlantic bottlenose dolphins, 4 beluga whales, and 2 killer whales. Age, species, and sex distribution of these animals can be found in Table 2. Eight cetaceans were from North American aquaria, and 2 were free-ranging animals stranded in Florida (case Nos. 10 and 11). One of the free-ranging dolphins was collected dead on the beach and thus must have become infected while in the wild. The second was in rehabilitation for 14 days (case No. 11). The duration of illness of the captive cetaceans ranged from 14 days to almost 2 years. One beluga whale was euthanatized because of the severity and progression of the disease (case No. 18); the other animals died spontaneously.
Distribution and severity of histologic Nocardia spp.-associated lesions in cetaceans.*
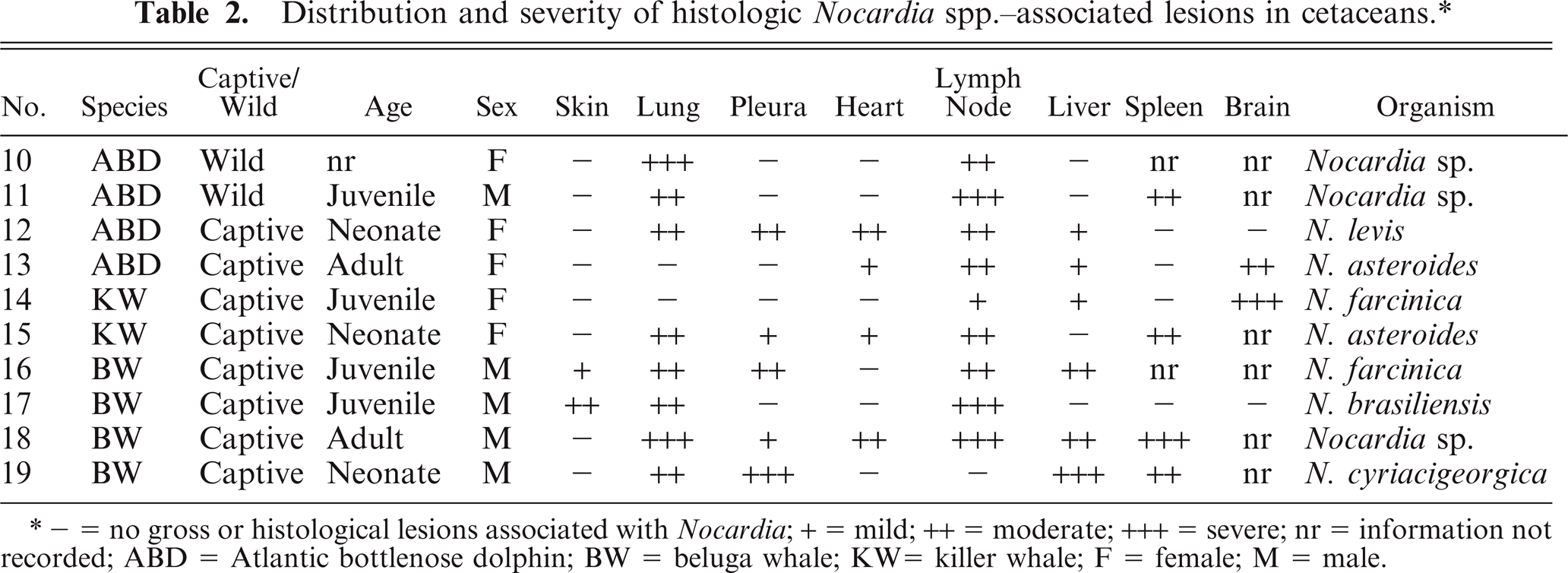
- = no gross or histological lesions associated with Nocardia; + = mild; ++ = moderate; +++ = severe; nr = information not recorded; ABD = Atlantic bottlenose dolphin; BW = beluga whale; KW= killer whale; F = female; M = male.
Pathologic findings
Thoracic disease as a component of systemic infection was the primary presentation in hooded seals. Pyothorax was present in 3 hooded seals (Nos. 1, 4, and 6). Single or multiple pulmonary abscesses or pyogranulomas ranging from 2 mm to 10 cm were seen in 5 hooded seals (Nos. 2–6). All seals with pulmonary lesions had thoracic lymph node abscesses (Figs. 1, 2). Four of these 5 hooded seals had generalized lymph node abscesses (Nos. 2, 3, 5, and 6). Two hooded seals had pleuritis (Nos. 1 and 6), 2 had brain abscesses (Nos. 7 and 8), and 2 had abscesses or pyogranulomas in skin or subcutaneous tissues (Nos. 2 and 3; Fig. 3). The cervical skin lesion in hooded seal No. 2 extended diffusely into the musculature. In hooded seal No. 3, a lesion in the right nasal cavity extended into the surrounding muscle and bone. Pyogranulomas were found grossly in liver (Nos. 2, 4, and 5) and spleen (Nos. 5 and 6) of 4 animals. Three hooded seals were in good nutritional condition at necropsy (Nos. 3, 4 and 8) and 2 were thin or emaciated (Nos. 2 and 5). Distribution and severity of relevant microscopically recognized lesions in pinnipeds are summarized in Table 1. Six hooded seal cases had concurrent conditions, including injuries from shark bites (Nos. 6 and 7), bacterial valvular endocarditis (No. 2), a traumatic lesion in oral cavity (No. 4), and parasitic ulcers in the gastrointestinal tract (Nos. 2 and 3). One hooded seal had an eye removed because of glaucoma 2 weeks before death (No. 8). None of these animals had lesions suggestive of concurrent immune suppression or morbillivirus infection.

Thoracic viscera; hooded seal No. 1. The tracheobronchial lymph nodes (arrows) of this hooded seal with nocardiosis are massively enlarged and abscessed.
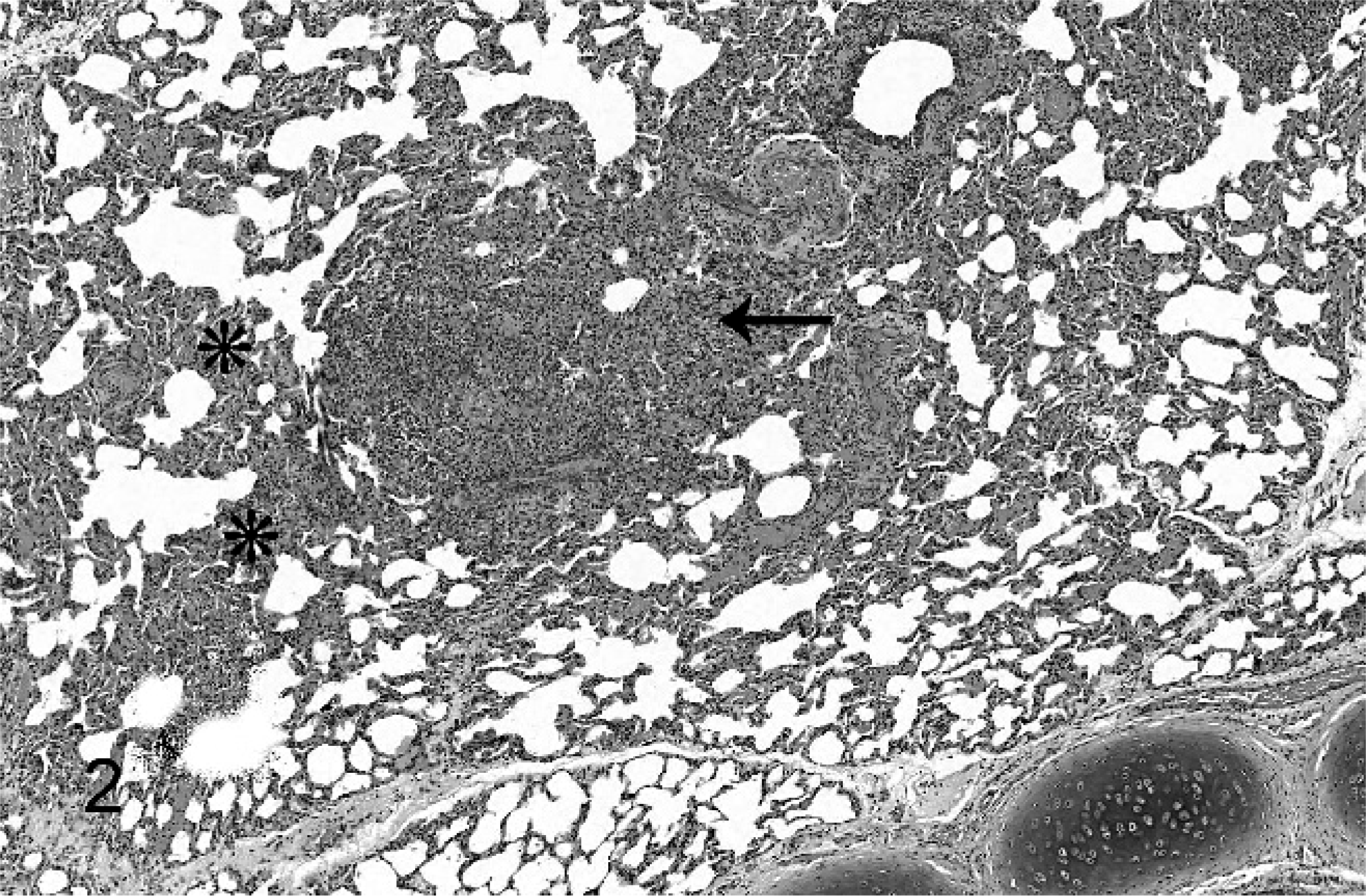
Lung; alveolar septa and a bronchiole from a hooded seal with nocardiosis. A granuloma (arrow). The alveolar septa are locally expanded with macrophages and lymphocytes (asterisks). No filamentous bacteria are seen with this stain. HE.
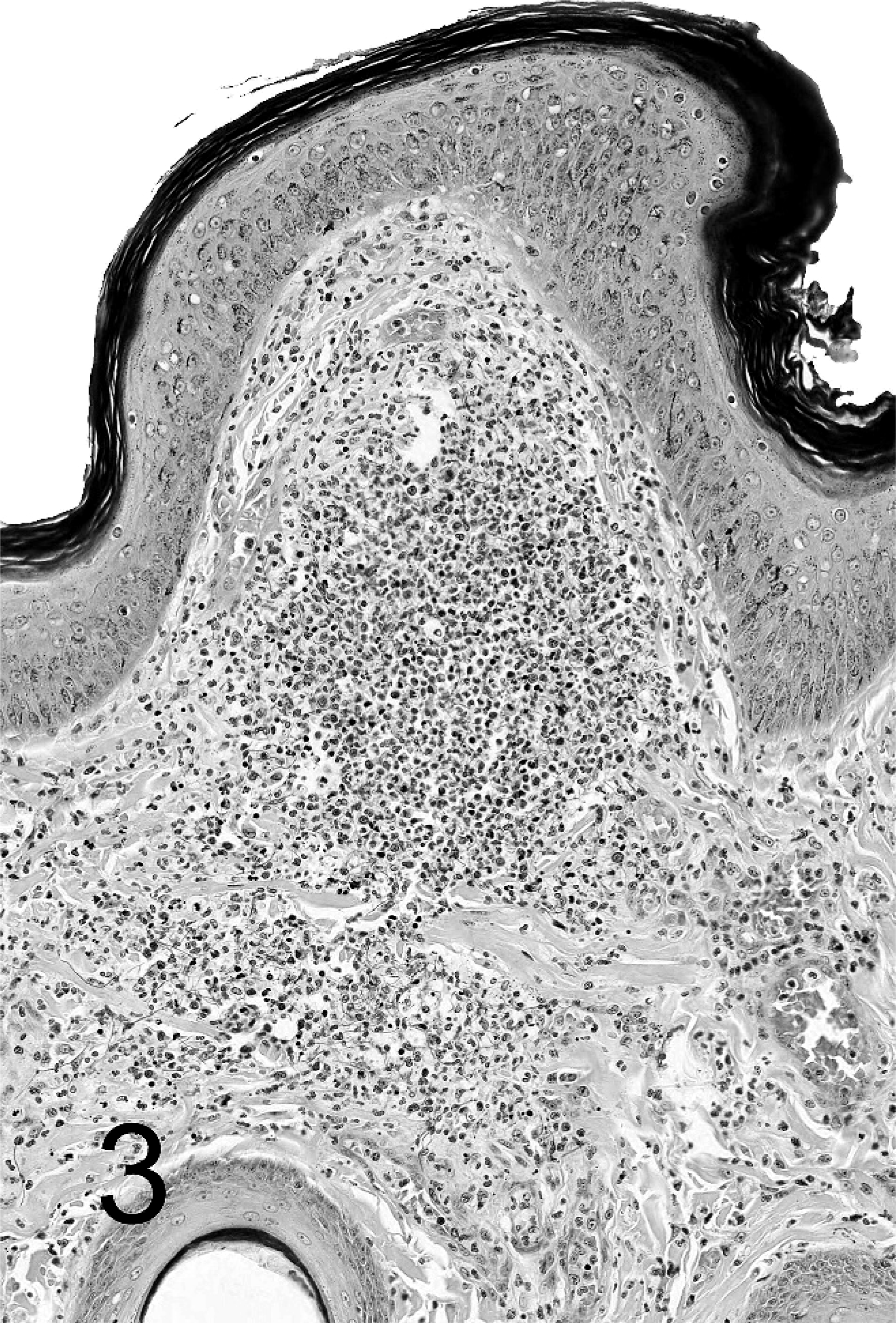
Skin; hooded seal No 2. Granulomatous focus of necrotic and viable neutrophils and macrophages in dermis with indistinct border. BB.
Gross and histologic evaluations in the leopard seal revealed multiple pyogranulomas in lung, thoracic lymph nodes, adrenals, and brain.
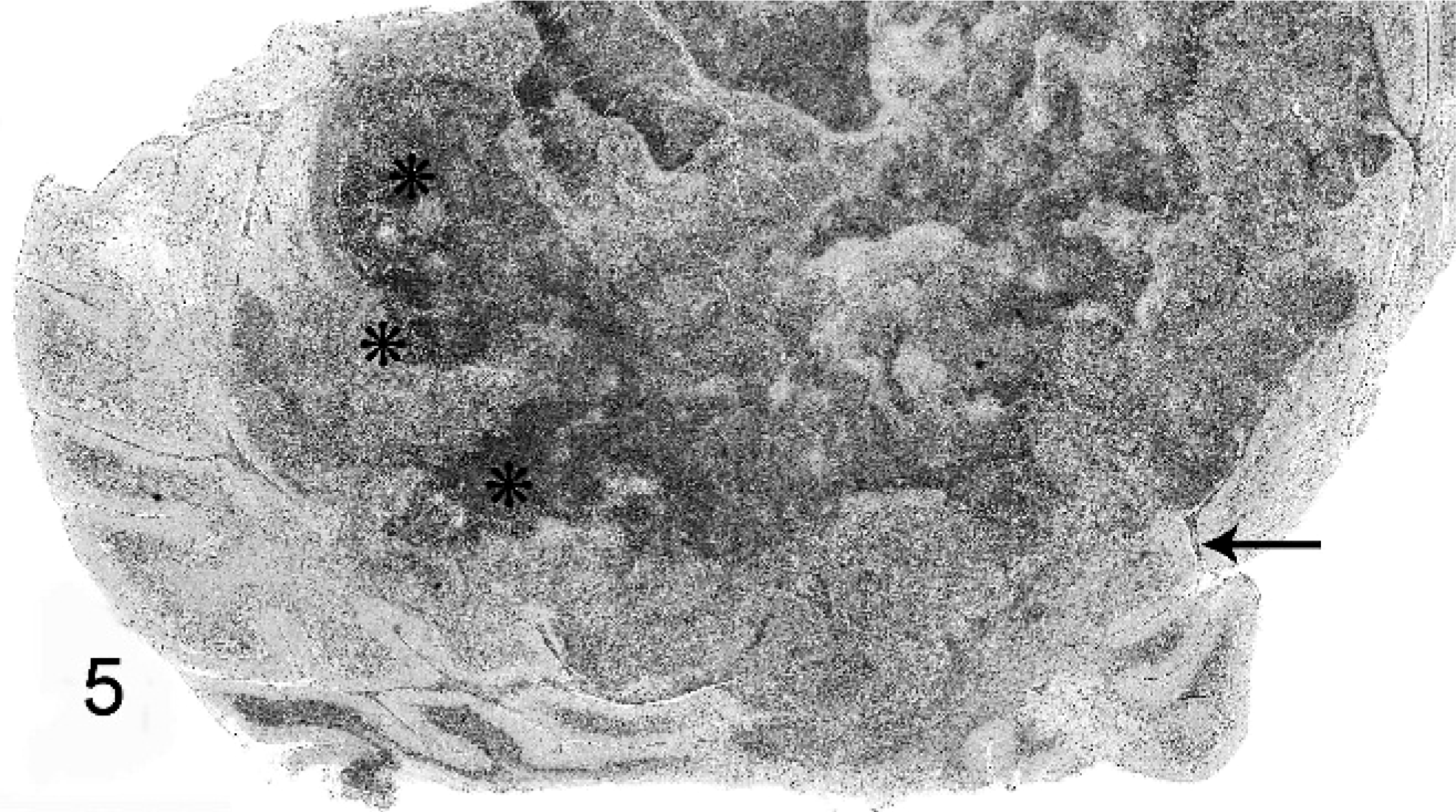
Distribution and severity of relevant lesions in the cetaceans are summarized in Table 2. Pulmonary pyogranulomas with thoracic lymph node abscesses were the most common presentation in cetaceans (Fig. 4). These were seen in 6 cetaceans (Nos. 12, 13, 15, 16, 18, and 19). Three of these animals had pleuritis and moderate amounts of clear yellow or serosanguineous thoracic fluid (Nos. 15, 18, and 19). The characteristics of the fluids are not recorded. Five of these 6 cases had generalized lymph node abscessation (Nos. 11, 15, 16, 18, and 19). Two cetaceans had brain abscesses (Nos. 13 and 14; Fig. 6). In cetacean No. 17, a pyogranulomatous dermatitis and cellulitis extended to the underlying vertebra. The vertebral body demonstrated focally extensive chronic osteonecrosis and remodeling (Fig. 5). Abscesses or pyogranulomas were also recognized in aortic endothelium (cetacean No. 18), spleen (Nos. 11, 15, 18, and 19), adrenals (Nos. 15 and 19), intestine (No. 11), humeroscapular joint (No. 14), kidney (No. 18), and liver (No. 19). Four cetaceans were in good nutritional condition at necropsy (Nos. 14, 15, 17, and 19), and 3 were thin or emaciated (Nos. 10, 12, and 19). Five of 10 cetaceans had concurrent conditions that included granulomatous dermatitis from lacaziosis (cetacean No. 13), ulcerative dermatitis containing ciliated protozoa (No. 10), skin lesions due to trauma (No. 11), and parasitic ulcers in the gastrointestinal tract (Nos. 11, 14, and 17). None of the animals had histologic findings suggestive of concurrent morbillivirus infection.

Lung; beluga whale No. 15. This lung of a beluga whale shows multifocal pyogranulomas scattered throughout the parenchyma.
Cerebellum; Atlantic bottlenose dolphin No. 13. The right area of the cerebellum demonstrates a severe suppurative encephalitis, the border is indicated by asterisks. Nonsuppurative meningitis (arrow). HE.

Thoracic vertebra; beluga whale No. 17. Focally extensive osteonecrosis with remodeling.
Histologic staining was evaluated for the ability to identify slender and filamentous bacteria typical of Nocardia sp. HE staining demonstrated the inflammatory reaction but failed to adequately reveal bacteria. Gram's stains consistently demonstrated organisms (Fig. 7). However, 9 samples revealed gram-positive bacteria and 2 demonstrated gram-variable Nocardia-like organisms. ZN staining conducted on tissues from 7 cases of Nocardia demonstrated acid-fast bacteria in 4 and variably acid-fast bacteria in 3 others. In 6 cases, a modified Fites or Fites stain was performed. Both stains consistently demonstrated numerous acid-fast bacteria. GMS stain consistently demonstrated argyrophilic bacilli in all of the 9 tissues on which this stain was performed (Fig. 8).
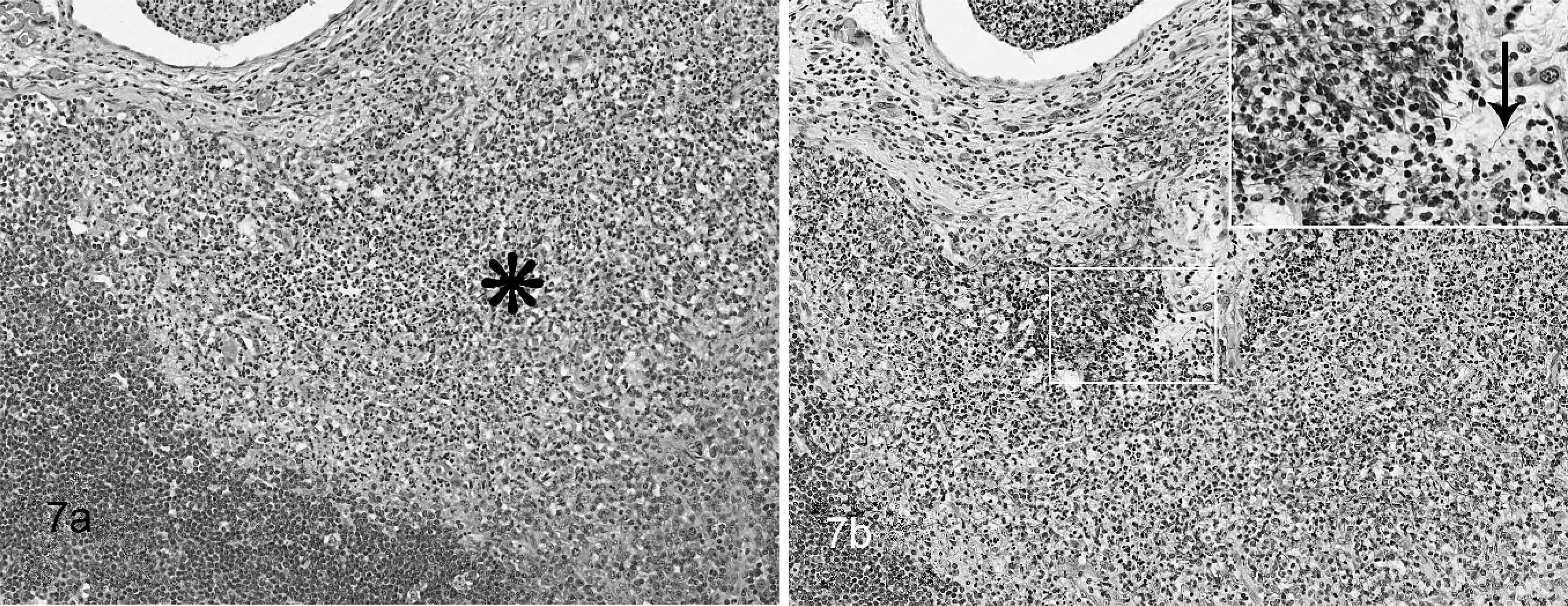
Lymph node; hooded seal No. 2. Granulomatous lymphadenitis Fig. 7a. The sinus is infiltrated by a neutrophils and macrophages (asterisk). No bacteria can be seen with this stain. HE. Fig. 7b. Multiple filamentous Nocardia sp. bacteria (arrow). The ZN staining technique demonstrates organisms, but the staining reaction is variable. Inset: Higher magnification. BB.

Adrenal gland; beluga whale No. 19. Fig. 8a. Large suppurative circumscribed nonencapsulated focus in zona fasciculata of adrenal gland. HE stain shows no bacteria. Fig. 8b. Section adjacent to 8a. The stain shows filamentous argyrophilic Nocardia sp. dispersed mainly at the border of the lesion. Inset: Higher magnification. GMS.
Bacterial culture and identification
Nocardiosis was confirmed by bacterial culture in 5 pinniped and 6 cetacean cases. For confirmation and species identification, PCR with sequencing of amplicons was performed on FFPE tissue in 5 pinnipeds and 7 cetaceans. In 13 of the 17 cases, bacterial culture, PCR, or both were able to speciate the Nocardia organisms.
A variety of Nocardia spp. caused fatal infections in marine mammals (Tables 1, 2). In pinnipeds, the pathogenic species include N. asteroides, N. otitisdiscaviarum, N. farcinica, and N. brasiliensis. The pathogenic species in cetaceans include N. asteroides, N. brasiliensis, N. farcinica, N. levis, and N. cyriacigeorgica.
Discussion
The lesions caused by Nocardia spp. in marine mammals were mild to severe multifocal pyogranulomas. This is consistent with the condition in man and domestic animals.∗ The most common presentation of nocardiosis in both pinnipeds (7 of the 9 cases) and cetaceans (9 of the 10 cases) was a systemic distribution. The most frequently affected organs were lungs and thoracic lymph nodes. Systemic distribution included brain abscesses in 2 pinnipeds and 2 cetaceans. Nocardiosis should be considered a differential diagnosis for pulmonary or neurologic disease in all marine mammals.
Our findings support and further the current knowledge of nocardiosis in marine mammals. The results of the currently published data are summarized in Table 3. A literature review demonstrates that the systemic presentation occurs most commonly, with the lungs most frequently affected. A single published case identifies involvement of the brain. 15 Brain tissues are often not examined, which could account for the infrequent reporting of brain involvement in marine mammal nocardiosis. The current literature describes an interesting Nocardia sp. mastitis case in a female beluga whale. Nocardia spp. mastitis is a condition of dairy cattle 6, 12, 20, 30, 48 but is otherwise rarely noted. In this study, no cases of mastitis were identified. The thoracic effusions seen in the cetaceans in this study are not previously reported. The character of the effusion was distinctly different from the pyothorax noted in 3 pinnipeds. These might reflect cardiovascular involvement or a unique pleural or lymphatic reaction to inflammation in these species. Further characterization of these fluids is needed in future cases.
Literature cases with pathology results of marine mammal nocardiosis and distribution of Nocardia-associated lesions.*

+ = Nocardia spp.-associated lesions mentioned; nr = information not recorded; PBD = Pacific bottlenose dolphin; SD = striped dolphin; HP = harbor porpoise; BW = beluga whale; PW =pilot whale; LS = leopard seal; F = female; M = male.
Nocardia spp. are relatively slow growing bacteria. Culture takes typically 3–5 days and can take up to 6 weeks. The relatively slow growth permits overgrowth by concurrently sampled bacterial species. 2, 13, 36, 54 In this study, 7 cases of presumptive nocardiosis were based on histologic findings alone. PCR proved to be a valuable way to identify Nocardia in the animals of this study and to confirm the diagnosis of nocardiosis in 11 of the animals. Improved culture techniques, or the use of molecular diagnostics can increase the definitive diagnosis of nocardiosis. PCR techniques identified a variety of Nocardia spp. as pathogenic in marine mammals. These investigations demonstrated an expanded host and geographic range of pathogenic Nocardia sp. No species specificity or increased prevalence in captive versus free-ranging populations was noted. N. otitisdiscaviarum was found only in pinnipeds and N. levis and N. cyriacigeorgica were found only in cetaceans. The small numbers of cases identified suggests that further case evaluations might expand the pathogenic spectrum in the future.
A review of the staining characteristics in these cases proved surprising. Hematoxylin and eosin staining failed to reveal the bacteria. Nocardia spp. were variably acid-fast with the ZN staining and gram-variable with BB staining. This variation could be due to changes in the tissue chemistry because of infection-associated necrosis or changes in the bacterial chemistry. 2, 3, 4, 21, 22 Modified acid-fast stains like the modified Fites, were consistently successful in demonstrating acid-fast–positive organisms. GMS was consistently successful in highlighting the Nocardia sp. organisms.
Nocardiosis has 3 main routes of infection: inhalation, inoculation, and ingestion. 2, 6, 13, 42 The high proportion of pulmonary involvement suggests that inhalation and aspiration are the major routes of infection for both cetaceans and pinnipeds. However, the route of infection is difficult to determine in most of our cases. Nocardia has been documented in fish and shell fish species. Half of the cases in both the pinnipeds and cetaceans had concurrent conditions such as skin lesions and gastrointestinal ulcers. These lesions could be another portal of entry. By this route of inoculation, Nocardia spp. can get into the bloodstream and cause infection in any organ, without establishing infection in lung or skin. 8, 11, 38, 42, 46, 50 In 2 hooded seals and 1 beluga whale with skin lesions, the medical history reveals that the skin lesions were noticed before the respiratory signs (animal Nos. 2, 3, and 17). The skin might have been the portal of entry in these animals. The systemic presentation that was most common in this study is seen in most other mammals and is best studied in humans. In humans, the primary focus of infection is mostly the lung or skin. The initial primary pulmonary or cutaneous infection has the tendency to clear spontaneously after dissemination to other organs has occurred. 37 This could be the course of infection in the 2 hooded seals (Nos. 7 and 8) and 2 cetaceans (Nos. 13 and 14) that did not show infection of skin and lung. The clinical history fails to provide decisive answers.
Nocardia bacteria are ubiquitous in the environment and can be found worldwide in fresh- and salt water, soil, dust, decaying vegetation, and decaying fecal deposits from animals. 7, 19, 20, 33 For most Nocardia sp. identified, there is no known environmental predisposition. The identified species do not help to identify the source of infection. Future Nocardia sp.–specific research of soil and water might provide clarity in this area. Molecular characterizations identified an expanded pathogen range for Nocardia sp. in marine mammals. However, comparisons of the various species affected and the associated syndromes do not indicate ancillary features of pathogenicity. Enhanced bacterial identification in future cases might provide the information needed to identify these patterns.
Current literature includes 16 cetacean cases of nocardiosis with variable demographic data available. 15, 18, 25, 29, 41, 45, 49, 52 In previously reported and present cases, captive animals are in the majority (of 26 cases, 18 were captive and 8 were wild). This discrepancy is likely due to better postmortem carcass condition and more thorough examination of captive cetaceans. The medical history of the 2 free-ranging cetaceans and a single hooded seal suggests that these animals were infected with Nocardia spp. while in the wild. The age and sex distribution previously reported and seen in this study suggest that age and sex are not predisposing factors in cetaceans. Ten different odontocete species have reported nocardiosis with no indication of a species predilection. Because Nocardia spp. infections can demonstrate variable chronicity, 2, 10, 40 it is difficult to determine the duration and origin of infection. The age and sex distributions in pinnipeds suggest that juvenile male pinnipeds are more susceptible. This bias could be due to the relatively small number of cases.
Hooded seals are dramatically overrepresented in this review. All affected hooded seals were free-ranging animals. They are held in very low numbers in North America only for research. 35 Given the high levels of rehabilitation for sea lions and harbor seals and the presence of these animals in captive collections, the lack of nocardiosis in these species compared with the hooded seals is very unusual. An inherent species susceptibility or immune deficit might be responsible for the high prevalence in this species. Nocardia spp. are considered to be opportunistic pathogens that typically cause systemic disease in individuals with causes of immunosuppression. 2, 9, 31, 40 No corticosteroid treatment before Nocardia sp. infection was reported in any of the cases. Well-known underlying causes of immunosuppression in marine mammals, like morbillivirus infections, 32 and toxicants such as heavy metals and organochlorines 16, 17, 26 were not identified, but neither were they specifically investigated. Environmental stressors are another possible cause of impaired immune function. All pinnipeds identified in this study were found outside their normal range. Aberrant environmental conditions and changes in prey availability could play a role in promoting nocardiosis in these cases. In future evaluations of marine mammal nocardiosis, evaluations for predisposing or concurrent viral and toxic conditions are indicated.
Nocardiosis is a significant cause of mortality in marine mammals. Fatal infections are most typically systemic. In addition to the previously reported species, hooded seals expand the list of affected marine mammals. Hooded seals appear to be particularly vulnerable to this infection compared with other pinnipeds. Diagnosis can be facilitated with GMS staining of granulomas. PCR is an effective way to confirm the diagnosis and determine the species of Nocardia. Molecular characterization of the pathogenic isolates expanded the known pathogenic species of Nocardia. Molecular identification and bacterial culture proved that N. asteroides, N. farcinica, N. brasiliensis, and N. otitisdiscaviarum are pathogenic for pinnipeds. In cetaceans, N. asteroides, N. farcinica, N. brasiliensis, N. cyriacigeorgica, and N. levis are pathogenic. Investigations into immune condition or associated viral infections might enhance our understanding of the infections in this species.
Footnotes
Acknowledgements
We are grateful to clinicians and pathologists that provided us with case materials, including Gregory Bossart, Paul Calle, Terry Campbell, Ronald Crowley, André Dallaire, Les Dalton, Fabio Del Pierro, Christopher Dold, Sam Dover, Lawrence Dunn, Kim Durham, Ruth Ewing, Michael Kinsel, Thomas Lipscomb, Dee McAloose, Richard Montali, Wendy Noke, Todd Robeck, René Roy, Todd Schmitt, Robert Schoelkopf, Megan Stolen, and Mike Walsh. We thank Robert Couey for graphic assistance as well as the staff of NOAA and US Marine Mammal Stranding Network for the stranding statistics, including Carley Lowe, Sarah Wilkin, Aleria Jensen, Joe Cordaro, and Elizabeth Tuohy-Sheen. We thank Stacy Schultz for technical assistance. Work in the laboratory of Dr. Terio was supported by an Office of Naval Research grant (N00014-06-1-0249).