
Abstract
Endothelial progenitor cells (EPCs) participate in vascular healing during both acute injury and chronic disease. The quantity and quality of circulating EPCs correlate inversely with the severity of vascular disease, such that reduced number and/or function of EPCs are significant independent risk factors for impaired healing capacity, dysfunctional endothelium, and progression of atherosclerosis and vascular disease. EPC therapy assists healing of cardiac and limb ischemia and has great potential for improving the quality of life and longevity of patients with severe cardiovascular and peripheral vascular disease who are not candidates for conventional revascularization procedures. In addition, EPCs can be used to promote vascular graft patency. This review focuses on the characterization of EPCs, positive and negative regulators of EPCs, the role of EPCs in vascular disease, and the potential for EPC therapy to ameliorate the sequelae of severe peripheral vascular disease.
Keywords
The ability of endothelium to repair itself depends on the recruitment, proliferation, and migration of surrounding mature endothelial cells and the mobilization and incorporation of circulating endothelial progenitor cells (EPCs) to the injured region. Recent studies have outlined the importance of EPCs with respect to atherosclerotic vascular disease, acute vessel injury, and tissue ischemia. EPCs are thought to promote healing of the endothelial monolayer and vessel wall primarily through processes of reendothelialization and neovascularization. These repair mechanisms have been shown to reduce the risk of atherosclerotic disease progression and minimize the negative sequelae of vessel wall injury and tissue ischemia.
Risk factors such as elevated cholesterol, hypertension, diabetes, smoking, physical inactivity, and advanced age are associated with chronic damage to the endothelial monolayer. 1 In addition, vessel trauma or ischemia can acutely compromise the integrity of the endothelium and result in endothelial cell apoptosis. 2 Vascular damage is repaired by two synergistic mechanisms, mediated by two different cell populations. Repair by local proliferation and migration of resident mature endothelial cells can repopulate the endothelial layer, but these cells have a limited capacity for regeneration; they become senescent and insufficient for proper healing as successive replications result in telomere shortening. Alternatively, circulating bone marrow–derived EPCs have been shown to migrate toward sites of injury and ischemia, proliferate, differentiate, and incorporate into injured vessel walls, as well as generate new vessels. This repair mechanism provides a source of healthy endothelium with longer telomeres and a renewed ability to respond to further vascular damage. However, EPCs are also susceptible to senescence and dysfunction; normal aging in the presence of risk factors for vascular disease can lead to the progressive depletion of marrow cells that give rise to progenitors necessary for arterial repair. 1,3 EPC obsolescence implies insufficient circulating number and/or functional capacity of EPCs and, consequently, a potential inability to maintain vascular integrity. In the setting of significant risk factors and a dysfunctional endothelium, which has a limited capacity to heal acute vessel injury, compensate for chronic vascular damage, and limit abnormalities in vasoreactivity, atherosclerotic disease will progress. 3
Pathophysiology of Atherosclerosis
Endothelial damage results in the release of chemokines and cytokines; these factors promote mobilization of EPCs from the bone marrow, migration to the peripheral circulation, and homing to sites of injury or ischemia. This activity is coordinated by complex interactions involving: vascular endothelial growth factor (VEGF), which responds to tissue ischemia, assists in the recruitment of EPCs, promotes new vessel formation, and is correlated with rapid EPC recruitment after traumatic vascular injury 4 ; granulocyte colony-stimulating factor (G-CSF), a growth factor for neutrophils that also mobilizes EPCs from the bone marrow; matrix metalloproteinase (MMP)-9, a type 4 collagenase and gelatinase that cleaves the extracellular matrix to allow EPC migration; and stromal cell–derived factor 1 (SDF-1), a chemokine that attracts EPCs to the site of injury.
Under circumstances in which a competent bone marrow is able to produce a sufficient quantity of functional EPCs, these cells arrive at the site of injury and proliferate, differentiate, and incorporate into the vessel wall. Vascular healing occurs via reendothelialization and neovascularization with resolution of the local inflammatory response; a negative feedback loop results in termination of cytokine release in the absence of inflammation. 1
If the bone marrow is not competent and produces an insufficient quantity of functional EPCs in response to the local inflammation at the site of vascular damage, then healing does not occur. This may result from ineffective recruitment or impaired function of senescent EPCs. In this setting, cytokines continue to be released by local inflammatory cells, causing further inflammation and tissue damage via a positive feedback loop. The failure of aging organisms to renew endothelial cells exposed to noxious stimuli leads to endothelial dysfunction, inflammation, and atherosclerosis. 1,3
EPC Number Is Inversely Correlated with Vascular Disease Severity
Atherosclerosis is the result of chronic chemical or mechanical stresses that cause vascular damage in the setting of deficient repair mechanisms. It has been shown that the number of circulating EPCs is a significant independent predictor for the severity of atherosclerosis. Hill and colleagues measured the number of circulating EPCs in 45 men with a mean age of 50 years having various degrees of cardiovascular risk but no history of cardiovascular disease. 5 Endothelium-dependent and -independent vascular function was assessed by ultrasonography of the brachial artery. Interestingly, the researchers observed a strong inverse correlation between the number of circulating EPCs and the combined Framingham risk score, which predicts a subject's risk for cardiovascular disease. They also observed a significant positive correlation between the number of circulating EPCs and brachial artery endothelial function.
Werner and colleagues determined the number of EPCs in 519 patients with angiographically proven coronary artery disease (CAD) and evaluated the association between baseline levels of EPCs and major cardiovascular events after 12 months. 6 After adjustment for relevant variables, increased levels of EPCs were associated with a reduced risk of death from cardiovascular causes (hazard ratio 0.31), a first major cardiovascular event (hazard ratio 0.74), revascularization (hazard ratio 0.77), and hospitalization (hazard ratio 0.76). Reduced levels of EPCs were a significant, independent predictor of poor prognosis even after adjustment for traditional cardiovascular risk factors and disease activity. 7
Characterization of EPCs: Lineage, Markers, and Abundance
EPCs are rare nonleukocyte bone marrow–derived mononuclear cells (BMCs) that are enriched in the mononuclear cell fraction of peripheral blood. These cells were first isolated and induced to differentiate into endothelium-like cells in vitro by Asahara and colleagues in 1997. 8,9 Many aspects of EPC characterization are currently debated, including their lineage, markers, and the best methods for isolation and culture.
During embryonic development, the mesodermal layer develops into bone marrow and resident stem cells, including hematopoietic and mesenchymal stem cells. It is likely that EPCs are derived from hemangioblasts, the multipotent stem cells that are a common precursor to hematopoietic stem cells (HSCs) and EPCs; however, EPCs may also be derived directly from HSCs. 10–12 Alternatively, there is evidence to show that myeloid cells-bone marrow–derived hematopoietic committed progenitor cells-may transdifferentiate into EPCs. 13 This lineage is shown in Figure 1.
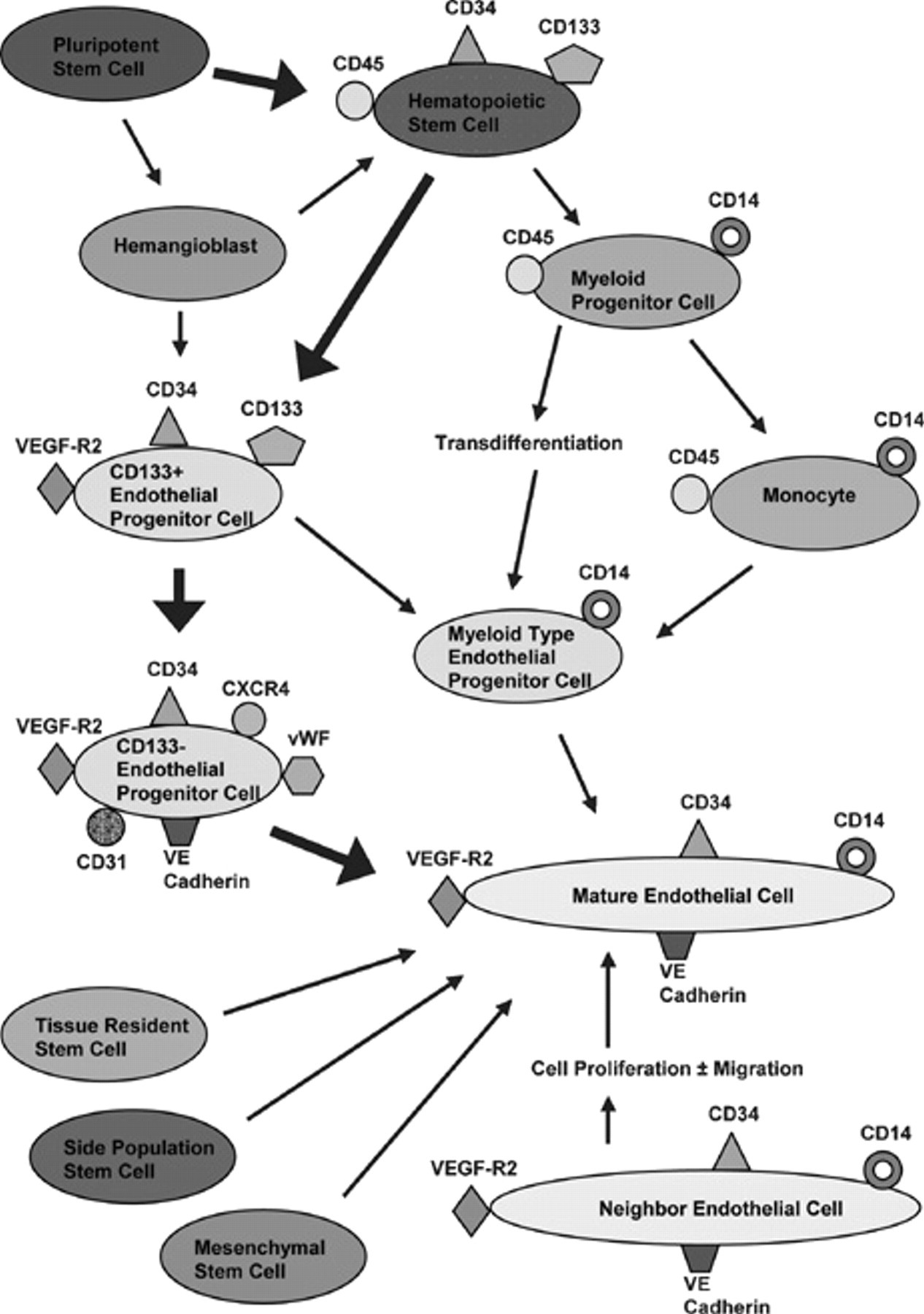
Contribution of endothelial progenitor cells (EPCs) and mature endothelial cells to the population of endothelial cells available for vascular repair. Proposed lineage and cell surface markers of EPCs are in the upper portion of the figure; mature endothelial cell contribution is in the lower right portion of the figure. Bold arrows show the pathway most commonly cited. Adapted from Urbich C and Dimmeler S. 10 VE cadherin = vascular endothelial cadherin; VEGF = vascular endothelial growth factor; VEGF-R2 = vascular endothelial growth factor receptor 2; vWF = von Willebrand factor.
In addition, several other non-HSC types have been induced to differentiate into mature endothelial cells. For example, multipotent adult progenitor cells isolated with mesenchymal stem cells and lacking HSC markers can be cultured with VEGF and induced to differentiate into an endothelial phenotype that expresses endothelial cell markers, functions as a mature endothelial cell in vitro, and contributes to both wound healing and tumor angiogenesis in vivo. 14
Tissue-resident stem cells may also give rise to endothelial cells under certain conditions. For example, adult cardiac stem cells have been induced to differentiate into vascular tissues, including vascular smooth muscle cells (SMCs) and endothelial cells. In one important study, Beltrami and colleagues demonstrated that an injection of clonally expanded tissue-resident cardiac stem cells into an ischemic heart resulted in regeneration of well-differentiated myocardium, including new vessels and myocytes with the characteristics of young cells. 15
Side population (SP) cells, a heterogeneous cell population identified by a capacity to efflux Hoechst-33342 and rhodamine 123 dyes, are composed of HSCs and non–hematopoietic organ–specific multipotent stem cells. 16 SP stem cells are able to acquire the endothelial cell phenotype with VEGF stimulation or acquire the SMC phenotype with transforming growth factor β1 or platelet-derived growth factor BB stimulation; in addition, SP cells can form branching vascular-like structures with evidence of both cell types in vitro. 17
EPCs have also been defined by their functional capacity to form endothelial cell colonies and incorporate acetylated low-density lipoprotein. In addition, they may exhibit enhanced endothelial nitric oxide synthase (eNOS) expression after shear stress exposure. 18,19
EPCs can be further characterized using cell surface markers. Most researchers agree that EPCs express the following markers: CD34, which is present on certain hematopoietic cells, EPCs, and endothelial cells; vascular endothelial growth factor receptor 2 (VEGF-R2), which is essential for angiogenesis and vasculogenesis in conjunction with its ligand VEGF; and CD133, which is present on a variety of stem cells. It should be noted that EPCs in the bone marrow and early circulating EPCs express CD133, but after differentiation occurs, circulating EPCs lose CD133 and gain several other surface markers, including platelet endothelial cell adhesion molecule (CD31), vascular endothelial cadherin (VE cadherin) and von Willebrand factor (vWF). 2,20 Another important cell surface marker is CXCR4, which is the receptor for SDF-1; SDF-1 is upregulated by hypoxia-inducible factor 1 (HIF-1) in the setting of tissue hypoxia. The CXCR4-SDF-1 pair is known to be important in HSC trafficking, including quiescence, homing to the bone marrow, and mobilization to sites of vascular injury and ischemia (see Figure 1). 21 Of note, interfering with this receptor–ligand interaction can mobilize HSCs into the circulation by disrupting the CXCR4-SDF-1 bond that tethers these cells to the bone marrow.
CD133 and CD34 are often used as markers for selection and purification of EPCs since CD133 is not found on mature endothelial cells and CD34 is not found on the undifferentiated stem cells from which EPCs derive. 22 However, there is significant overlap in terms of cell surface markers. For example, mature endothelial cells shed from the vessel wall may express VEGF-R2, VE cadherin, and CD34. In addition, certain HSCs may express CD34, CD31, VEGF-R2, and vWF. 11 CD133 is found on bone marrow EPCs and early circulating EPCs, but the specificity of this marker is compromised because hematopoietic progenitor cells may also express CD133. 2,23
The lack of a specific marker that is present on all EPCs implies a technical challenge with respect to isolating these cells from peripheral blood and has hampered the development of an established protocol for isolating these cells. Lack of a specific marker has been cited as a potential source for conflicting evidence in studies evaluating the number of circulating EPCs. 1 In addition, the possibility that there may be subpopulations of EPCs with different functional characteristics and surface markers adds to the complexity of the issue. 24 One consequence of this problem is that many of the clinical studies evaluating the therapeutic effects of EPCs have been performed using bone marrow samples that have not been purified to exclude other cell types, leading to the possibility that functional effects ascribed to EPCs may not be due to these cells alone. 11
Another confounding factor is that other cell types bearing different cell surface markers and derived from bone marrow, peripheral blood, or specific tissues may be precursors to endothelial cells, making the “true” EPCs more elusive. As described above, this is further complicated by the fact that other stem cell types may be induced to transdifferentiate into EPCs, including mesenchymal stem cells and the SP phenotype of human stem cells (see Figure 1). 9,11
During steady-state conditions in healthy patients, circulating EPCs comprise approximately 0.01% of peripheral blood cells. 11 It has been repeatedly demonstrated that patients with chronic vascular disease have even fewer circulating EPCs. The rarity of this cell population is another factor that makes isolation of these cells for therapy or for research very challenging.
Regulators of EPC Number and Function
There are many positive and negative regulators of EPC. These factors have a quantifiable effect on EPC quantity and quality, as described in the following studies (Figure 2).
Positive and negative regulators of endothelial progenitor cell (EPC) number and/or function.
Negative Regulators
Aging
Heiss and colleagues evaluated circulating EPC number and function in 20 aged and young subjects with no major cardiovascular risk factors. 25 They reported that although older people had impaired endothelial function, there was no significant difference in the number of circulating EPCs between the two age groups. However, EPCs from older people had significantly decreased survival, migration, and proliferation rates, suggesting that the impaired maintenance of the endothelium in older subjects correlates with functional EPC deficits rather than total EPC number. 25,26
Normal aging may diminish the potential for mature endothelial cells to participate in effective vascular repair, resulting in progression of atherosclerosis. Impaired endothelial cell function, bone marrow senescence, and limited availability or compromised function of circulating EPCs are all factors that contribute to decreased reserve for repair of vascular injury with aging.
Hypercholesterolemia
Hypercholesterolemia is a significant risk factor for atherosclerosis. Rauscher and colleagues performed an elegant study demonstrating that long-term treatment with bone marrow–derived progenitor cells from young nonatherosclerotic apolipoprotein E (ApoE)-deficient mice was successful in preventing atherosclerosis progression in ApoE-deficient recipients despite persistent hypercholesterolemia. 3 In contrast, treatment with bone marrow cells from older ApoE-deficient mice with atherosclerosis was much less effective. This study demonstrates the role of competent EPCs in preventing atherosclerosis and the decreased effectiveness of senescent EPCs to engraft on recipient arteries in areas at risk for atherosclerotic injury. 3
Diabetes
Fadini and colleagues analyzed the association between type 2 diabetes, peripheral vascular disease, and levels of circulating EPCs using flow cytometry to quantify circulating progenitor cells (CPCs; CD34+) and EPCs (CD34+VEGF-R2+) in 51 diabetic patients and 17 control subjects. They reported that the numbers of CPCs and EPCs from diabetic patients were reduced by 33% and 40%, respectively, compared with healthy subjects. An inverse correlation was found between the number of EPCs and the values of fasting glucose. In addition, they determined that peripheral vascular disease was associated with a 47% reduction in circulating EPCs and that EPC number directly correlated with the ankle-brachial index (ABI). 27
In a second study, Fadini and colleagues quantified EPCs by flow cytometry in 127 diabetic patients with and without peripheral arterial disease (PAD). 28 Diabetic patients with PAD displayed a 53% reduction in circulating EPCs compared with non-PAD patients, and EPC levels were negatively correlated with the degree of carotid stenosis and claudication. In addition, the proliferative and adhesion capacity of cultured EPCs were significantly lower in diabetic patients with PAD versus patients without PAD. 28
Tepper and colleagues expanded on these findings and noted that proliferation of diabetic EPCs relative to control subjects was decreased by 48% (p < .01) and inversely correlated with patient levels of hemoglobin A1C (p < .05). 29 In addition, diabetic EPCs demonstrated decreased adherence and were 2.5 times less likely to participate in tubule formation compared with controls (p < .05).
Smoking
Kondo and colleagues examined the effects of chronic smoking and smoking cessation on EPC levels. 30 Circulating EPCs were quantified by flow cytometry in 14 nonsmokers and 15 smokers. The number of CPCs and EPCs was reduced in chronic smokers and inversely correlated with the number of cigarettes smoked. CPCs and EPCs increased rapidly after smoking cessation (p < .0001) and decreased again after resumption of smoking to the level similar to that before cessation (p = .0031), suggesting that EPC levels directly respond to the effects of smoking.
Michaud and colleagues studied peripheral blood EPCs in 15 healthy smokers and 11 age-matched nonsmokers. 31 The number of EPCs was significantly reduced in smokers versus control subjects, and the functional activities of EPCs isolated from smokers were severely compromised. The proliferative response was reduced by 75%, and the migratory response was reduced by 19% (p < .05). EPCs from smokers also showed decreased adherence and diminished capacity to form tubes in a Matrigel assay. They also found that EPCs from smokers had a significant reduction in the expression of endothelial cell–specific markers (VE cadherin, VEGF-R2, and vWF).
Positive Regulators
Exercise
It is generally accepted that exercise and physical training decrease the risk and severity of cardiovascular and peripheral vascular disease, but the molecular mechanisms for this protective effect have been elusive. Laufs and colleagues studied EPCs in mice randomized to running wheels or no exercise. 32 The number of EPCs circulating in the peripheral blood of trained mice were enhanced to 267%, 289%, and 280% of control levels after 7, 14, and 28 days, respectively. The researchers identified an exercise-induced nitric oxide (NO)-dependent mechanism that elevated serum levels of VEGF and reduced the rate of apoptosis in spleen-derived EPCs. In addition, running inhibited neointima formation after carotid artery injury by 22% and increased neoangiogenesis by 41% compared with controls.
Steiner and colleagues examined the effect of exercise on circulating EPCs in patients with cardiovascular risk factors and/or CAD. 33 Twenty patients with documented CAD and/or cardiovascular risk factors joined a 12-week supervised running program. After 12 weeks of exercise, there was a significant 2.9-fold increase in circulating EPCs, which was positively correlated with both the change in flow-mediated dilation and the increase in NO synthesis. Plasma VEGF and erythropoietin did not change in response to exercise. Thus, regular exercise training appears to augment the number of circulating EPCs in patients with cardiovascular risk factors.
Another study by Sandri and colleagues showed that ischemic exercise training in patients with PAD increased VEGF levels by 310% and the number of EPCs by 440% compared with controls. 34 However, subischemic exercise training in patients with PAD or CAD did not increase VEGF levels or EPC number, but it was associated with improved integrative capacity of progenitor cells to organize into endothelial networks and increased CXCR4 expression. In this study, ischemic exercise training increased EPC number and function, whereas nonischemic exercise appeared to improve only EPC function.
Hyperbaric Treatment
EPC trafficking is generally thought to be regulated by hypoxic gradients and induced by VEGF-mediated increases in bone marrow NO. Interestingly, Goldstein and colleagues found that hyperoxia induced by hyperbaric oxygen (HBO2) exposure paradoxically results in a signaling cascade similar to that induced by tissue hypoxia. 35 In an ischemic hindlimb model, the wound closure rate for a wound in the nonischemic limb did not significantly improve in response to HBO2 treatment, but a wound in the ischemic limb showed a significantly improved healing rate in the presence of HBO2. The researchers propose that HBO2 treatment increases EPC mobilization from the bone marrow but does not increase homing of EPCs into injured nonischemic tissues. A recent study by Gallagher and colleagues showed that HBO2 treatments increase nitric oxide synthase, which elevates NO in the blood. NO nitrosylates MMP-9, which cleaves membrane-bound stem cell factor, prompting EPC proliferation and migration. 36 Several studies demonstrate that HBO2 treatment mobilizes EPCs from the bone marrow and stimulates wound healing coincidently with the accumulation of EPCs in ischemic tissues. 35,37
Exogenous Drug Therapy
Many studies have investigated the effects of certain drug therapies for inducing recruitment, migration, proliferation, and incorporation of EPCs into target vascular epithelium. Many drugs have proven useful in this respect, both in vitro, in animal models, and in human clinical trials.
VEGF is a growth factor that is induced in hypoxic cells via HIF-1 and stimulates release of NO from endothelial cells. VEGF increases vascular permeability and exhibits a dose-dependent mitogenic and chemotactic effect on endothelial cells to promote new vessel formation. 38 VEGF is correlated with rapid but transient EPC recruitment after traumatic vascular injury 4 but has been tested more extensively in the context of therapeutic angiogenesis; initial hindlimb ischemia studies showed a dose-dependent increase in neovascularization and improvement in the hemodynamic deficit of the animal model. 38,39
However, VEGF is not entirely the answer. The Regional Angiogenesis with Vascular Endothelial Growth Factor (RAVE) trial was a randomized study of adenoviral vascular endothelial growth factor (AdVEGF) gene transfer for the treatment of PAD. One hundred five patients with unilateral exercise-limiting intermittent claudication were randomized to receive low-dose AdVEGF, high-dose AdVEGF, or placebo, administered as 20 intramuscular injections to the index leg in a single session. The change in peak walking time, ABI, claudication onset time, and quality-of-life measures were similar among all three groups at 12 and 26 weeks. In addition, AdVEGF administration was associated with increased peripheral edema, consistent with its known effect of increasing vascular permeability. The researchers were forced to conclude that VEGF-based treatment had limited utility since it was not associated with improved exercise performance or quality of life. 40
G-CSF, a growth factor for neutrophils, interferes with the CXCR4-SDF-1 receptor–ligand interaction, mobilizing HSCs into the bloodstream. Degradation of SDF-1 in the bone marrow releases EPCs by disrupting the CXCR4-SDF-1 receptor–ligand interaction; upregulation of SDF-1 in peripheral tissues attracts EPCs from the bone marrow to the periphery, particularly to sites of tissue hypoxia and HIF-1 expression. In this way, the CXCR4-SDF-1 receptor–chemokine interaction is instrumental for mobilizing, recruiting, homing, and incorporating EPCs to sites of vessel injury and ischemia. Notably, the CXCR4-SDF-1 interaction has been shown to simultaneously recruit bone marrow–derived SMC progenitors to regions of vessel injury where medial SMCs undergoing apoptosis express SDF-1. SMC precursors may assist in vascular repair; however, this response has also been shown to result in pathologic healing and neointimal hyperplasia. 41
Known primarily for their lipid-lowering activity, hydroxy-3-methylglutaryl coenzyme A reductase inhibitors increase neovascularization by increasing circulating EPC number and/or augmenting EPC function. Vasa and colleagues studied 15 patients with angiographically documented stable CAD to determine the effect of statin therapy on circulating EPC number. 42 These patients were prospectively treated with 40 mg of atorvastatin per day for 4 weeks. Statin treatment was associated with an approximately 1.5-fold increase in the number of circulating EPCs after 1 week, followed by sustained increased levels to approximately 3-fold during the 4-week study period. Walter and colleagues demonstrated that statins increased circulating rat EPCs by 2.5-fold at 4 weeks. 43 Dimmeler and colleagues used a mouse model to show that statins potently augment EPC differentiation from mononuclear cells and CD34-positive HSCs isolated from peripheral blood. 44 Interestingly, statins, VEGF, erythropoietin, estrogen, and exercise all exert effects on EPCs via the PI3K/Akt signal transduction pathway. This observation suggests an essential role for Akt in regulating hematopoietic progenitor cell mobilization, which is likely mediated through the eNOS pathway. 45
EPC and Peripheral Vascular Disease
Most studies evaluating the effectiveness of stem cell therapy use unpurified bone marrow containing a combination of EPCs, HSCs, and other cells. Asahara and colleagues originally decribed the incorporation of autologous CD34+ mononuclear cells into foci of neovascularization in the rabbit hindlimb ischemia model. 8 Kalka and colleagues confirmed these studies using human cells in a nude mouse model and found that blood flow recovery and capillary density in the ischemic hindlimb were markedly improved and the rate of limb loss was significantly reduced. The rate of limb necrosis and autoamputation was reduced by 50% compared with controls. 13,46
In addition to autologous transplantation of EPCs, certain factors can be administered or upregulated to help augment EPC function and increase ischemic limb perfusion. Takeshita and colleagues demonstrated a significant dose-dependent augmentation in ischemic limb perfusion accompanied by increased collateral formation after intramuscular administration of VEGF.
39
Murohara and colleagues showed a significant role for eNOS and NO in revascularization of ischemic limbs using
Tateishi-Yuyama and colleagues investigated the efficacy of autologous implantation of BMCs, including EPCs, into the ischemic limbs of patients with PAD. Twenty-two patients with bilateral leg ischemia were injected with BMCs in one leg and peripheral blood–derived mononuclear cells in the other. At 4 weeks, the ABI was significantly improved in legs injected with BMCs compared with those injected with peripheral blood mononuclear cells. Similar improvements were seen for transcutaneous oxygen pressure, rest pain, and pain-free walking time. These improvements were sustained at 24 weeks. The authors concluded that autologous implantation of BMCs could be safe and effective for achievement of therapeutic angiogenesis because of the natural ability of marrow cells to supply EPCs and to secrete various angiogenic factors or cytokines. 48 Esato and colleagues demonstrated similar results in patients with PAD who had failed traditional medical treatment and/or surgical bypass procedures. 49 After BMC transplantation, patients reported improved subjective symptoms.
In contrast to the studies evaluating BMC therapy, Ishida and colleagues used peripheral blood mononuclear cells in patients with PAD. 50 These cells were mobilized with G-CSF and then harvested and injected intramuscularly. After 4 weeks, the researchers observed improvement in ABI, healing of ischemic ulcers, and increased mean maximum walking distance. As a group, these studies suggest that the specific cell type, or the combination of cells and cytokines, necessary and sufficient for generating beneficial healing effects in patients with PAD is still incompletely understood.
Bone Marrow Cells and Graft Patency
Although studies have not been performed with isolated EPCs, several studies have shown that some bone marrow–derived stem cells can colonize and epithelialize synthetic and biologic vascular grafts and devices to render the foreign surface less thrombogenic. Shi and colleagues used a canine bone marrow transplantation model in which the marrow cells from the donor and recipient were genetically distinct. 18 Between 6 and 8 months after transplantation, a Dacron graft, made impervious to prevent capillary ingrowth from the surrounding perigraft tissue, was implanted in the descending thoracic aorta. After 12 weeks, the graft was retrieved, and cells with endothelial morphology were identified by silver nitrate staining. Using repeat polymorphisms to distinguish between the donor and recipient deoxyribonucleic acid (DNA), Shi and colleagues observed that only donor alleles were detected in DNA from positively stained cells on the impervious Dacron graft. These results strongly suggest that a subset of CD34+cells localized in the bone marrow can be mobilized to the peripheral circulation and can colonize endothelial flow surfaces of vascular prostheses.
In a similar study, Bhattacharya and colleagues demonstrated accelerated endothelialization on polyethylene terephthalate (PET) grafts treated with enriched CD34+autologous bone marrow cells in a canine model. 51 The authors used composite grafts implanted in the dog's descending thoracic aorta for 4 weeks. The composite grafts had a 4 cm PET graft in the center flanked by two 4 cm standard polytetrafluoroethylene grafts; the entire composite was coated with silicone rubber to make it impervious, and the PET segment was shielded from perigraft and pannus ingrowth. On the day before surgery, 120 mL bone marrow was aspirated and CD34+ cells were enriched. During surgery, these cells were mixed with venous blood and seeded onto the PET segment of composite study grafts; the control grafts were treated with venous blood only. After harvesting, there was a significant increase in surface endothelialization on the seeded grafts (approximately 92% vs 27%), with markedly increased microvessels in the neointima, graft wall, and external area compared with controls.
In humans, Peichev and colleagues found that the neointima formed on the surface of left ventricular assist devices was colonized with cells expressing the markers CD34, CD133, and VEGF-R2. 20 Furthermore, Matsuo and colleagues showed that senescent EPCs may be a risk factor for in-stent restenosis in a study of 46 patients who underwent coronary stenting. 52 Blood samples were collected at the time of follow-up coronary angiography after stent placement. Patients (n = 16) with in-stent restenosis, defined as > 40% stenosis, had decreased number of EPCs and increased senescent cells compared with patients without restenosis (n = 30). There was no significant difference in angiogenic growth factors secreted by EPCs between the two groups. On multivariate analysis, increased number of senescent EPCs was the independent factor associated with in-stent restenosis (odds ratio 1.10).
Shin'oka and colleagues pioneered the exciting field of tissue-engineered biodegradable grafts seeded with autologous bone marrow cells for the repair of cardiovascular defects in children. 53 Previous studies showed that bone marrow cells contribute to the construction of tissue-engineered vascular autografts (TEVAs) in vivo, and given that these constructs contain living cells, they may have the ability to grow, self-repair, and self-remodel. These characteristics are especially desirable in growing children to decrease the number of revision surgeries. Biodegradable conduits (n = 23) and patches (n = 19) were implanted in children for extracardiac total cavopulmonary connection and congenital heart defects, respectively. TEVAs were implanted only in low-pressure (venous or pulmonary artery) systems. Patients received anticoagulation therapy for 3 to 6 months postoperatively. Intermediate-term follow-up after a median of 16.7 months showed no complications, such as thrombosis, stenosis, or obstruction of TEVAs. There was no evidence of aneurysm formation or calcification. All tube grafts were patent, and the diameter of the tube grafts increased to approximately 110% of the implanted size over time. These results show that after intermediate-term follow-up, TEVAs are safe and feasible in pediatric cardiovascular surgery and have excellent hemodynamic performance without anticoagulation therapy. 53–55 However, the implications for adult patients with arterial disease are not clear.
EPC Therapy
Therapy that targets the recruitment, mobilization, proliferation, and incorporation of EPCs into injured vessels and ischemic tissues augments healing and tissue regeneration in animal models and in patients with severe vascular disease, as described above.
As these therapies become established, three categories of patients may be considered for therapy with EPCs. The first category includes patients with severe ischemic peripheral vascular disease, a threatened limb, and no surgical or endovascular options for reconstruction. The second category includes patients with severe comorbid disease who are not operative candidates for potential procedures. The third category includes patients with bypass grafts or stents in place; under these circumstances, EPC therapy may improve conduit survival by preventing restenosis. 13 However, EPC therapy has the potential to stimulate neovascularization in tumors and therefore is not indicated for patients with or at high risk for cancer. 56,57
Another important consideration in EPC therapy is the possibility of resultant neointimal hyperplasia after arterial injury owing to the contribution of SMCs stimulated by SDF-1. Zernecke and colleagues described the involvement of local SDF-1α and its receptor CXCR4 in neointimal hyperplasia via recruitment of bone marrow–derived SMC progenitors. After arterial injury, SDF-1α expression in medial SMCs is preceded by apoptosis; SDF-1α binds to platelets at the site of injury, triggers CXCR4- and P-selectin-dependent arrest of progenitor cells on injured arteries or matrix-adherent platelets, and preferentially mobilizes and recruits progenitors for neointimal SMCs. 41
In patients who are selected as candidates for EPC therapy, several methods for augmentation of EPC number and function exist. Strategies include improving the intrinsic function of a patient's native cells, processing of precursor cells, providing allogeneic progenitors, or using a combination of these approaches.
Most studies in which EPCs were used to treat patients with vascular disease used either unpurified bone marrow samples or a combination of EPCs, HSCs, and other cells derived from bone marrow. Thus, it is unclear whether EPCs alone are sufficient to repair vascular injury or whether other supporting cells or components of the bone marrow are also necessary for a therapeutic effect. This is a critical point, as introduction of nonessential cells may be associated with aggravation of inflammation and vascular injury, thereby exacerbating the problem for which the patient seeks treatment. 11
Conclusion
Recent studies demonstrate the correlation between improved quality and/or quantity of EPCs with reduced risk for vascular disease. Preclinical and clinical trials also demonstrate the beneficial effects of EPC therapy. Understanding the mechanism of EPC function and the optimal cell type and delivery methods are topics of active research.
Footnotes
This work was supported by a Doris Duke Clinical Research Fellowship, an American Vascular Association Lifeline Student Research Fellowship, a National Institutes of Health Career Development award (HL079927), and the American Vascular Association William J. von Liebig Award, as well as the resources and use of the facilities at the VA Connecticut Healthcare System, West Haven, CT.