
Abstract
Antithrombin, the most potent anticoagulant in vivo, displays a significant conformational flexibility. The native five-stranded anticoagulant form transforms under different conditions or mutations to inactive six-stranded conformations: latent or polymer. However, the function, potential deleterious effects, and clearance of these forms are not completely known. The dimerization of latent antithrombin with a native molecule has been suggested to have thrombotic potential. We have assessed the potential thrombogenicity of high amounts of latent and polymeric antithrombin by experiments performed in mice and human plasma. Moreover, we have analyzed the clearance of 125I-labeled native, latent, polymer, and thrombin-complexed antithrombins in rat, as well as the clearance of latent antithrombin from plasma of patients treated with commercial concentrates. Our results show that high plasma levels of latent or polymeric antithrombin do not interfere with the anticoagulant function of native antithrombin. Moreover, we confirm that all monomeric forms of antithrombin have similar turnover. Finally, we show that polymers have the longest half-life of all conformers, being in circulation for prolonged periods of time. In conclusion, our data support that latent and polymeric antithrombin would not likely have a thrombotic effect, thus dispelling doubts about the potential harmful effect of latent antithrombin present in commercial concentrates for therapeutic use. Moreover, the suggested antiangiogenic role of latent antithrombin, together with its stability in plasma and its negligible thrombogenicity raises the possibility of its use as a new antiangiogenic drug.
Introduction
Antithrombin, a member of the serpin superfamily of proteins, is an essential anticoagulant regulator of blood coagulation (1). The function of antithrombin, like that of other serpins, can be attributed to its flexible structure, which allows it to display extraordinary conformational changes leading to the entrapment and inactivation of the protease (2). After docking with the proteinase, antithrombin undergoes a striking conformational transition that drives the insertion of the cleaved-reactive loop as an extra strand into β-sheet A (2). However, such structural flexibility also makes serpins vulnerable to even minor changes affecting their conformational stability (3). Antithrombin might transform to a relaxed six-stranded form by cleavage of the reactive loop by non–target proteinases (4). Further observations have raised the possibility that a similar A-sheet transition may occur in antithrombin even if the loop is intact (5). By homology with PAI-1, this six-stranded noncleaved structure was named latent antithrombin. In addition, sequential insertion of the reactive loop of one molecule into the β-sheet A of another molecule results in chains of polymers (6). Heating promotes transformation of antithrombin to the latent and polymeric forms. Antithrombin (and other serpins) rapidly polymerizes at temperatures higher than 60°C (7). Transition to the latent form is slow and requires lower temperatures. Thus, spontaneous conformational change of native antithrombin to its latent form can be achieved even at physiological temperatures (37°C) (7–9). These data explain that the pasteurization process is probably responsible for up to 40% of the latent antithrombin present in some commercial antithrombin concentrates (10). Severe reduction of the pH also promotes antithrombin polymerization (11). Finally, mutations affecting flexible regions of antithrombin and other serpins might favor the spontaneous or lower-temperature–induced transition to the latent or oligomeric forms (12).
Conformational changes leading to six-stranded forms have key functional relevance. The insertion of the reactive loop (either inside the molecule itself or into another molecule to give latent or polymeric forms, respectively) transforms native antithrombin to a noninhibitory form, losing its anticoagulant function (13). Thus, any factor able to induce transformation of native antithrombin to any inactive form would increase the risk of venous thrombosis (12, 13). In addition, such conformational changes might confer a disadvantage over and beyond that loss of activity. The intracellular aggregation of polymers might cause cell damage (14). Also, polymers of other serpins both in plasma and in tissues have been suggested to have direct pathological consequences (15, 16). Of even greater interest is the suggested thrombotic gain-of-function of latent antithrombin (13). The latent conformation is able to form heterodimers with native antithrombin, resulting in inactivation of the partner native molecule (17). For all these reasons, high concentrations of latent antithrombin have been suggested to be linked to an increased risk of developing venous thrombosis (13).
The objective of our study was to investigate the potential thrombogenicity of latent and polymeric antithrombins, and to evaluate the clearance of these molecules from circulation.
Materials and Methods
Materials.
Purified human antithrombin was obtained from commercial concentrates for therapeutic use (Kybernin-P, ZLB Behring, King of Prussia, PA). Purified human thrombin was acquired from Calbiochem (EMD Biosciences Inc., San Diego, CA).
Venous blood from five healthy blood donors was anticoagulated with 3.2% citrate. Fresh plasma from these samples was obtained by centrifugation and immediately stored in aliquots at −70°C. We also prepared a pool of citrated plasma obtained from 100 healthy individuals. Finally, we obtained plasma from a patient who had a type-I antithrombin deficiency caused by a R129X mutation in heterozygous state, and who was not undergoing anticoagulation therapy.
Preparation of Latent, Polymeric, and Thrombin-Complexed Antithrombin.
In order to avoid polymer formation, transformation of antithrombin to its latent conformation was slowly achieved by moderate heating (45°C for 5 weeks). The latent form used in our study was validated by means of its loss of anticoagulant activity and heparin affinity, its metastability on urea, and by mass spectrometry. In contrast, heating of purified antithrombin at 65°C for 10 mins led to its transformation into polymers. Thrombin-antithrombin complexes (TAT) were formed by mixing purified human antithrombin with thrombin (molar ratio 3 : 1, in order to completely entrap thrombin molecules) for 60 mins at 37°C.
Detection of Conformational Variants of Antithrombin.
Crossed immunoelectrophoresis of antithrombin in the presence of heparin was carried out as previously described (8).
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was performed in 10% (w/v) polyacrylamide gels, as previously described (17). Non-denaturing PAGE was essentially performed as indicated elsewhere (9). Addition of 6 M urea to the nondenaturing gel allowed differentiation of native and latent antithrombin, as previously reported (9).
Gels were stained using Coomassie blue when using purified proteins, or dried and exposed to autoradiographic films when using radiolabeled proteins. For plasma samples, 0.5–1 μl of plasma was run in nondenaturing gels, blotted to polyvinylidene difluoride (PVDF) membranes, and immunostained with goat anti-human antithrombin polyclonal antibody (Sigma Chemical Co., St. Louis, MO), followed by rabbit anti-goat IgG–horseradish peroxidase conjugate (Sigma), with detection via an ECL kit (Amersham Biosciences, Piscataway, NJ).
Measurement of Plasma Antithrombin Activity and Levels.
Antithrombin activity was determined by chromogenic methods. Anti-factor Xa (anti-FXa) assays were performed with heparin, bovine FXa, and S-2765 chromogenic substrate (Instrumentation Laboratory, Lexington, MA). Anti-IIa assays were performed in presence and absence of heparin, bovine thrombin, and chromogenic substrate (ethyl-malonyl-S-Pro-Arg-pNA.AcOH–CBS 61.50-; STA Antithrombin, Diagnostica Stago, Asnières sur Seine, France).
Antigen levels were measured both by enzyme-linked immunoabsorbent assay (ELISA) and Laurell rocket technique.
Hypercoagulability Marker.
Levels of F1+2 were quantified by means of a commercial ELISA system (Enzygnost F1+2, Behring), according to manufacturer’s instructions.
Thrombogenicity of Conformational Variants of Antithrombin.
Female C57 mice, 6 weeks of age, were caged in groups of five. They were intraperitoneally injected for 12 days with 50 mg/kg/day of either latent, polymeric antithrombin, or vehicle (phosphate-buffered saline, PBS). Citrated plasma was obtained 1 hr after the last injection of antithrombin and used to identify the presence of latent and polymeric antithrombin in circulation, as well as for assessing the antithrombin activity and levels.
In addition, we developed an in vitro model in which citrated human plasma from five healthy donors was mixed with latent or polymeric antithrombin to achieve concentrations of 600 μg/ml (four times the normal concentration of native antithrombin in plasma). As a control, identical volume of PBS was added to the plasma. Plasmas were incubated for 24 hrs at 37°C and evaluated for antithrombin activity and F1+2 levels. Finally, to further investigate the effect of latent antithrombin on plasma, citrated human plasma from five healthy donors was mixed with either PBS or latent antithrombin (600 μg/ml), and anti-IIa activity in presence and absence of heparin was determined. The prothrombin time of these samples was also evaluated. As a reference for a reduced anticoagulant function, we used plasma with a type-I antithrombin deficiency (caused by a R129X mutation in heterozygous state) supplemented with PBS. In all cases, experiments were performed in triplicate.
Protein Radiolabeling and Clearance Studies.
Native, latent, and polymeric antithrombin complexes as well as TAT complexes were radiolabeled with 125I using Iodogen, essentially as described elsewhere (18). Specific activities of the labeled proteins were as follows: native antithrombin 0.43 mCi/mg; latent antithrombin 0.41 mCi/mg; polymers of antithrombin 0.49 mCi/mg; and thrombin-antithrombin complexes 0.51 mCi/mg.
Sprague-Dawley rats (200 g) were anesthetized with ketamine (90 mg/kg)/xylazine (8 mg/kg) and injected intracardiacally over a period of approximately 10 secs with 100–200 μCi kg−1 of either native, latent, polymeric, or thrombin-complexed antithrombin. At selected time intervals (1, 10, 30, and 60 min, 20 and 40 hrs), blood (200 μl) was collected from the heart into 20 μl of 0.5 M EDTA. Blood samples were centrifuged, and plasma was aliquoted and immediately frozen (−70°C). Twenty-five μl of plasma was counted for its 125I content in a γ-counter (LKB–Wallac Pharmacia, Turku, Finland) in triplicate. The initial time point, taken 1 min after injection, was considered to represent 100% radioactivity in the circulation.
All experimental procedures were performed in accordance with the University of Murcia approved institutional animal care guidelines.
Patients with Antithrombin Deficiency Treated with Antithrombin Concentrates.
Four patients with antithrombin deficiency (2 congenital and 2 acquired by severe sepsis) were injected intravenously with a bolus of 20 U/kg of Kybernin-P. Citrated plasma samples were obtained before treatment, and at 10 min, and 6, 24, 48, and 72 hrs after the injection. Patients and family members were fully informed of the aim of this study, which was performed according to the declaration of Helsinki as amended in Edinburgh 2000.
Statistical Analysis.
Data are presented as mean ± SD. Statistical comparisons were made by paired two-tailed t test using Stat View software (Abacus Concepts, Berkeley, CA).
Results
Preparation of Latent and Polymeric Antithrombin.
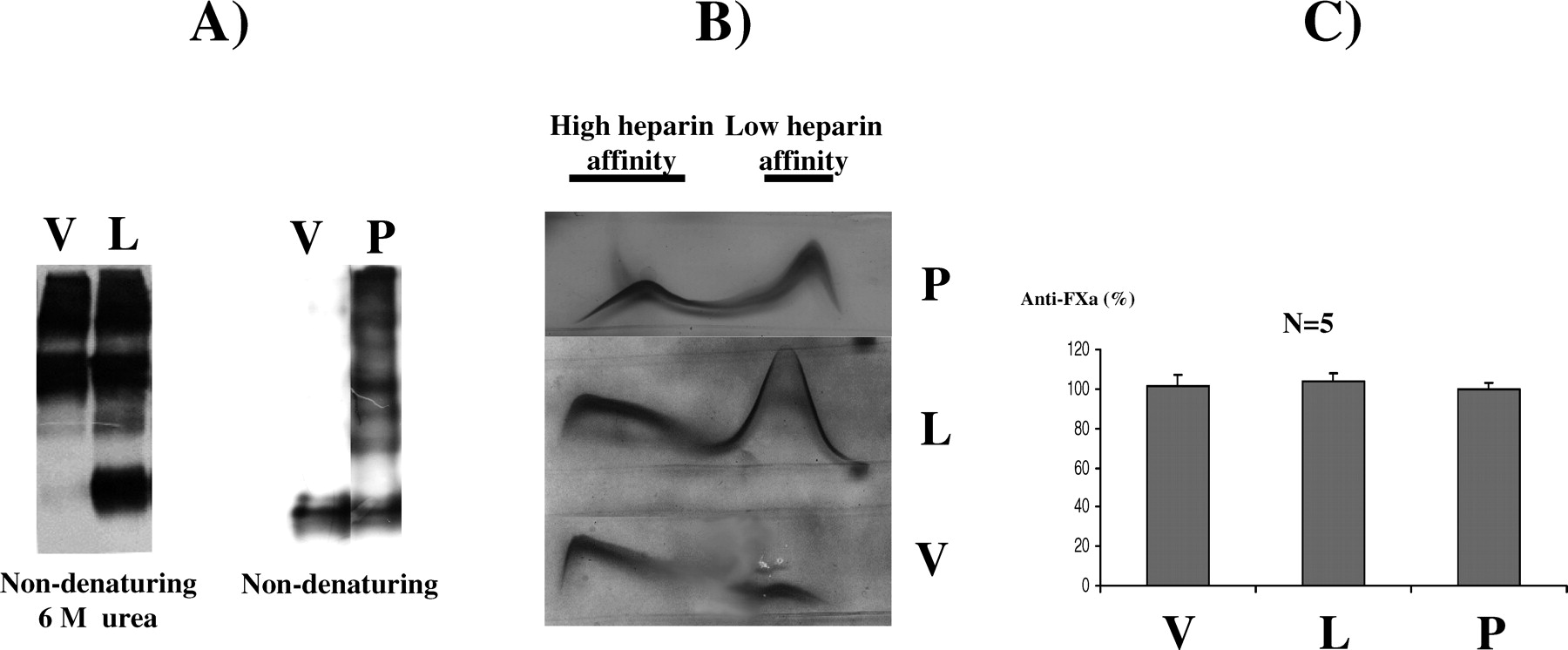
The commercial concentrate of purified human antithrombin used in our experiments (Kybernin-P) contains up to 5% of latent antithrombin. The native antithrombin of this concentrate displayed biochemical and functional properties identical to that of native antithrombin purified from plasma (data not shown). Incubation of this concentrate at 45°C for 4 weeks fully transformed antithrombin to the latent form, as demonstrated by nondenaturing PAGE in presence of 6 M urea (Fig. 1A). As previously shown (8), the latent form had low heparin affinity (Fig. 1B) and displayed no anti-FXa activity (Fig. 1C). As expected, mass spectrometry revealed an identical mass for native and latent antithrombin (57863.6 Da and 57866.9 Da, respectively). This antithrombin was biochemically and functionally identical to latent antithrombin purified from plasma (data not shown). In contrast, incubation of the commercial concentrate at 60°C for 15 min induced polymerization of the native antithrombin (Fig. 1A). Polymers also displayed low heparin affinity (Fig. 1B) and had no anti-FXa activity (Fig. 1C).
Native, latent, and polymeric antithrombin displayed identical electrophoretic profile on SDS-PAGE under reducing or nonreducing conditions (Fig. 1D), as reported elsewhere (8). In contrast, the TAT complex showed a slower electrophoretic mobility (high molecular weight bands in Fig. 1D).
Iodine labeling of these forms modified neither the electrophoretic (on SDS or nondenaturing PAGE) nor the functional features of the antithrombin conformers (data not shown).
Potential Thrombogenicity of Latent and Polymeric Antithrombin.
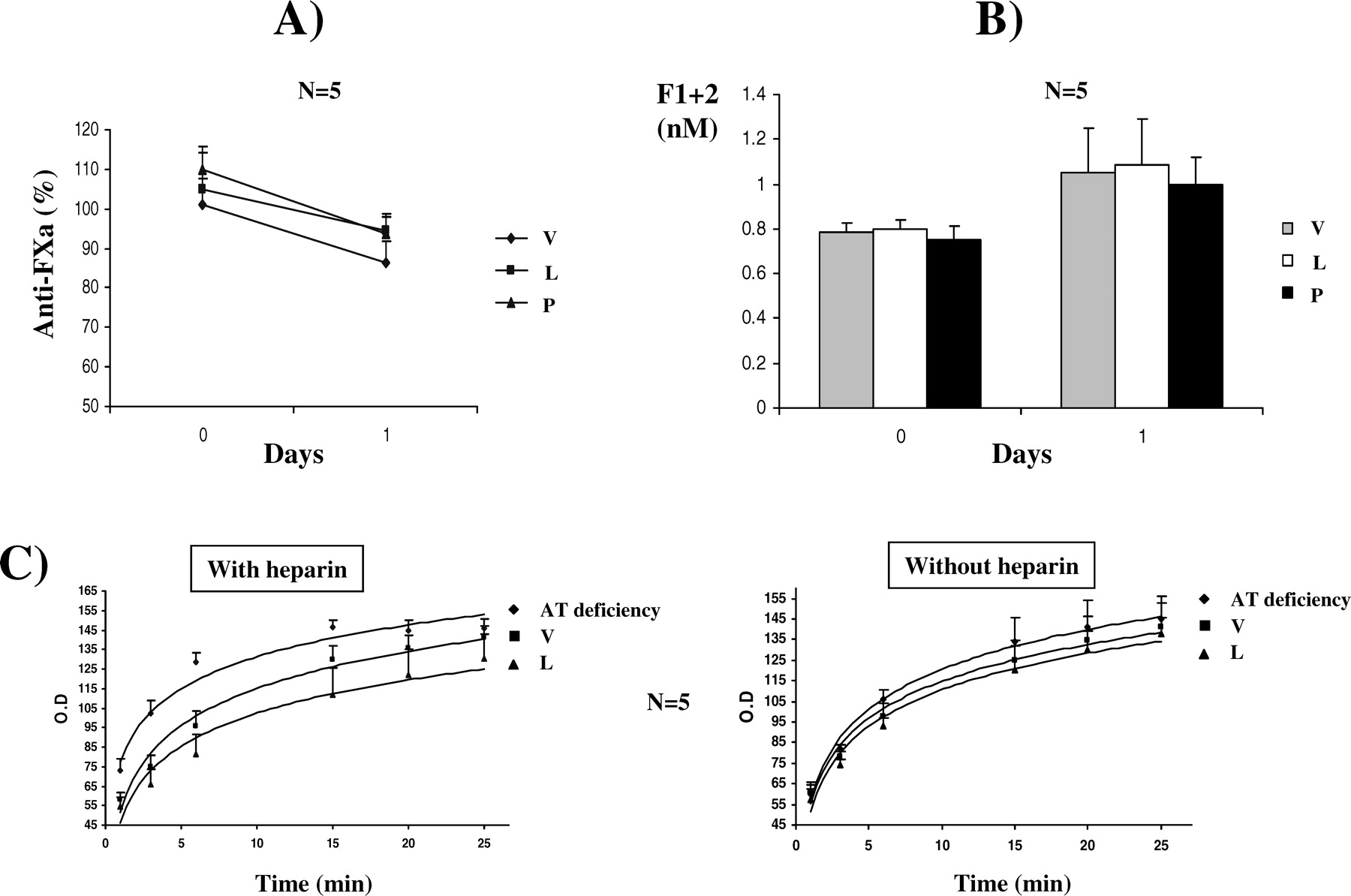
Mice injected with 50 mg/kg/day of latent and polymeric antithrombin for 12 days had significant amounts of these forms in blood compared with control animals. One hour after the last injection, half of the circulating antithrombin was latent or polymeric (Figs. 2A and B). However, these animals displayed no thrombotic signs for the length of treatment. Postmortem analyses revealed no signs of thrombosis or intravascular coagulation, although we did not assess thrombosis in small vessels (data not shown). In agreement with this result, plasma anti-FXa activity in mice treated with latent and polymer conformers was similar to that observed in controls (Fig. 2C).
It could be argued that the conformational variants of human origin might have minor effect in mice hemostasis, despite the high homology of antithrombin in both species (19). To clarify this issue, we incubated citrated human plasma with both forms of antithrombin at 600 μg/ml for 24 hrs at 37°C. The presence of high amounts of latent or polymeric antithrombin (four times the concentration of native antithrombin) did not significantly modify the anticoagulant activity of the sample (Fig. 3A). The minor loss of anti-FXa activity observed after 24 hrs might be explained by the transformation to the latent form of a few molecules of native antithrombin induced by the incubation at 37°C, as reported previously (8, 9). In accordance with this finding, only a slight increase in the F1+2 levels was observed in these samples (Fig. 3B), although these differences did not reach statistical significance. We also used plasma with other anticoagulants (heparin, EDTA, and hirudin), and in all cases, addition of latent antithrombin did not shift the position of the major band from that of α-antithrombin to the slightly slower mobility characteristic of the dimer. Moreover, in all these conditions, the anti-FXa was not affected (data not shown). In addition, the presence of 600 μg/ml of latent antithrombin (four times the normal concentration of native antithrombin in plasma) did not impair the anti-IIa activity of plasma, both in presence and absence of heparin (Fig. 3C). Finally, we determined the prothrombin time of these samples to evaluate the effect of 600 μg/ml of latent antithrombin on the coagulation reaction after stimulation with thromboplastin and calcium. The presence of latent antithrombin (up to four times the normal concentration of native antithrombin in plasma) did not significantly modify the prothrombin time (20.5 ± 3.1 secs without latent antithrombin and 18.6 ± 2.2 secs with latent antithrombin; P = 0.429).
Clearance of Different Forms of [125I]-Antithrombin in Rats.
An assessment of the in vivo clearance of latent, native, polymeric, and thrombin-complexed antithrombins was carried out in rats. The 125I-labeled forms of antithrombin were injected into rats, and blood samples were collected at various time intervals from 1 min to 2 days for determination of their elimination. All four conformers had a biphasic clearance pattern, with a rapid phase of initial clearance. Data of radioactivity disappearances were fitted to the double exponential equation in all cases, as described previously (20), using Ultra Fit software (Biosoft, Great Shelford, UK). The results from 5 different experiments with each conformer supported that latent antithrombin was removed from the circulation at a similar rate as the native protein, with a half-life of 71 ± 16 mins and 73 ± 19 mins, respectively (P = 0.795). As expected, TAT complexes were more rapidly removed from the circulation, with a half-life of 26 ± 7 mins (P = 0.005). Of interest, polymers had the slowest clearance rates, with a half-life of 269 ± 110 mins (P = 0.015 vs. native antithrombin). The elimination curve within the first 60 min showing the kinetic of clearance for the different conformers is shown in Figure 4.
Nondenaturing electrophoresis (with and without urea) of these samples and further autoradiography demonstrated the presence of all antithrombin forms in circulation after injection and were consistent with the radioactive counts, supporting that the differential clearance might depend on the quaternary structure of antithrombin (Fig. 5).
Clearance of Latent Antithrombin in Patients.
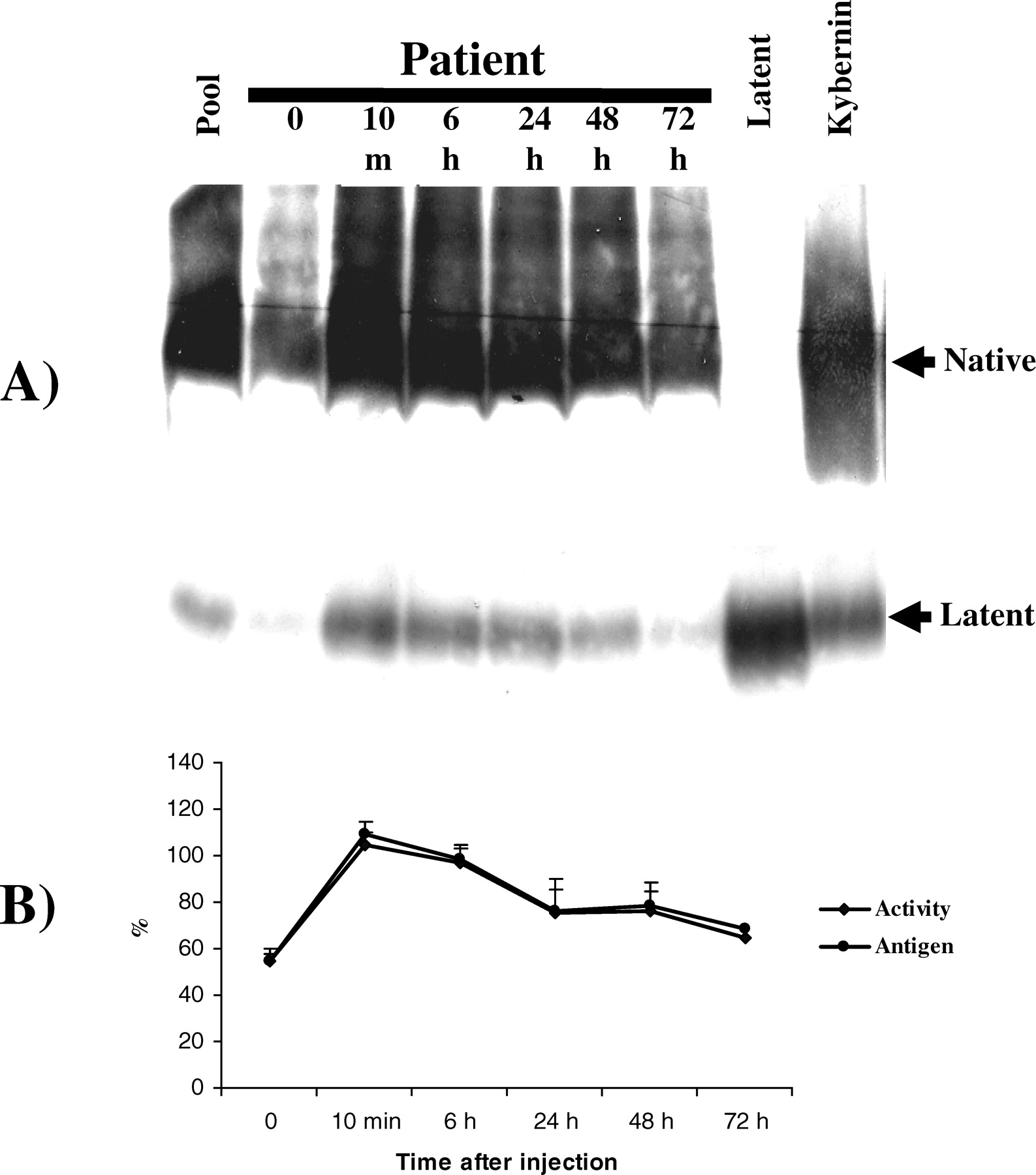
Because of concerns that foreign antithrombin could be different than homologous antithrombin, we evaluated the clearance of human latent antithrombin in humans. Injection of Kybernin-P to four patients with antithrombin deficiency resulted in a significant increase of both native and latent antithrombin because of the presence of both forms in the commercial concentrate. Of interest, in all cases, native and latent antithrombin displayed similar clearance rates (Fig. 6A). Moreover, we observed a similar rate of disappearance from plasma of functional antithrombin and antithrombin antigen (Fig. 6B).
Discussion
Antithrombin is the most important anticoagulant serpin in blood, as determined by the predisposition to thrombosis of subjects with heterozygous antithrombin deficiency, and by the embryonic lethality of homozygous deficiency (21). Recent reports have demonstrated a significant conformational flexibility of antithrombin and have supported the key functional consequences that some conditions or mutations have in inducing the transformation of native antithrombin to inactive latent or polymeric antithrombin (12, 13). These conformations might have additional deleterious effects, which may increase the risk of developing venous thrombosis, particularly latent antithrombin (13). There is no doubt that latent antithrombin loses its anticoagulant function. Thus, the transition of native antithrombin to its latent conformation will necessarily increase the risk of thrombosis. However, the aim of this study was to determine if the latent antithrombin could have clinical relevance per se; in other words, whether transition to the latent conformation might also result in a gain-of-function as has been suggested (13). Certainly, in 1994, two independent groups crystallized antithrombin, finding an unexpected result: the crystal was a heterodimer instead of the expected monomer (22, 23). There is consensus agreement that this dimer is formed by the linkage of a molecule of active (native) antithrombin to a second molecule of antithrombin in the inactive latent conformation (which was spontaneously converted from native during the crystallization procedure). Based on this observation, Zhou et al. suggested that the presence of latent antithrombin could be potentially deleterious because its linkage to form the heterodimer resulted in inactivation of the otherwise normal molecule linked to the latent antithrombin (17). The conversion of active antithrombin to the latent form is a slow process, which in health is likely only contributing to the natural senescence and turnover of the protein in the plasma. However, the increase of such form, which could be caused by different stress conditions or by treatment with commercial concentrates of antithrombin (with up to 40% of the antithrombin in the latent form), may be a contributory factor to an increased risk of thrombosis. In fact, some suppliers of concentrates of antithrombin have modified their procedures (with an additional heparin-affinity purification after the pasteurization process in some cases) to reduce or even eliminate any amount of latent antithrombin. Therefore, it is relevant to clarify whether these abnormal forms are indeed deleterious instead of merely ineffective. However, to our knowledge, the harmful effects of nonnative forms of antithrombin have been suggested by in vitro studies using purified antithrombins but have never been demonstrated in vivo or in plasma.
This study supports that high concentrations of both human latent or polymeric antithrombin have negligible effects on the anticoagulant activity (anti-FXa and anti-IIa) of plasma, both in vivo for mice and in vitro for humans, and therefore they are unlikely to have any thrombotic effects. One limitation of this in vivo model is that the generation of thrombin was not tested under different procoagulant stimuli or thrombotic situations. However, the results, which show that the injection of high amounts of polymers or latent antithrombin to mice does not modify the anticoagulant function of plasma antithrombin, argue against a potential thrombotic gain-of-function of polymeric and particularly latent antithrombin (13, 17). How could we explain such a discrepancy? One possibility is that these conformers might have a rapid metabolism. Certainly TAT complexes are rapidly cleared from circulation like other serpin-enzyme complex (24). If this clearance was dependent on the six-stranded conformation, latent, cleaved, and polymeric antithrombins (all with six-stranded conformation like the antithrombin complexed with thrombin) could be rapidly removed from circulation. This hypothesis could also explain the reduced levels of latent and cleaved antithrombin and the absence of polymers in plasma from healthy individuals (8, 9). However, the results in mice (Fig. 2) argue against this possibility, as significant amounts of latent and polymeric antithrombin remained in plasma 1 hr after injection. To gain further insights into this issue, we analyzed the clearance of radiolabeled conformers of antithrombin in a rat model. The results of this study are not definitive, being limited by the species difference and by the broad time intervals used for blood collection, but the findings here are consistent with previous data. Thus, TAT complexes are rapidly cleared from circulation, a result that is comparable with previous reports (9, 20, 25). More relevant, our results confirm that all monomeric antithrombin forms (native, latent, and cleaved) have similar half-lives, independent of their conformations (9, 25, 26). This finding in rats is further supported by the similar turnover of latent and native antithrombin seen in patients. By contrast, we show for the first time that polymeric antithrombin is retained longer in circulation than monomeric forms. Our findings may be relevant for understanding the clearance of similar conformers of other serpins, given the highly conserved structure of this superfamily (19). Moreover, preliminary analysis of the organs involved in the clearance of the antithrombin conformers supports a similar behavior for both monomeric forms, which differs from that of TAT. Thus, TAT complexes were cleared largely by the liver, but both monomeric forms (native and latent) were retained mainly within the spleen (data not shown).
All these results suggest that the rapid metabolism of these conformers could not explain the negligible role of polymers and particularly of latent antithrombin on the anticoagulant activity of the native form in plasma. More likely, the suggested deleterious effects of latent antithrombin, caused by its dimerization with a native molecule, would have minor influence in the control of anticoagulation if the formation of such dimers is reversible, as has been suggested (17, 27).
In summary, this study confirms that all monomeric antithrombins have similar clearance rates, while polymers of antithrombin remain within the circulation for prolonged periods of time. This study has only evaluated the effect of latent and polymeric antithrombin on the anti-FXa and anti-IIa activities of plasmatic antithrombin, supporting that both forms, even if present at high concentrations, seem to be inert. Our results do not confirm the suggested deleterious effects of latent antithrombin, and thus dispel doubts about their putative harmful effects in commercial concentrates and the suggested gain-of-function when generated in vivo. By contrast, the stability of latent antithrombin in blood and the absence of significant thrombotic effects may indirectly support its safe therapeutic use as a potent anti-angiogenic drug (28). It is unlikely that there are any other side-effects of latent and polymeric antithrombin, but further investigation is needed.
Conformational, biochemical, and functional features of purified antithrombin from a commercial concentrate transformed by heat to latent (45°C for 5 weeks) and polymeric antithrombin (65°C for 10 mins). (A) Nondenaturing PAGE (8%) ± 6 M urea. (B) Crossed immunoelectrophoresis in the presence of heparin. (C) Anti-FXa activity (percentage with respect to the activity observed in a pool of citrated plasma obtained from 100 healthy individuals). (D) SDS-PAGE under reducing (R) or nonreducing (NR) conditions. In all cases, the radiolabeled preparation was diluted in PBS to achieve the plasma concentration of antithrombin (150 μg/ml) and assayed for activity or electrophoretic mobility. Nat, native; Lat, latent; Pol, polymer. Mice injected for 12 days with 50 mg/kg/day of latent (L), polymeric (P) antithrombin, or vehicle (V). (A) Nondenaturing PAGE with or without urea. (B) Crossed immunoelectrophoresis. (C) Anti-FXa activity of citrate plasma. Values are presented as percentages of the activity displayed by a pool of five untreated mice (100%). Anticoagulant function of plasma antithrombin supplemented with 600 μg/ml of other antithrombin conformers. (A) Anti-FXa and (B) F1+2 levels observed in plasma from healthy individuals supplemented in vitro with 600 μg/ml of latent (L) or polymeric (P) antithrombin or PBS (vehicle V) and incubated at 37°C for 1 day. Mean values ± SD are given. (C) Kinetics of the chromogenic reaction of thrombin in presence and absence of heparin of plasma from healthy subjects supplemented with (L) or without (V) latent antithrombin. As a reference for a reduced anticoagulant function, plasma with a type-I antithrombin deficiency supplemented with PBS was used. Clearance of different radiolabeled antithrombin conformers in mice. Representative autoradiography of plasma samples showing the clearance of [125I]antithrombin forms from rat plasma. Samples were run in nondenaturing PAGE (with or without 6 M urea). Clearance of latent antithrombin in patients. (A) Representative Western blot of a nondenaturing PAGE in presence of 6 M urea of plasma samples from a patient with congenital antithrombin deficiency supplemented with 20 U/kg of antithrombin from a commercial concentrate (Kybernin-P). (B) Evolution of antithrombin levels and activity in patients supplemented with a commercial concentrate. Values are the percentage of the amount of antithrombin present in a pool of citrated plasma obtained from 100 healthy individuals.


Footnotes
This study was supported by Fundación Séneca (00583/PI/04), SAF2003–00840 (MCYT & FEDER), and SAF2004–07535 (MEC & FEDER). JC and JR contributed equally to this work.
Acknowledgements
The authors would like to thank Dr. Carrell for latent and native antithrombin purified from plasma.