
Abstract
Worldwide, ocular cataracts are a major cause of human blindness. A key goal of cataract-related research is to identify simple, cost-efficient but effective ways to prevent cataract formation or progression. Genistein is a naturally occurring dietary isoflavone with well-documented estrogenic, antioxidant, and protein tyrosine kinase inhibitor activity, which in turn modulates the activity of several enzymes involved in cell signaling and proliferation. Furthermore, many isoflavones have been shown to be potent inhibitors of aldose reductase, which is an important rate-limiting enzyme in the process of cataract induction in the metabolic disease galactosemia. In order to assess the potential for genistein to mitigate cataract formation, we have studied its effects in the animal model of dietary galactose-induced cataracts in adult male rats. Our experimental hypothesis was that dietary genistein would prevent or delay the progression of cataracts induced by high dietary intake of galactose. Our results show that the isoflavone genistein was not able to completely prevent galactose-induced cataract formation, but genistein did delay the progression of cataracts induced by dietary galactose. In addition, we found that dietary galactose decreased concentrations of serum somatostatin, while adding genistein decreased the serum glucose level but increased the serum testosterone level. As an initial inquiry into the mechanisms by which the partial protective effect of genistein could be mediated, we found that genistein increased the expression of connexin (Cx) 43 in the lens but did not affect the expression of soluble guanylyl cyclase (sGC) subunits. This finding suggests that the partial protective effect of genistein on cataract induction appears to be unrelated to sGC but may be mediated by enhanced expression of Cx43 and changed metabolic state.
Introduction
Cataract is a major ocular disease that causes blindness in the world (1). It is well established that various factors such as oxidative stress, UV, and other toxic agents can induce both in vivo and in vitro cataract formation (2–5). Blindness due to opacity of the lens, or cataract, afflicts 50 million persons worldwide (1). In the United States, over 541,000 cataract extractions are done annually at a cost of over $3.8 billion (6). Conservative estimates indicate that the prevalence of cataracts in Americans aged 65–75 and 75–85 years are 18% and 46%, respectively (7). Cataracts are even more prevalent in some other countries (8). It is estimated that the cost for cataract extractions would diminish by half if onset of cataract could be delayed by only 10 years (6). Thus for both human health and health-care cost considerations, identification of any simple, cost-efficient but effective way to prevent cataract formation or progression could have a profoundly beneficial impact.
The toxicity of galactose on the lens has been studied for nearly 70 years, and galactose-induced cataract is an accepted animal model for the investigation of the cataracts in humans (9–12). Legumes contain isoflavones that are known to have pharmacologic properties in mammals (13, 14). One such isoflavone, genistein, is well demonstrated to act as an estrogen-receptor ligand, to have antioxidant properties, and to be an inhibitor of several key enzymes such as protein tyrosine kinases involved in cell signaling and proliferation (15, 16). Thus the aim of our current study was to investigate the effects of genistein on galactose-induced cataracts in rats.
Although there are many reports on induction and inhibition of cataract (17, 18), the relevant molecular and cellular mechanisms are still unclear. Two plausible mechanisms are changes in gap junction formation and function (19) and changes in soluble guanylate cyclase (sGC) (EC 4.6.1.2) expression (20).
Gap junctions are composed of the connexins, a family of proteins, which have been shown to be the structural and functional building blocks of gap junctional intercellular channels (21, 22). Connexin (Cx) 43 has been identified as the major connexin of lens epithelial cells. Osmotic balance within the lens is markedly altered in Cx43 (−/−) animals, highlighting the importance of intercellular communication mediated by lens epithelial Cx43 gap junctions in the function of this tissue (23, 24). Grummer et al. (1994, 1999) reported that the expression of connexin genes, Cx26 and Cx43, is regulated by ovarian steroid hormones in the rat endometrium, but not in heart (Cx43) and liver (Cx26) tissue of the same animals (25, 26). This finding raises the question about tissue-specific regulation properties of connexin genes. Genistein is known to bind to the estrogen receptor (27). Therefore, in our current study, we have investigated whether expression of Cx43 is involved in the action of genistein in this animal model of cataract.
Soluble guanylate cyclase is a receptor component for the cyclic guanosine monophosphate (cGMP)-dependent signaling system in various cells, including vascular smooth muscle cells and neurons (28, 29). This enzyme is a heme protein composed of two subunits and is able to bind nitric oxide (NO) and carbon monoxide (CO) to increase their activity (30). NO has been implicated as a key regulator of synaptogenic events in a number of systems, most convincingly in the developing visual system (31, 32). Although there are several reports about sGC expression in the eyes (33–35), we know of no data addressing the possibility that sGC might be involved in cataract induction or prevention. Thus our current study also investigated whether the NO/sGC/cGMP signaling system was changed in cataract lenses.
Materials and Methods
Animals.
Eighteen male Long-Evans rats (25 days old) were obtained from the Cedars-Sinai Vivarium (Los Angeles, California) and pair-housed in air-conditioned quarters with food and water available ad libitum. The lighting schedule was 12:12-hr light:dark cycle with lights on 0500 hrs through 1700 hrs. The Cedars-Sinai Vivarium is accredited by the American Association for Laboratory Animal Care. All animal procedures were conducted in accordance with the Institutional Animal Care & Use Committee (IACUC) approved protocol.
Diets.
To avoid dietary exposure to isoflavones during the experiment, AIN93G (7% corn oil diet) was used as the control diet. The galactose-rich diet was iso-caloric and iso-protein to AIN93G but contained 20% galactose. The diets were prepared by Harlan Teklad (Madison, WI).
Antibody.
Monoclonal connexin-43 antibody (Product C8093, Lot 28H4827), anti-rabbit sGC alpha 1 serum (Product G4280, Lot 011K4888), and beta 1 serum (Product G4405, Lot 011k4880) were purchased from Sigma-Aldrich Inc. (St. Louis, MO).
Experimental Treatment.
The male rats were randomly divided into three groups of six rats each as follows: Group 1, control; Group 2, galactose only; and Group 3, galactose + genistein. For the controls (Group 1), rats were fed AIN93G, while Group 2 and Group 3 were fed the galactose-rich diet. All rats were gavaged daily over the experimental treatment period of 4 weeks. Control (Group 1) and galactose only (Group 2) animals were given 0.5 ml corn oil alone, while Group 3 rats were given 0.5 ml corn oil containing genistein (15 mg/kg body weight). The progression of cataracts was determined by slit lamp check before rats were necropsied. Cataracts in every animal were scored and recorded by an experienced observer who was blinded to the identity of each individual animal at the time of the assessment (36–38). Cataract severity was graded on a decimal scale, and the standards had regularly spaced intervals on a decimal scale. Five grades were used in the current experiments, 5 indicates severe cataract and 0 indicates normal lens. These grades were as follows: Clear or grade 0, no vacuoles present or clear lens; Grade 1, vacuoles of less than one-fourth of the lens radius; Grade 2, vacuoles located at the periphery of the lens occupying an area of between one-fourth and one-half of the radius from the periphery; Grade 3, vacuoles extending between one-half and three-fourths of the radius from the periphery; Grade 4, vacuoles extending up to three-fourths of the radius from the periphery (nuclear opacity may be seen); Grade 5, vacuoles cover the entire lens, which appears white to the naked eye. The incidence of cataract appearance was expressed as the percentage of total lenses in each group. All eyes were then collected when rats were necropsied.
Eye Histology.
Following collection, one eye of each rat was fixed in 4% paraformaldehyde solution, and subsequently transferred to 70% ethanol after 24 hours and prepared for microscopic analysis using routine techniques. All eyes were sectioned at 5 μm and stained with hematoxylin and eosin (HE).
Immunohistochemistry.
Slides with 5-μm sections were deparaffinized in xylene and rehydrated in a graded ethanol series. Slides were transferred to Antigen Retrieval Citra (pH 6.0) (Biogenex, San Remon, CA) in plastic Coplin jars and heated for 5 mins in a microwave oven and then washed in phosphate-buffered saline (PBS). Nonspecific binding was blocked with 10% normal goat serum in PBS for 30 mins, followed by incubation with Cx43 monoclonal antibody (1:500), soluble guanylyl cyclase alpha (1:5000), or beta subunit (1:2000) antisera in 10% goat serum in PBS for 24 hrs at 4°C refrigerator or with negative control serum. Sections were then washed in Autobuffer (Biomeda Corp., Foster City, CA) four times for 1 min each, followed by incubation with the biotinylated secondary antibody for 30 mins. After Autobuffer washing four times for 1 min each, sections were incubated for 30 mins with alkaline phosphatase-conjugated streptavidin for 30 mins. Again, after Autobuffer washing four times for 1 min each, the localization of the primary antibody was performed by incubation with alkaline phosphatase substrate kit I (Vector Laboratory Inc., Burlingame, CA) for 10 min, producing a red-colored precipitate. Sections were then counterstained in Mayer’s hematoxylin, dehydrated through graded ethanol solutions, cleared in xylene, and mounted with permount. Relative levels of immunostaining between animals were evaluated by three independent observers with five eye samples per group. Results described represent consistently observed patterns of immunostaining, and representative photographs were taken. Images were acquired digitally using a microscope coupled with a Polaroid camera (Polaroid Corp., Waltham, MA). An image analysis system (Image Pro Plus 4.0, Media Cybernetics, Inc., Silver Spring, MD) was used to analyze the staining results.
Hormone Profiles.
Serum insulin and leptin concentration were determined by commercial double-antibody radioimmunoassay (RIA) kits (Linco Research Company, St. Charles, MO). A rat somatostatin RIA kit was provided by (Peninsula Laboratories Europe Ltd., St. Helena, UK). Serum testosterone was determined by RIA kit (Diagnostic Products Corp., Los Angeles, CA). Samples were used in duplicate for the RIA assay according to the instructions of the manufacturer. The following example of a leptin assay is provided to demonstrate the procedure of RIA hormone assay. Briefly, pipette 300 μl of assay buffer to the nonspecific binding (NSB) tubes Nos. 3–4; pipette 200 μl of assay buffer to tubes Nos. 5–6; pipette 100 μl of assay buffer to tubes 7 through the end of the assay; pipette 100 μl of standards, quality controls, and samples in duplicate; pipette 100 μl of rat leptin antibody to all tubes except total count tubes (Nos. 1–2) and NSB tubes (No. 3–4); vortex, cover, and incubate overnight (20–24 hrs) at room temperature; pipette 100 μl of 125I-rat leptin to all tubes; vortex, cover, and incubate overnight (22–24 hrs) at room temperature; add 1.0 ml of cold (4°C) precipitating reagent to all tubes except total count tubes (Nos. 1–2); vortex and incubate 20 mins at 4°C; centrifuge, at 4°C, for 20 mins at 2000–3000 g; immediately decant the supernate of all tubes except total count tubes (Nos. 1–2), drain tubes for at least 15–60 secs and blot excess liquid from lip of tubes; count all tubes in a gamma counter for 1 min. Finally, calculate the ng/ml of rat leptin in unknown samples using automated data reduction procedures. All analyses are performed within 2 months of sample collection.
Glucose Assays.
Concentrations of serum glucose were measured by the enzymatic methods using commercial kit (Sigma-Aldrich, St. Louis, MO). Briefly, dilute glucose standard and samples; then, transfer 1-ml diluted standards and samples to appropriately labeled tubes, next add 2.0 ml of assay reagent to each tube and incubate the tubes for 30 mins at 37°C. Stop reaction by adding 2.0 ml of 12 N H2SO4 into each tube and carefully mixing each tube thoroughly. Finally, measure the absorbance of each tube against the reagent blank at 540 nm.
Statistical Methods.
All the data were expressed as mean ± SEM. One-way analysis of variance (ANOVA) was used to analyze the data. Comparisons between the different diet groups were made at a two-sided alpha level of 0.05 adjusted by Tukey’s procedure for multiple comparisons. Statistical calculations were performed using Sigma Stat Version 2.03 (Systat Software, Inc, San Jose, CA).
Results
Cataract Incidence and Degree.
In order to initially verify that our dietary galactose treatment had induced cataract formation, we examined lenses of all experimental animals by slit lamp prior to necropsy. We then reinspected all lenses at the time of necropsy and recorded these findings as final results for the rate and grade of cataracts in each treatment group. Then the eye samples were prepared for routine histology and observed under the microscope. All rats in Groups 2 (galactose only) and 3 (galactose + genistein) were found to have cataracts, while no cataracts were found in the control rats, which was demonstrated by HE staining on lens section as shown in Figure 1A–C. Even though genistein treatment did not prevent cataract formation, the score and the extent of cataracts found in the genistein-treated rats were significantly decreased in comparison with the galactose-only animals. The cataract incidence and degree in all experimental groups are shown in Table 1.
Effects of Galactose and Genistein on Some Metabolic Markers and Hormone Levels.
To profile the extent to which the experimental diets might or might not have affected the metabolism of exposed animals, we measured some metabolic markers and hormone levels. Serum glucose concentrations were significantly lower in rats fed the galactose diet and treated with genistein (Group 3) compared with the other two groups. There were no differences in serum concentrations of insulin and leptin among three groups. The concentrations of serum somatostatin were significant lower in the galactose-only rats (Group 2) compared with control (Group 1) and galactose + genistein (Group 3) rats. Finally, the concentrations of serum testosterone in Group 3 (galactose + genistein) were significantly higher compared with the other two groups (Fig. 2).
Immunohistochemistry of Cx43.
The Cx43 expression on the lens was examined by immunohistochemical staining. Cx43 was located in the layer of epithelium cells in the lens (Fig. 1D–F). It is noteworthy that in comparison to the galactose-only (E) and control (D) animals, genistein (F) increased the immunohistochemical expression of Cx43 on the epithelial cells of lens.
Localization sGC Alpha and Beta Subunits in the Rat Eyes.
The immunohistochemical localization of sGC alpha and beta subunits in rat eyes showed that sGC alpha and beta subunits were colocalized in the outer plexiform layer, inner nuclear layer (INL), inner plexiform layer, and layer of ganglion cells (GCL), but not in the outer nuclear layer, external limiting membrane, inner segments of rods and cones, or the outer segments of rods and cones (Fig. 3). Nevertheless, there were no differences in expression of sGC alpha and beta subunits among three groups of experimental rats suggesting that this molecular mechanism is not likely to be involved in cataract formation or prevention, at least in this model.
Discussion
The results of our present study show that the isoflavone genistein is not able to prevent the induction of cataract induced by a galactose-rich diet, but genistein did show a beneficial effect by delaying the progression and reducing the extent of cataract formation. This novel finding suggests that some degree of protection against cataract might be afforded in humans by dietary intake of genistein or possibly other isoflavones in common foodstuffs such as soy and other leguminous food items.
Galactose is an energy-providing nutrient and also a necessary basic substrate for the biosynthesis of many macromolecules in the body. Problems with galactose metabolism can cause a variety of clinical manifestations in animals and humans (39). The feeding of a 20% galactose diet to weanling rats uniformly induces cataracts within 4 weeks in the present experiment. This might mean that even normal animals have a threshold to digest a certain amount of dietary galactose. Our previous study shows that long-term feeding of high lactose diets as a means to elicit potential toxicity of dietary galactose retards the growth rate in the female rats (40). Results of the present experiment indicate that dietary galactose decreases the serum concentration of somatostatin, suggesting a potential mechanism for this effect of dietary galactose on growth in both male and female rats. However, in contrast to some findings in our previous 7-month lactose experiment in the female rats (40, 41), the present study did not show any significant effects of galactose diet on serum concentrations of glucose, insulin, leptin, and testosterone. This finding suggests that these effects of galactose may be duration related and/or sex specific in rats.
Genistein is a phytoestrogen that occurs naturally in the diet. There is widespread interest in phytoestrogens as chemopreventive agents for a variety of diseases and cancers based on epidemiologic evidence (42). In recent years, phytoestrogens have attracted increased attention among the public and in the medical community because of accumulated evidence from a large body of literature suggesting that consumption of plant-based foods rich in these phytochemicals may benefit human health (43). Results of our present study are interesting because they clearly show that dietary genistein decreases serum glucose levels and increases serum testosterone levels in the adult male rats. These unexpected results imply that further research on metabolic effects of genistein is needed.
Beyond demonstrating a partial protective effect of genistein against cataract progression in this animal model, a noteworthy finding is that genistein increased the immunohistochemical expression of Cx43 in the lenses of rats on a galactose-rich diet. Cx43 is associated with osmotic regulation within the lens (23, 24). It is well recognized that a metabolite of galactose, galactitol, can accumulate in the lens, which may cause osmotic imbalance and cellular swelling, and in turn alter gap junction expression (39). Since the protein kinase inhibitor staurosporine prevents Cx43 phosphorylation and the loss of Cx43 from intercellular junctions (44), the genistein-related increase in Cx43 maybe due to local inhibition of protein kinase C (PKC) activity (45).
Cyclic GMP plays a central role in visual excitation at the level of photoreceptor cells, where the light-evoked decrease in cGMP causes a change in the membrane potential because this second messenger controls the conductance of cation influx through cGMP-gated channels (46–48). Such a regulatory effect of cGMP at the level of photoreceptor cells is known to be governed mainly by particulate guanylate cyclase (GC) (49, 50). Our immunohistochemical results are similar to previous reports (35, 51, 52). But we could not find any effects of genistein on expression of sGC, which suggests that the beneficial effect of genistein on cataract progression in this animal model does not involve the GC/cGMP pathway.
Given the results of this preliminary study showing a beneficial effect of genistein on progression of galactose-induced cataracts and a change in expression of the gap junction component Cx43 in the lens, additional animal studies are needed to more completely characterize these observations. For example, we speculate that if the galactose-rich diet were discontinued, rats being treated with genistein would show more prompt resolution of cataracts. A shorter period of loading galactose and earlier checks on the presence of cataracts might very well show that genistein delays the onset of cataracts by galactose. Cataract studies should be conducted in animals maintained on different diets, in order to compare the effects of isoflavone-rich versus isoflavone-poor foods. Ultimately, well-informed human studies could result.
Cataract Incidence and Score in Experimental Rats

Representative photographs of lens sections stained with hematoxylin and eosin (HE, panels A, B, and C). In panel A, arrow indicates the lens area is intact; in panels B and C, arrow indicates the cataract-damaged lens area. Cx43 expression on the epithelial cells on the lens are shown in panels D, E, and F by immunohistochemical staining. Positive immunostaining for Cx43 is indicated by a red reaction product. The arrow indicates the epithelial cells on lens. The rats were fed with control (A, D); galactose (B, E); and galactose + genistein (C, F) diets. Magnification −200. Color figure is available in the on-line version.
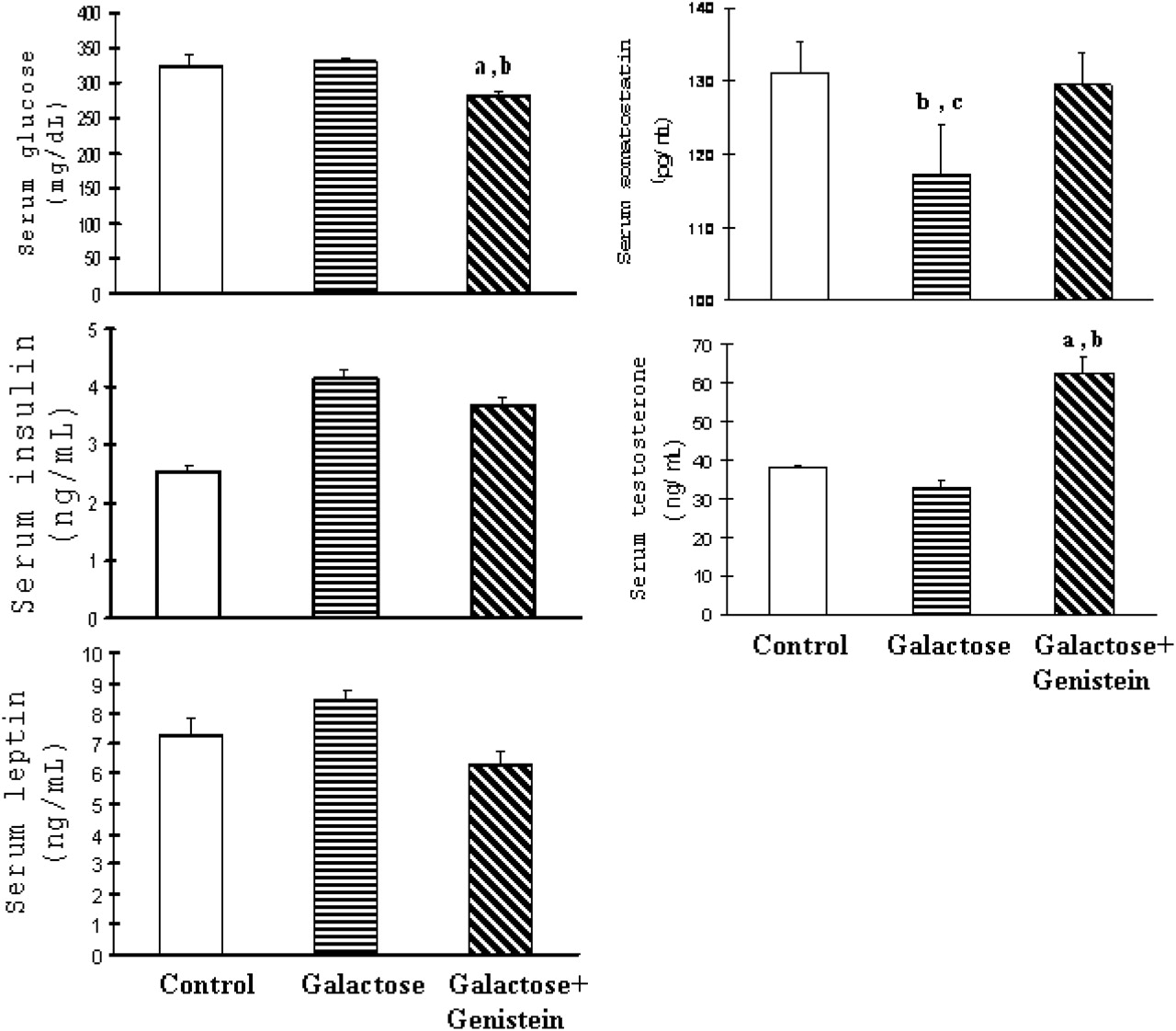
Serum concentrations of glucose (A), insulin (B), leptin (C), somatostatin (D), and testosterone (E) in rats fed with control, galactose, and galactose +genistein diets. Each value represents the mean ± SEM of six animals. In the figure, “a” denotes P < 0.05 when galactose + genistein group is compared with control group; “b” denotes P < 0.05 when galactose +genistein group is compared with galactose group; “c” denotes P < 0.05 when galactose group is compared with control group.
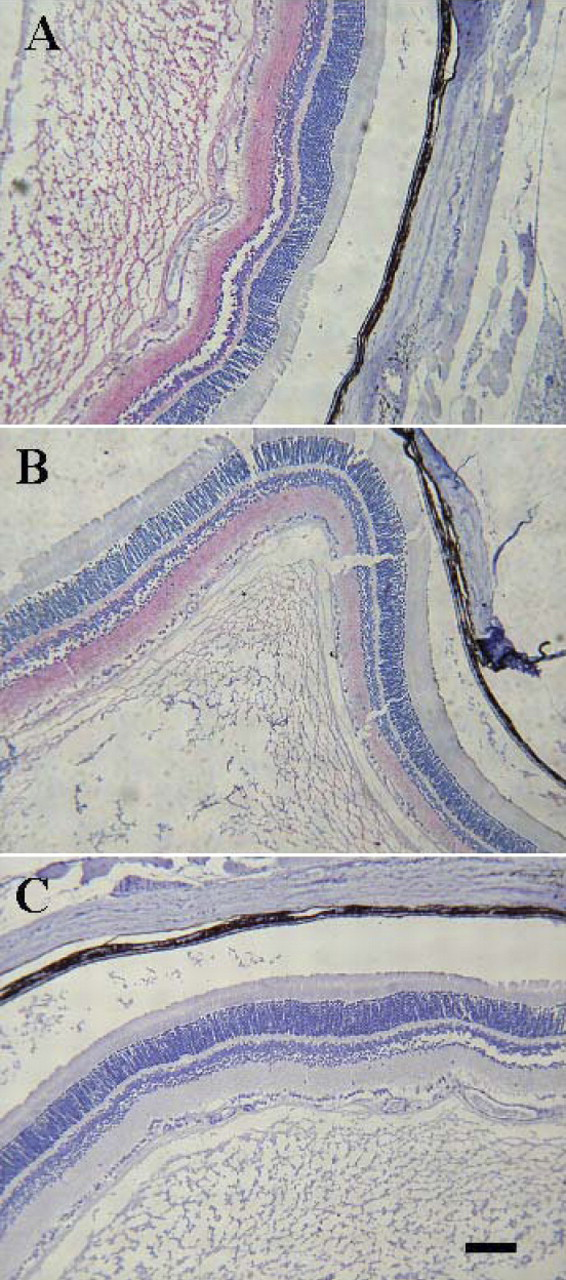
Immunohistochemical localization of sGC subunit proteins in the sections of rat eyes. Sections were incubated with primary antibody against sGC alpha (A), beta (B), or normal rabbit serum (NRS, C). Positive immunostaining for sGC alpha and beta is indicated by a red reaction product. Bar = 100 μm. Color figure is available in the on-line version.
Footnotes
This study was supported in part by NIH grant of USA (R03HD350830 to CLH) and a Grant-in-Aid for Introduction of Outstanding Scientists from Nanjing Agricultural University of China (G804002 to FS).