
Abstract
Systemic lupus erythematosus (SLE) is a complex systemic autoimmune disorder of unknown etiology. However, the definitive mechanisms remain obscure. Recently, transglutaminase 2 (TG2) was implicated in the pathogenesis of SLE. Cystamine, which inactivates TG2 activity by forming a mixed disulfide, may interfere with and inhibit other thiol-dependent enzymes such as caspases. To investigate the effects of cystamine in SLE pathogenesis, this in vivo study assessed the serum and macrophage response after administration of cystamine to NZB/W F1 mice. The experimental results demonstrated for the first time a significant reduction in TG2 and matrix metalloproteinase (MMP)-9 activity; tissue inhibitor of metalloproteinases (TIMP)-1, TIMP-2, TG2, tumor necrosis factor alpha, and tumor growth factor beta mRNA expression; and anticardiolipin autoantibodies (aCL) in NZB/W F1 mice following cystamine administration. It strongly suggests the therapeutic potential of cystamine in SLE.
Introduction
Systemic lupus erythematosus (SLE) is a complex systemic autoimmune disorder, affecting mainly females, of unknown etiology, with skin, joint, renal, and nervous system manifestations (1). SLE is characterized by dysregulation in multiple arms of the immune system and production of antinuclear antibodies (2). Multiple cytokines are involved in SLE pathogenesis, including upregulation of tumor necrosis factor (TNF)-α and interleukin (IL)-1, IL-4, IL-6, IL-10 and IL-18 (3, 4). However, the definitive mechanisms underlying this disease remain obscure. Although SLE pathogenesis is complex and remains poorly understood, consensus exists that impaired clearance functions of macrophages for dying cells are important factors to the accumulation of apoptotic cells in tissues of SLE patients (5). Recently, transglutaminase 2 (TG2) was implicated in the pathogenesis of SLE (6, 7). TG2 is a multifunctional enzyme with Ca2+-dependent protein cross-linking activity and guanosine triphosphate-dependent G protein function (8–10). Thus, TG2 has a variety of functions, including cell adhesion, cell spreading, angiogenesis, transmigration, and apoptosis (11). A recent study indicated that accumulated TG2 could cause apoptosis in human U937 monocytic cells and neuroblastoma cells (12–13). Additionally, another study demonstrated that TG2−/− mice develop lupus-prone symptoms, such as autoanti-bodies, immune complex glomerulonephritis, and impaired engulfment of apoptotic cells by macrophages (14). These experimental observations strongly suggest that TG2 may play important roles in SLE pathogenesis.
Recently, studies observed various physiological processes during inflammatory response in SLE patients, including elevated matrix metalloproteinases (MMPs; Refs. 15, 16). The MMPs are members of a family of zinc-dependent enzymes and are known to disrupt the cysteine-Zn2+ bond via a conformational alteration (17). In addition to their involvement in many normal biological processes, MMPs are also active in pathological processes including angiogenesis, inflammation, and autoimmunity (18, 19). MMP-9, as a member of the MMP family, degrades basement membranes, including type IV collagen. These processes help leukocytes entering and exiting the vascular system and lymph circulation (20). This study also indicated that MMP-9 could cause autoimmunity by cleaving myelin basic protein or type II gelatins to produce remnant epitopes. Furthermore, a recent study demonstrated significantly elevated MMP-9 activity in SLE patient sera (16). Therefore, elevated MMP-9 likely plays an important role in SLE pathogenesis.
Cystamine, which inactivates TG2 activity by forming a mixed disulfide (21–25), may interfere with and inhibit other thiol-dependent enzymes such as caspases (26). Recently, it has been demonstrated in a number of neurodegeneration models that cystamine plays important roles in neuroprotection (24, 27). Cystamine prolonged survival and decreased abnormal movements in a transgenic model of Huntington’s disease (27). To date, no report has investigated the effect of cystamine on TG2 in SLE pathogenesis. Because MMP-9 and TG2 play important roles in SLE pathogenesis, this study examines the effects of cystamine on MMP-9 and TG2 in a NZB/W F1 mice model. To investigate the beneficial effects of cystamine in SLE pathogenesis, this in vivo study assessed the serum and macrophage response after cystamine was administered to NZB/W F1 mice. These experimental results demonstrated for the first time a significant reduction in MMP-9 activity, TG2, TNF-α, and tumor growth factor beta (TGF-β) mRNA expression and anticardiolipin autoantibodies (aCL) in NZB/W F1 mice following cystamine administration.
Materials and Methods
Mice and Macrophage Isolation.
Female BALB/c and NZB/W F1 mice were obtained from the animal center, SINICA, Taiwan, and housed under supervision of the Institutional Animal Care and Use Committee at Chung Shan Medical University. To confirm the onset of disease in female NZB/W F1 mice, proteinuria was monitored biweekly with Albustix test strips (Bayer Diagnostics, Hong Kong) from the age of 14 weeks for 10 weeks and scored as trace (<0.3 mg/ml), 1+ (<0.3–0.9 mg/ml), 2+ (<1–2.9 mg/ ml), 3+ (<3–20 mg/ml), or 4+ (<20 mg/ml), according to the manufacturer’s scoring system. The score over 1+ is considered as disease positive. The 188-day-old mice were divided into four groups (12 mice/group) including BALB/c given phosphate-buffered saline (PBS), BALB/c given cystamine, NZB/W F1 given PBS, and NZB/W F1 given cystamine for experiments. For cystamine (Sigma, St. Louis, MO) treatment, BALB/c and NZB/W F1 mice were injected intraperitoneally (ip) with 100 μl of 10 mM PBS or 100 μl of 10 mM cystamine daily for 14 days (27). Heart blood of mice was obtained following CO2 sacrifice by ventricle puncture with a 1-ml syringe for serum collection. Peritoneal macrophages were collected as described elsewhere (28). The procedure is briefly described below. Mice were sacrificed by CO2 asphyxiation and immersed in 5% Dettol (chloroxylenol) solution (Dettol Co., Clevedon, UK). When the fur was dry, the mice were injected ip with 10 ml of ice-cold Iscove’s Modified Dulbecco’s Medium-fetal calf serum (IMDM-FCS) (Gibco BRL, Eggenstein, Germany) (5%, decomplemented). The anterior and lateral walls of the abdomen were gently massaged. The peritoneum washes were collected and filtered through a nylon filter into a 15-ml Falcon tube. Cells were then spun down by centrifuge at 1200 rpm for 5 mins and resuspended in IMDM-FCS (5%, decomplemented). All cells were transferred onto the petri dish and allowed to adhere for 2 hrs. Nonadherent cells were discarded.
TG Activity.
TG activity was determined by a modification as previously described (29). Microplates were coated with 20 mg/ml of N, N′-dimethylcasein and blocked with nonfat dry milk (0.5% in 0.1 M Tris-HCl, pH 8.5). Twenty micrograms of cell lysates was extracted and sonicated for 10 secs in 5 mM Tris-HCl, pH 7.4, 0.25 M sucrose, 0.2 mM MgSO4, 2 mM dithiothreitol (DTT), 0.4 mM phenylmethylsulfonyl fluoride, and 0.4% Triton X-100, and was then loaded onto the previously coated microplates. The lysates were incubated for 1 hr in 100 mM Tris, pH 8.5, 20 mM CaCl2, 40 mM DTT, and 2 mM 5-(biotinamido) pentylamine (BP) (Pierce Biotechnology Inc., Rockford, IL), and detection of the bounded BP was performed at OD of 405 as previously described (29). Purified transglutaminase (0.1 U) from guinea pig liver was used as positive control (Sigma).
Gel Zymography.
MMP-9 and MMP-2 activities were analyzed by gelatin zymography. Ten microliters of 10× diluted serum or 20 μg protein lysate of macrophages from BALB/c or NZB/W F1 mice was separated by an 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gel polymerized with 1 mg/ml gelatin. Gels were washed once for 30 mins in 2.5% Triton X-100 to remove the SDS and then soaked in the reaction buffer containing 50 mM Tris-HCl, 200 mM NaCl, 10 mM CaCl2, and 0.02% (w/v) Brij 35 (Sigma, St. Louis, MO; pH 7.5) for 30 mins. The reaction buffer was changed to a fresh one, and the gels were incubated at 37°C for 24 hrs. Gelatinolytic activity was visualized by staining the gels with 0.5% Coomassie brillant blue and quantified by densitometry (Appraise; Beckman-Coulter, Brea, CA).
RT-PCR.
All studies were carried out in a designated PCR-clean area. RNA was extracted by using Trizol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. Total RNA was isolated from macrophages of BALB/c and NZB/W F1 mice. RNA samples were resuspended in diethyl pyrocarbonate–treated water, quantified, and then stored at −80°C until use. RNA concentration and purity were determined by a spectrophotometer by calculating the ratio of optical density at wavelengths of 260 and 280 nm. The first-strand cDNA for RT-PCR was synthesized from total RNA (2 μg) using the Promega RT-PCR system (Promega, Madison, WI). The cDNAs encoding mouse TG2, tissue inhibitors of metalloproteinases type 1 to 4 (TIMP-1–TIMP-4), TNF-α, TGF-β, IL-10, and GAPDH were amplified by RT-PCR using the following primer pairs: 5′-AGGAC ATCAACCTGACCCTG-3′ (forward primer for TG2 cDNAs) and 5′-TTAGGCCGGG CCGATGATAAC-3′ (reverse primer for TG2 cDNAs), 5′-GCAGTGAAGAGTTTCTCATC-3′ (forward primer for TIMP-1 cDNAs) and 5′-TCATCGGGCCCCAAGG GATCT-3′ (reverse primer for TIMP-1 cDNAs), 5′-GCAGGAAAGGCAGAAGG AGAT-3′ (forward primer for TIMP-2 cDNAs) and 5′-TTACGGGTCCTCGATGTC AAG-3′ (reverse primer for TIMP-2 cDNAs), 5′-GCA-GATGAAGATGTACCGAGG-3′ (forward primer for TIMP-3 cDNAs) and 5′-TCAGGGATCTGTGGCGTT-GCT-3′ (reverse primer for TIMP-3 cDNAs), 5′-CTCTGT-GGTGTGAAGCTAGA-3′ (forward primer for TIMP-4 cDNAs) and 5′-CTAGGGCTGGATGATGTCAAC-3′ (reverse primer for TIMP-4 cDNAs), 5′-GCACAGAAAG-CATGATCCGC-3′ (forward primer for TNF-α cDNAs) and 5′-CTTGGTGGTTTGCTACGACG-3′ (reverse primer for TNF-α cDNAs), 5′-CCACTATTGCTTCAGCTCCAC-3′ (forward primer for TGF-β cDNAs) and 5′-TCAGCTG-CACTTGCAGGAGCGCAC-3′ (reverse primer for TGF-β cDNAs), 5′-GGCGCTGTCATCGATTTCT-3′ (forward primer for IL-10 cDNAs) and 5′-AGAAATCGATGACA-GCGCC-3′ (reverse primer for IL-10 cDNAs), and 5′-CCCACTAACATCAAATGGGG-3′ (forward primer for GAPDH cDNAs) and 5′-CCTTCCACAATGCAAAGTT-3′ (reverse primer for GAPDH cDNAs). The amplifications were performed in 50-μl reaction volumes containing 10× reaction buffer (Promega), 1.5 mM MgCl2, 200 mM deoxynucleotide triphosphates (dNTPs), 1 mM of each primer, and 2.5 units of Taq DNA polymerase (Promega) using a Perkin-Elmer Gene Amp PCR system 2400. Each cycle consists of denaturation at 95°C for 1 min, annealing at 55° for 45 secs, and amplification at 72°C for 45 secs. The RT-PCR–derived DNA fragments obtained by 25 PCR cycles were subjected to electrophoresis on a 1.7% agarose gel. The results were also scanned and quantified by densitometry (Appraise; Beckman-Coulter).
SDS-PAGE and Immunoblotting.
SDS-PAGE, using 12.5% acrylamide gel with 5% acrylamide stacking gel, was performed according to the method of Laemmli (30). Samples were reduced for 5 mins in boiling water with 0.0625 M Tris-HCl buffer, pH 6.8, containing 2.3% SDS, 5% 2-mercaptoethanol, and 10% glycerol. Samples applied to the gel were run at 100–150 V for 1.5 hrs and then electrophoretically transferred to nitrocellulose. The membrane was soaked in PBS with 5% nonfat dry milk for 30 mins at room temperature to saturate irrelevant protein-binding sites. Anti-TG2 and antiactin antibodies (Upstates, Charlottesville, VA) were diluted in PBS with 5% nonfat dry milk and incubated for 1.5 hrs at room temperature. The membranes were washed twice with PBS-Tween for 1 hr and secondary antibody conjugated with alkaline phosphatase was added. The substrate nitroblue tetrazolium/5-bromo-4-chloro-3 indolyl phosphate was used to detect antigen-antibody complexes. The blots were also scanned and quantified by densitometry (Appraise; Beckman-Coulter).
Anti–dsDNA Antibodies and aCL.
The anti-dsDNA antibodies and aCL were measured with commercial enzyme-linked immunoabsorbent assay kits (INOVA Diagnostic Inc., San Diego, CA). The assay is based on the binding of anti-dsDNA antibody or aCL from serum samples to purified dsDNA or cardiolipin antigen immobilized on microtiter wells. Goat anti-mouse IgG-HRP conjugate was used as the secondary antibody and the color developed with chromogenic substrate 3,3′,5,5′-tetramethylbenzidine. The enzymatic reaction (blue color) is directly proportional to the amount of anti-dsDNA antibodies and aCL present in the sample.
Statistical Analysis.
The paired t test was used to analyze for statistical significance. A P value of <0.05 was considered significant.
Results
Cystamine Downregulates TG2 Activity in Macrophages of NZB/W F1 Mice.
Because TG2 deficiency is associated with SLE, which is reportedly impaired in clearance of apoptotic bodies, this study assesses the effect of cystamine, a well-known TG2 inhibitor, on NZB/W F1 mice. Mice received peritoneal injections of 100 μl 10 mM cystamine daily for 14 days. No significant change in body weight was detected during the period of treatment (data not shown). To assess the effects of cystamine, a TG2 activity assay was performed (Fig. 1A). Protein extract was harvested from macrophages of BALB/c and NZB/W F1 mice treated with PBS or cystamine. Activity of TG2 was significantly decreased in macrophages of both BALB/c and NZB/W F1 mice treated with cystamine. The expression of TG2 protein was also examined in macrophages of BALB/c and NZB/W F1 mice treated with PBS or cystamine. Figure 1B presents the immunoblotting results for TG2. No variation in TG2 protein expression was detected for all mice groups (Fig. 1B).
Cystamine Downregulates MMP-9 Activity in NZB/W F1 Mouse Serum.
Recent studies have demonstrated that elevated MMP-9 activity is associated with various autoimmune diseases (31–33), including SLE (16). To investigate the effect of cystamine on MMP9 activity in SLE, NZB/W F1 mice were used as the experimental animal model. Sera, obtained from the mice (age 201 days) and treated with PBS or cystamine, were analyzed for MMP-9 and MMP-2 activity. Sera was sampled from four groups of mice for analysis: BALB/c mice given PBS; BALB/c mice given 100 μl 10 mM cystamine; NZB/W F1 mice given PBS; and, NZB/W F1 mice given 100 μl 10 mM cystamine. Figure 2A shows a representative gel zymogram. Activity of MMP-9 was reduced in sera of NZB/W F1 mice given cystamine, and not in the other three mice groups. Additionally, no significant difference in MMP-2 activity was observed among the four groups (Fig. 2A). To quantify MMP-9 and MMP-2 activity levels in the serum of BALB/c and NZB/W F1 mice treated with PBS or cystamine, densitometric analysis was utilized as described in the Materials and Methods Section. Figure 2B presents MMP-9 activity. Notably, MMP-9 activity was significantly decreased in sera of NZB/W F1 mice given cystamine (P < 0.01), and not in sera of BALB/c mice given cystamine. Conversely, differences in MMP-2 levels were not significant among all mice groups (Fig. 2C).
Cystamine Downregulates MMP-9 Activity and TIMP-1 and TIMP-2 mRNA Expression in Macrophages of NZB/W F1 Mice.
Because macrophages are crucial to SLE pathogenesis, the effects of MMP levels in macrophages from BALB/c or NZB/W F1 mice were also examined. Figure 3 presents the activity levels of MMP-2 and MMP-9 in BALB/c and NZB/W F1 mouse macrophages following treatment with PBS or cystamine with gel zymography analysis. Figure 3A shows a representative gel. A significantly decreased level of MMP-9 activity was observed in NZB/W F1 mice. To quantify MMP-9 and MMP-2 activity levels in macrophages of BALB/c or NZB/W F1 mice given PBS or cystamine, densitometric analysis was performed as described in the Materials and Methods section. Figure 3B indicates that the activity of MMP-9 was significantly decreased (P < 0.01) in macrophages from NZB/W F1 mice given cystamine, and not in BALB/c mice given cystamine. Additionally, a significant difference (P < 0.01) in levels of MMP-2 activity was observed between NZB/W F1 mice treated with PBS and NZB/W F1 mice treated with cystamine (Fig. 3C).
Because TIMPs, natural inhibitors of MMPs, are also expressed in macrophages and associated with the pathological processes of SLE (15), this study further assessed the expressions of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 mRNA by RT-PCR in macrophages of BALB/c and NZB/W F1 mice given PBS or cystamine. Figure 4A shows a representative RT-PCR gel. Expression of TIMP-1 and TIMP-2 mRNA was decreased. However, expression of TIMP-3 and TIMP-4 mRNA was not identified. Figures 4B and C present the quantified results for TIMP-1 and TIMP-2 mRNA expression in macrophages from BALB/c and NZB/W F1 mice given PBS or cystamine. A significant reduction in TIMP-1 mRNA expression was identified in macrophages from BALB/c (P < 0.01) and NZB/W F1 (P < 0.01) mice given cystamine. Furthermore, a significant decrease of TIMP-2 mRNA (P < 0.01) expression was only observed in macrophages from NZB/W F1 mice given cystamine, and not in macrophages from BALB/c mice given cystamine.
Cystamine Downregulates mRNA Expression of TNF-α and TGF-β in Macrophages of NZB/W F1 Mice.
Elevated expressions of various cytokines, including TNF-α, TGF-β, and IL-10, were implicated in the pathogenesis of SLE (3, 4, 34, 35). To further assess the affect of cystamine in NZB/W F1 mice on cytokine expressions, mRNA expression of TNF-α, TGF-β, and IL-10 was assessed. Figure 5A shows a representative result of RT-PCR analysis. Decreased expression of TNF-α and TGF-β mRNA was observed. However, expression of IL-10 mRNA was not detected. Figures 5B and C present quantified results for TNF-α and TGF-β mRNA expression in macrophages of BALB/c and NZB/W F1 mice given PBS or cystamine. Expression of TNF-α mRNA was significantly decreased in macrophages of NZB/W F1 mice given cystamine (P < 0.01), and not in macrophages from BALB/ c mice given cystamine. Additionally, expression of TGF-β mRNA was significantly reduced (P < 0.01) in macrophages of NZB/W F1 mice given cystamine, and not in macrophages of BALB/c mice given cystamine.
Cystamine Downregulates Levels of Anticardiolipin Autoantibody in Sera of NZB/W F1 Mice.
Anti-dsDNA antibody and aCL were examined in sera of BALB/c and NZB/W F1 mice (Table 1). No significant difference for anti-dsDNA autoantibody levels was detected in BALB/c or NZB/W F1 mice given cystamine, as compared with control mice given PBS. Additionally, no difference for aCL autoantibody level was detected in BALB/c mice given cystamine or PBS. However, the aCL autoantibody level was significantly reduced (P < 0.05) in NZB/W F1 mice given cystamine, as compared with control mice given PBS. Bovine serum albumin (BSA) antigen was used as a control.
Discussion
SLE is a complex disease, characterized by production of multiple antibodies, and it impacts various organs and systems. However, the precise mechanism of and treatment for SLE remain uncertain. This disease also has various clinical symptoms, including proteinuria, elevated MMP-9 activity, and increased TNF-α and TGF-β mRNA expression, that are associated with disease severity. This study demonstrates for the first time the beneficial effects of cystamine in SLE mice. Our results reveal that MMP-9 activity, and not MMP-2 activity, is significantly reduced in sera and macrophages of NZB/W F1 mice given cystamine, as compared with controls given PBS. Additionally, significant decreases of TIMP-1 and TIMP-2 mRNA levels, and not of TIMP-3 and TIMP-4 mRNA levels, were detected in macrophages of NZB/W F1 mice given cystamine. Moreover, significantly decreased TNF-α and TGF-β mRNA expression was demonstrated in macrophages of NZB/W F1 mice given cystamine, as compared with the controls given PBS. The level of aCL autoantibody was significantly reduced (P < 0.05) in NZB/W F1 mice given cystamine compared with that in control mice given PBS.
As a diverse-functional enzyme, TG2 was implicated in causing apoptosis in various cells, including human U937 monocytic cells and neuroblastoma cells (6, 12–13). Additionally, TG2 knockout mice developed lupus-prone symptoms, including autoantibodies, immune complex glomerulonephritis, and impaired engulfment of apoptotic cells (14). This study investigated the effects of cystamine on TG2 of macrophages in BALB/c and NZB/W F1 mice. As expected, TG2 activity is decreased by cystamine in macrophages of BALB/c and NZB/W F1 mice (Fig. 1A). Moreover, no effects of TG2 protein expression by cystamine were observed in cystamine-treated BALB/c and NZB/W F1 mice (Fig. 1B). These results are similar to results of a previous study in the YAC128 mouse model of Huntington’s disease. Decreased TG2 activity is observed in YAC128 mice and their wild-type controls given cystamine, and normal levels of TG2 protein expression are presented in all groups of mice (36). Because TG2 is associated with apoptosis and impaired engulfment of macrophages in SLE, our experimental results suggest that the downregulated TG2 activity by cystamine in SLE is beneficial. However, the involved actions of downregulated TG2 activity by cystamine in macrophages of NZB/W F1 mice are still unclear.
Because TG2 activity is required for membrane type 1–MMP-dependent activation of MMP-2 (37), this study analyzed MMP activities and TIMP mRNA expression in macrophages from cystamine-treated BALB/c and NZB/W F1 mice. In BALB/c mice, TIMP-1 mRNA expression (Fig. 4B) was reduced and no significant difference existed between MMP-9 and MMP-2 activity and TIMP-2 mRNA expression after administration of cystamine. In NZB/W F1 mice, wide-ranging results were obtained. MMP-9 activity was significantly reduced in both sera and macrophages, and MMP-2 activity was decreased significantly in macrophages of NZB/W F1 mice after treatment with cystamine (Figs. 2 and 3). Additionally, mRNA expression of TIMP-1 and TIMP-2 in macrophages of NZB/W F1 mice was decreased significantly (Fig. 4) after treatment with cystamine. These experimental results indicate that decreased MMP-9 and MMP-2 activity is independent of the variation of TIMPs in BALB/c and NZB/W F1 mice treated with cystamine, also suggesting that TG2 is involved in modulating MMP activity and TIMP mRNA expression. However, the relationship between modulation of MMPs and TIMPs in NZB/W F1 mice requires further identification.
Various cytokines are implicated in SLE pathogenesis. Notably, TNF-α is significantly elevated in SLE patients and is associated with increased apoptosis in SLE (7). TNF-α also plays a crucial role in SLE pathogenesis in mouse models (34) and in human disease (35). Cystamine reduces expression of TNF-α mRNA (Fig. 5), suggesting that cystamine has a therapeutic effect on SLE. Additionally, a recent study demonstrated that TG2 participates in activation of TGF-β, which is essential for in vivo induction of TG2 in both macrophages and apoptotic cells (6, 38). This study accounts for the reduction of TGF-β in macrophages of NZB/W F1 mice given cystamine.
Complicated and multifactorial interaction among various genetic and environmental factors is probably involved in SLE. T cells from patients with SLE have been known to be activated in vivo and have provided help to autoreactive B cells that led to various abnormal expressions, including signaling molecules and defective signal transduction pathways (39, 40). Lack of the 36-bp exon 7 [zeta mRNA/exon 7(−)] leads to downregulation of zeta protein in T cells from SLE patients and is responsible for the cytokines’ dysregulation (41). Additionally, abnormal inflammatory response to thioglycolate and impaired clearance of apoptotic cells by peritoneal macrophages in NZB/W F1 mice have been reported (42). The differential responses to thioglycolate between NZB/W F1 and BALB/c mice could be evidence to account for the selective macrophage responses in this study. Moreover, recent studies indicate that cystamine or its metabolites could act through a TGase-independent mechanism, including inhibition of caspase-3 activity (26), increasing cellular levels of the antioxidants glutathione and cysteine (26, 43), and heat-shock protein expression (27). These findings indicate that cystamine may cause differential effects on various signal molecules and signal transduction pathways that are defective solely in SLE.
Taken together, the diverse functions of cystamine and multiple disorders in SLE could contribute to selected phenomena, including reduced MMP-9, TG2, TNF-α, TGF-β, and aCL autoantibodies, in NZB/W F1 mice given cystamine. However, further study will be needed to clarify the precise mechanisms.
This study identified for the first time a significant reduction in MMP-9 activity and in TG2, TNF-α, and TGF-β mRNA expression in macrophages from NZB/W F1 mice administered cystamine. Additionally, a reduced level of aCL autoantibodies was detected in NZB/W F1 mice given cystamine as compared with control mice given PBS. Although the precise mechanism of cystamine action on SLE remains unclear, these experimental results may suggest the therapeutic potential of cystamine in treating SLE.
Autoantibodies in Sera of BALB/c and NZB/W F1 Mice a
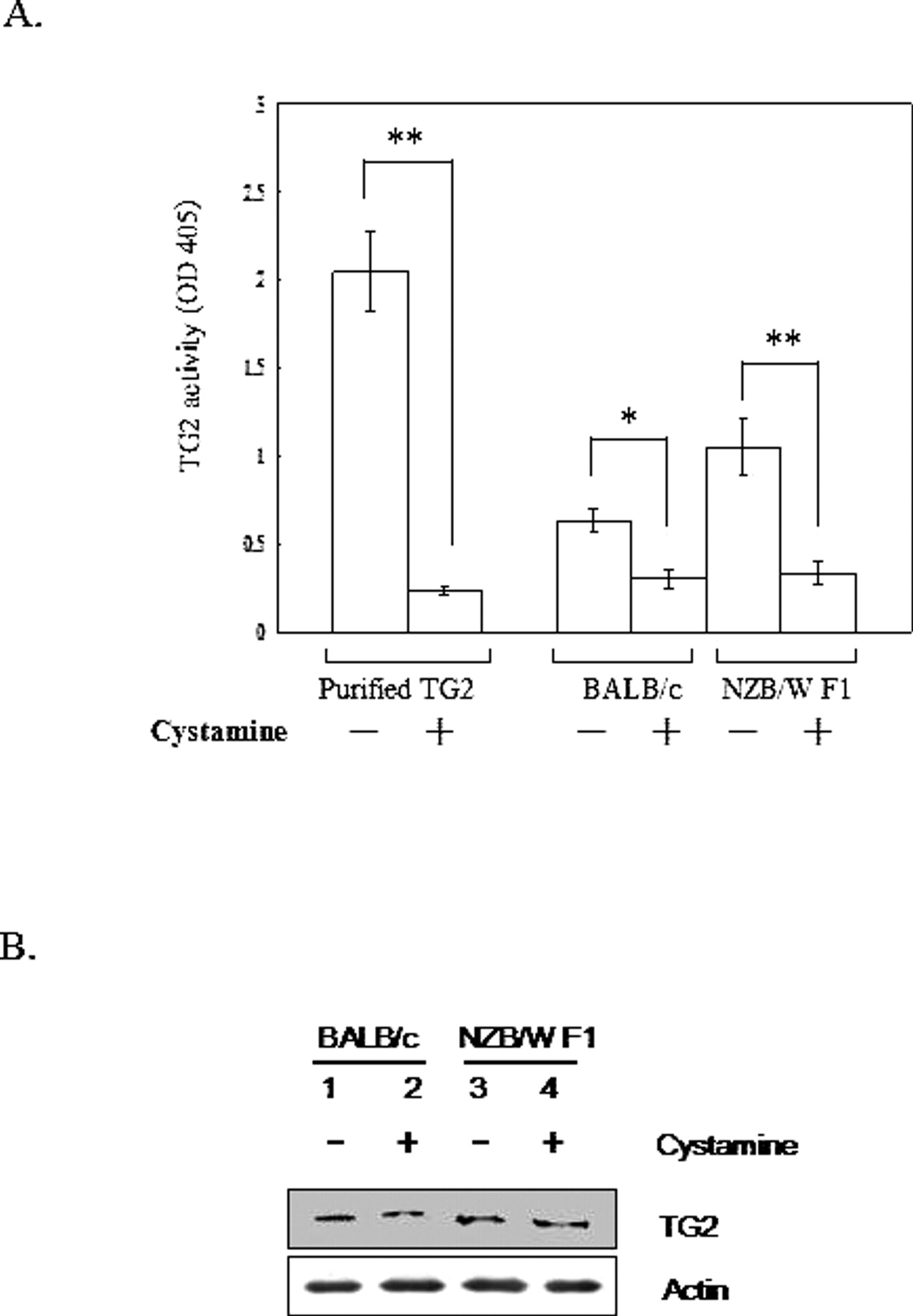
Protein lysate and total RNA of macrophages derived from BALB/c and NZB/W F1 mice treated with PBS or 100 μl 10 mM cystamine for 14 days were harvested. (A) Optical density of transglutaminase reactivity was measured at OD 405. Data are presented as mean ± SE. Three independent experiments were performed. **P < 0.01; *P < 0.05. (B) Immunoblot analyses were performed to detect TG2 expression. Actin was used as internal control. Three independent experiments were performed.
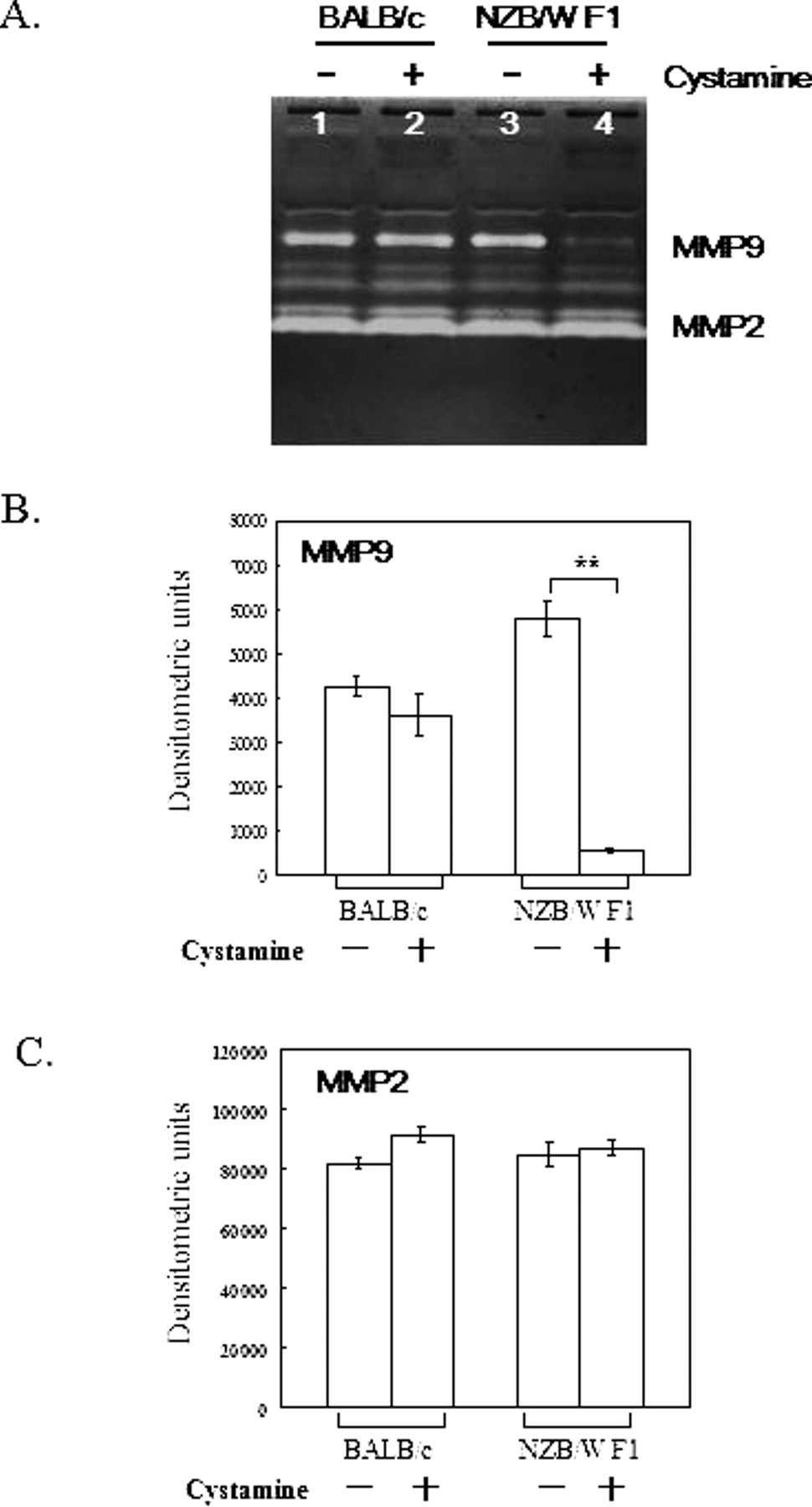
Activity of MMP-9 and MMP-2 in sera of BALB/c and NZB/W F1 mice. (A) Sera of BALB/c and NZB/W F1 mice treated with PBS or 100 μl 10 mM cystamine were analyzed for their MMP-9 and MMP-2 activity by gel zymography. The figure shows representative results for mice serum samples. (B and C) Densitometric analysis of MMP-9 and MMP-2, respectively. Results are from four separate experiments and are shown in bar graphs. **P < 0.01 compared with their counterparts.

Activity of MMP-9 and MMP-2 in macrophages of BALB/c and NZB/W F1 mice. (A) Macrophages of BALB/c and NZB/W F1 mice given PBS or 100 μl of 10 mM cystamine were evaluated for their MMP-9 and MMP-2 activity by gel zymography. The figure shows representative results from three similar experiments. (B and C) Densitometric analysis of MMP-9 and MMP-2, respectively. Results are from three separate experiments and are shown in bar graphs. **P < 0.01 compared with their counterparts.

Expression of TIMP mRNA in macrophages of BALB/c and NZB/W F1 mice. (A) Macrophages of BALB/c and NZB/W F1 mice treated with PBS or 100 μl 10 mM cystamine were assessed for their mRNA expressions of TIMP-1, TIMP-2, TIMP-3, and TIMP-4. The figure shows representative results of three similar experiments. (B and C) Densitometric analysis of TIMP-1 and TIMP-2, respectively. Results are from three separate experiments and are shown in bar graphs. **P < 0.01 compared with their counterparts.
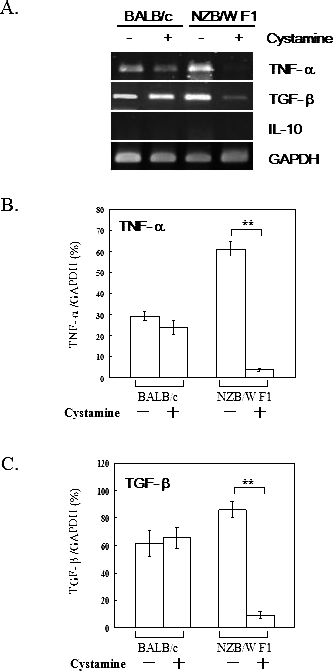
Expression of TNF-α, TGF-β, and IL-10 mRNA in macrophages of BALB/c and NZB/W F1 mice. (A) Macrophages of BALB/c and NZB/W F1 mice were treated with PBS or 100 μl 10 mM cystamine and analyzed for TNF-α, TGF-β, and IL-10 mRNA expressions. This figure shows representative results from three similar experiments. (B and C) Densitometric analysis of TNF-α and TGF-β, respectively. The results are from three independent experiments and are shown in bar graphs. **P < 0.01 compared with their counterparts.
Footnotes
This study was supported by grant NSC 94-2312-B-040-002 from the National Science Council, Taiwan.
Acknowledgements
We thank Dr. Mauro Piacentini, Department of Biology, University of Rome Tor Vergata, Rome, Italy, for his exquisite and crucial suggestions.