
Abstract
Objective:
The aim of the present study was to check the potential interaction of two neurodevelopmental proteins, Disc1 and Gas7, in the adult mice brain.
Methods:
Twenty-four male Swiss albino mice were used for the study. The mice were 12 weeks old in the beginning of the experiment. Immunohistochemistry and co-immunofluorescence were performed on the coronal sections of mice brain and immunoblotting and co-immunoprecipitation were done on the whole brain lysate.
Results:
Data from immunohistochemistry and co-immunofluorescence indicate the occurrence and co-localization of Disc1 and Gas7 proteins in soma and projections of the brain cells. Immunostaining was observed in cerebral cortex, hypothalamus, midbrain, pons, medulla oblongata and CA3 of hippocampus of the brain. The data from Immunoblotting and co-immunoprecipitation validates the presence and interaction of Disc1 and Gas7 protein in whole brain lysate.
Conclusion:
Data indicates the potential interaction of Disc1 and Gas7 protein in adult brain. The study highlights the need for further research on Disc1–Gas7 protein interaction in brain development and neuro-disorders.
1 Introduction
Disrupted-in-schizophrenia 1 (Disc1) gene was originally identified in a Scottish family where balanced translocation between chromosomes 1 and 11 caused the disruption of Disc1 on chromosome 1. The event resulted in a major cognitive disorder associated with learning and memory [1]. Later, reports on Disc1 established a crucial role in neuro developmental process that includes neurite outgrowth, neuronal migration and synapse formation [2–4]. The role of Disc1 in neurodevelopment was found due to the interaction of Disc1 with other binding proteins called the Disc1 interactomes [5]. In the year 2003, Morris et al. identified protein binding motifs on Disc1 and suggested Disc1 could be a scaffold protein for the interaction with other proteins [6]. Understanding the Disc1 interactome proteins would help in knowing the neuro developmental process and identify therapies for neuro developmental disorders.
Growth arrest specific gene 7 (Gas7) was initially identified in cultured cells that had entered a quiescent state induced by serum starvation [7]. In addition to the expression in response to growth arrest, Gas7 protein was shown to involve in a variety of biological process including, neurite extension and neuronal migration [8, 9]. The Gas7 requirement in neuronal differentiation was demonstrated in cultured neurons [8], where Gas7 overexpression induced the formation of membrane protrusions in Neuro2a cells [9] and inhibition prevented neurite outgrowth in Purkinje cells [8]. Gas7 is considered as a highly conserved among species and is an important protein from the evolutionary standpoint [10, 11]. Till date, all the research on Gas7 points the biological significance of this protein in neuronal development, therefore suggesting the worthy of Gas7 in further investigation.
The current study focuses on the potential interaction of Disc1 and Gas7 proteins in adult mice brain. The interaction of Disc1 and Gas7 proteins are very important to understand, since this would help in unfolding many complex network of reactions linked to the regulation and execution of cellular functions and biological processes. To the current knowledge, this is the first study on the Disc1 and Gas7 protein interaction at microscopy and biochemistry level.
2 Materials and methods
2.1 Animals
A total of twenty-four male Swiss albino mice were used for the study. Twelve-week-old mice were obtained from the Kings Institute of Preventive Medicine, Chennai, India. They were given ad libitum food and water, and were housed four per cage in a temperature-controlled environment at 22°C, with an artificial 12-12-hour light-dark cycle. Eight mice were used for immunohistochemistry, eight for double immunofluorescence and eight for immunoblotting and co-immunoprecipitation. All the experimental procedures were approved by the Institutional Animal Care and Use Committee at University of Madras, India.
2.2 Primary Antibodies
Rabbit anti-Gas7 antiserum was generated against histidine-tagged full-length Gas7 protein expressed in E. coli from plasmid pET15b (gift from Dr. Sue Lin-Chao, Academia Sinica, Taiwan, China). Disc1 rat polyclonal antibody used in this study was from Spring Valley Laboratories, Inc., Woodbine, MD. (kind gift from Dr. Mikhail V. Pletnikov, Johns Hopkins University, USA).
2.3 Immunohistochemistry
The procedure for immunohistochemistry was performed as per the protocol described in the previous publication [12]. In specific, mice were euthanized with sodium pentobarbital (100 mg/kg) and perfused with ice-cold phosphate-buffered saline (PBS), followed by 4% paraformaldehyde. Brains (n = 8) were removed, fixed in para-formaldehyde and paraffin embedded brain samples were cut coronally at 8 μm thickness. Representative brain sections were dewaxed in xylene and rehydrated in graded alcohol. Sections were incubated in 3% hydrogen peroxide for 15 min and nonspecific binding was blocked by incubation with 3% bovine serum albumin in PBS for 1 h. They were later replaced with Disc1 or Gas7 anti-serum (dilution 1:250 and 1:200) and incubated overnight at 4°C in a saturated humidity chamber. Antibody binding was demonstrated using biotinylated secondary antibody (Cat. # ab6733, Cat. # ab97049, Abcam, Cambridge, United Kingdom) and 3,3’-diaminobenzidine as the chromogenic substrate, counterstained with Mayer’s haematoxylin. Sections were dehydrated, mounted with DPX, and examined using Axioskop-2 Plus Phase Contrast Microscope (Code: SKU-012, Carl Zeiss, Munich, Germany). Negative controls for tissue sections consisted of substituting the primary antibody with 2% bovine serum albumin in PBS.
2.4 Co-immunofluorescence
Brains (n = 8) postfixed in 4% paraformaldehyde were transferred to 30% sucrose for cryoprotection. After refrigeration in 30% sucrose solution for 48 h, brains were frozen and sectioned at 40 μm thickness. Free-floating sections were incubated in 3% bovine serum albumin to reduce nonspecific background. Sections were incubated with anti-Disc1 antibody, 1:400, for 3 h at room temperature, washed in PBS and then again incubated with anti-Gas7 antibody, 1:300 for 3 h. They were washed in PBS and then stained for 1 h at room temperature with Alexa Fluor 350-conjugated anti-rat secondary antibody (Cat. # A21093, Invitrogen, Waltham, MA, USA), diluted 1:800 in PBS. They were washed again and stained with Alexa Fluor 594-conjugated anti-rabbit secondary antibody (Cat. # A32740, Invitrogen, Waltham, MA, USA), dilution 1:600. After washed in PBS, sections were mounted using Vectashield mounting medium (Cat. # H-1300, Vector Labs, Burlingame, CA, USA). Images were collected on Axioskop-2 Plus Phase Contrast Microscope (Code: SKU-012, Carl Zeiss, Munich, Germany) and single-channel images were overlaid in Adobe photoshop package (Version: CS6, San Jose, CA, USA) to create a merged image file. Negative controls consisted of substituting the primary antibody with 2% bovine serum albumin in PBS.
2.5 Immunoblotting
Whole brain from adult mouse was homogenized in ice cold RIPA buffer (Cat. # R0278, Sigma-Aldrich, St. Louis, MO, USA) containing protease and phosphatase inhibitors (Cat. # 5892970001 and 4906845001, Roche, Basel, Switzerland) and centrifuged at 15,000 g for 20 min. The protein level in the supernatant was determined using Micro BCA protein assay kit (Cat. # 23235, Thermo Scientific, Waltham, MA, USA) with bovine serum albumin as the standard. Lysate was boiled with SDS sample buffer for 5 min, subjected to SDS-polyacrylamide gel electrophoresis (PAGE) and transferred to the polyvinylidene fluoride (PVDF) membrane. After blocking with 5% milk-Tris-buffered saline-Tween, the membrane was incubated with the primary antibody for 12 h at 4°C. For the detection of Disc1 or Gas7, antibodies raised against these proteins were used at 1:800 and 1:1000 dilutions respectively. The membrane was then incubated with corresponding secondary IgG, HRP-linked antibody (Cat. # ab97057, Cat. # ab6721, Abcam, Cambridge, United Kingdom) at 1:5000 dilution for 1 h at room temperature. Immunoblotting was visualized by chemiluminescence using the ECL kit (Cat. # RPN2232 and 28906835, Amersham Biosciences, Amersham, United Kingdom).
2.6 Co-immunoprecipitation
Individual whole brain lysate (100 μg of protein) was subjected to immunoprecipitation with 5 μg of anti-Disc1 antibody. Immunocomplex was allowed to form overnight at 4°C with gentle rocking. Following an overnight incubation, sample was loaded with 30 μL of Protein A agarose bead slurry and gently rocked for 3 h at 4°C. Sample was then centrifuged for 30 s, washed, and 20 μL of SDS sample buffer was added. It was then heated at 100°C for 5 min and 30 μL of sample was loaded onto a 10% SDS-PAGE resolving gel. Following electrophoresis, resolved protein was transferred onto PVDF membrane, blocked, and then probed with anti-Gas7 primary antibody for 12 h at 4 °C. Signal was visualized using the ECL kit.
3 Results
3.1 Localization of Disc1 and Gas7 protein in adult mice brain
To determine Disc1 and Gas7 protein expression, immunohistochemistry was performed. The immunostaining was performed on 8 individual brain samples. Disc1 and Gas7 staining showed a similar labeling with general distribution throughout the brain in all the 8 samples. Although in certain areas such as cerebral cortex, hypothalamus, midbrain, pons, medulla oblongata and CA3 of hippocampus the labeling was more evident and intense (Figs. 1 and 2). Magnified images of the immunostaining revealed that in cerebral cortex, the Disc1 and Gas7 staining was more concentrated in cell processes [Figs. 1(b) and 2(b)]. In hippocampus, Disc1 and Gas7 immunoreactivity was high in the pyramidal cells of CA3 region [Figs. 1(l) and 2(l)]. In hypothalamus, midbrain, pons and medulla oblongata, the cells were highly positive for Disc1 and Gas7 proteins. In these areas, Disc1 and Gas7 were concentrated both in cell bodies and projections [Figs. 1(d), (f), (h), (j) and Figs. 2(d), (f), (h), (j)].
3.2 Co-immunolocalization of Disc1 and Gas7 protein in adult mice brain
To determine Disc1 and Gas7 protein interaction, co-immunofluorescence was performed on adult mice brain sections. Immunofluorescence signal for Disc1 and Gas7 protein was scattered throughout the brain tissue in all the individual mice brain samples (n = 8). Though the overlap of Disc1 and Gas7 was noticed in a few cells in cerebral cortex and CA3 of hippocampus, the signal was more evident in midbrain and medulla oblongata. The co-immunofluorescence was distinct in the cell body in these brain areas [Figs. 3(c), (d), (g), (h)].

Disc1 localization in adult mouse brain. Immunohistochemistry of Disc1 in the coronal section of adult mouse brain show Disc1 labeling throughout the section, with Disc1 staining being more evident in specific areas: cerebral cortex (a, b), hypothalamus (c, d), midbrain (e, f), pons (g, h), medulla oblongata (i, j) and CA3 of hippocampus (k, l). Photograph in the right-hand panel (b, d, f, h, j, l) show enlarged view of boxes on the left-hand panel (a, c, e, g, i, k). The sections were counterstained with Mayer’s haematoxylin. Disc1 labeling is especially evident in cell bodies (arrows) and in projections (arrows). Scale bars: 200 μm (a, c, e, g, i, k), and 50 μm (b, d, f, h, j, l).
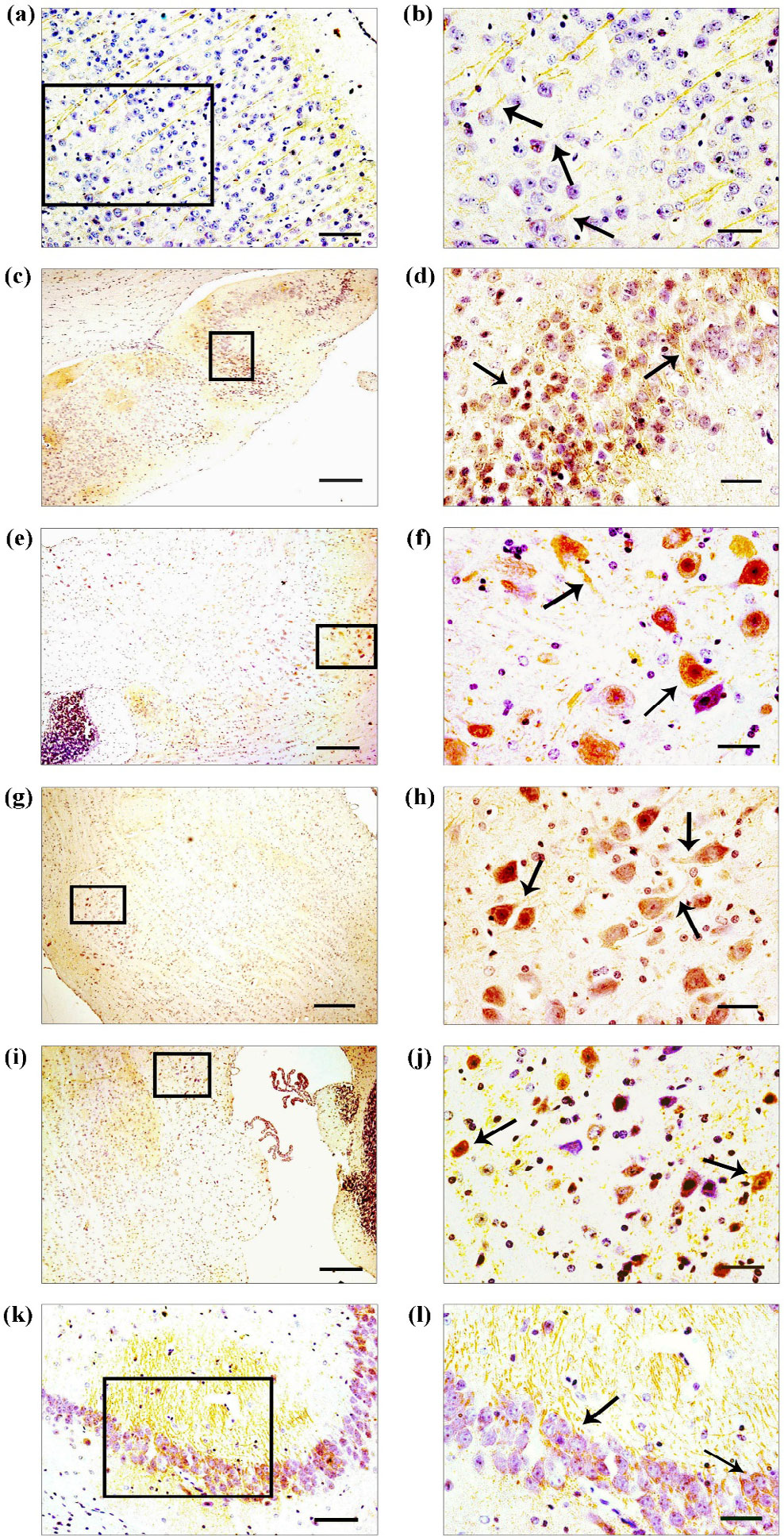
Gas7 immunololocalization in adult mouse brain. Gas7 protein distribution is represented on coronal sections of mouse brain. Gas7 protein was differentially expressed within brain regions. Gas7 was strongly expressed in the: cerebral cortex (a, b), hypothalamus (c, d), midbrain (e, f), pons (g, h), medulla oblongata (i, j) and CA3 of hippocampus (k, l). The right-hand panel has enlarged photographs to show the staining pattern in cell bodies (arrows) and in projections (arrows). Scale bars: 200 μm (a, c, e, g, i, k), and 50 μm (b, d, f, h, j, l).
3.3 Detection and co-immunoprecipitation of Disc1 and Gas7 protein in mice brain lysate
Immunoblot and co-immunoprecipitation was performed on eight individual brain samples. Immunoblot analysis of brain lysate showed the presence of 97 kDa Disc1 [Fig. 4(a)] and 48 kDa Gas7 protein [Fig. 4(b)] in all the samples. The data verifies the result obtained from immunohistochemistry where Disc1 and Gas7 proteins were distributed in multiple brain areas including cerebral cortex, hypothalamus, midbrain, pons, medulla oblongata and hippocampus. To validate the data obtained from co-immunofluorescence, co-immunoprecipitation was performed. The experiment successfully demonstrated the Disc1 and Gas7 co-immunoprecipitation in all the eight brain samples [Fig. 4(c)].

Co-immunofluorescence analysis of Disc1 and Gas7 in coronal sections of adult mouse brain. Co-immunofluorescence labeling show that Disc1 and Gas7 co-localize in the mouse brain tissue. The co-localization signal seen in midbrain (c, d) and in medulla oblongata (g, h). Figure (d) and (h) are the enlarged image of figure (c) and (g) to show Disc1 and Gas7 co-localization in cell body (arrows) and in cell projection (arrows). Scale bars: 200 μm (a, b, c, e, f, g), and 50 μm (d, h).
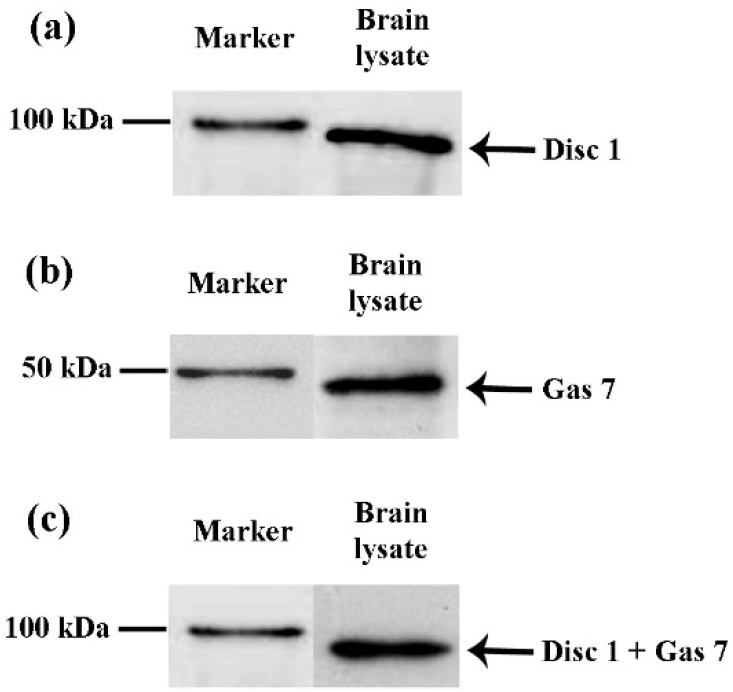
Disc1 and Gas7 protein expression and co-immunoprecipitation in adult mouse brain. The immunoblot analysis of brain lysate show the presence of 97 kDa Disc1 (a) and 48 kDa Gas7 protein (b) in mouse brain. Co-immunoprecipitation analysis of mouse brain lysate display 97 kDa band, suggesting a strong preference of Disc1 with Gas7 (c). The blots were cropped and were positioned to the protein marker to determine protein mass.
4 Discussion
Previous studies have demonstrated the independent expression of Disc1 and Gas7 protein in rodent brain [8, 12, 13]. But to-date, no studies have shown their co-expression in brain tissue. In this study, using antibody specific for Disc1 and Gas7 the co-expression of these proteins was studied in the adult mice brain.
Disc1 and Gas7 proteins are known to express in brain cells of different areas and during the various stages of brain development [8, 13]. These proteins are shown to interact with cytoskeletons including the actins and microtubules and plays a crucial role in neurogenesis [14 –16]. The Disc1 and Gas7 proteins are known to involve in neuronal outgrowth, both in vitro and in vivo [6, 7, 10, 13]. Loss of function of these proteins are shown to affect neuronal differentiation and migration of neurons [8, 17]. Studies point that neurodevelopmental proteins, in specific, fasciculation and elongation protein zeta-1 (Fez1), platelet-activating factor acetylhydrolase, isoform Ib, PAFAH1B1 or lissencephaly 1 protein (Lis1), nuclear distribution element-like (Nudel) interact with Disc1 [6, 17]. Similarly, WISH (WASP interacting SH3 protein), Neural-Wiskott Aldrich syndrome protein (N-WASP), tau protein interacts with Gas7 [18, 19], and assist in neuronal differentiation. The results from current co-immunolocalization indicate that Disc1 and Gas7 co-localize in a functionally relevant neuronal subtype in mice brain. The co-immunoprecipitation data supports the result from co-immunolocalization where the co-immunoprecipitation of Disc1 and Gas7 were observed in the brain tissue lysate. Studies by Morris et al. [6] and Shimizu et al. [19] indicate that, Disc1 and Gas7 protein binding partners are specific to nervous system. Ju et al. [8] and Schurov et al. [13] points that Disc1 and Gas7 are associated with the neuronal development. Together, along with the current results from co-immunolocalization and co-immunoprecipitation study, it can be specified that Disc1 and Gas7 may physiologically interact in the brain tissue.
Previous research has indicated the similarity in role of Disc1 and Gas7 in binding to the cytoskeletons and aid in neuronal growth and development. Results from current immunohistochemistry study points the similarity in staining pattern of Disc1 and Gas7 in various brain areas and in brain cells including the projections and soma of, cerebral cortex, hypothalamus, midbrain, pons, medulla oblongata and CA3 of hippocampus. The resemblance in staining pattern of Disc1 and Gas7 in these brain areas indicate that these proteins may interact with each other in brain cells.
Millar et al. in 2000 [1] have reported that Disc1 encode an 854 long amino acid protein with N-terminal globular domain and a C-terminal coiled-coil region. Similarly, Gas7 has been shown to express 320 amino acid residues in the C-terminal region with an FCH and coiled-coil domain [18]. Mason and Arndt in 2004 [20] have reported that proteins with coiled-coil domain in C-terminus interact with each other and play an important role in cellular growth and development. Disc1 and Gas7 protein plays a significant role in neuronal growth and development. These proteins share a similarity in protein structure, with coiled-coil domain in the C-terminus and interact with other proteins associated to cell development. The findings suggest that Disc1 and Gas7 may interact with each other during the brain development.
Aberrant expression of Disc1 and Gas7 have been shown to cause various neurological disorders. In specific, both are linked to Alzheimer’s and schizophrenia [2, 21 –24]. The association of Disc1 and Gas7 to a common neurodevelopmental disorder indicate that these proteins may share common root during the neuropathology process. The findings emphasize a likelihood of Disc1–Gas7 protein interaction in vivo. A detailed investigation of Disc1–Gas7 protein interaction is needed to further understand its role in brain development and neuro disorders.
There are some limitations to this study. For future research, a larger sample size should be used for each experimental group to better validate the hypothesis. Since this is the first to identify the potential interaction of Disc1 and Gas7 proteins in adult brain, further molecular and biochemical approaches have to be done to confirm the functional interaction of these proteins at any biological conditions.
Footnotes
Ethical approval
All the experimental procedures were approved by the Institutional Animal Care and Use Committee at University of Madras, India.
Conflict of interests
The author declares no conflict of interests.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Acknowledgments
The author thanks Divya Chandrababu and Vahin Uday for their support.