
Abstract
Neuropeptide pituitary adenylate cyclase-activating polypeptide (PACAP) is a pleiotropic hormone that is involved in numerous physiologic functions. The present study examines the presence and the functional significance of PACAP and its receptor in the brain and astrocytes of tilapia (Oreochromis mossambicus). This is the first demonstration of the full-length nucleotide sequence of tPACAP gene in tilapia pituitary, brain, and cultured astrocytes. Two cDNA variants of the growth hormone–releasing hormone (GHRH)–PACAP gene were identified in tilapia pituitary, brain, and cultured astrocytes as a result of exon skipping with a long form (271 bp) encoding both tPACAP38 and tGHRH and a short form (166 bp) encoding only tPACAP38. The short form was found to be more abundant in astrocytes. Addition of ovine PACAP38 (1 nM) to cultured astrocytes significantly stimulated the expression of tPACAP38 at 4 hrs, but the effect dropped after 8 hrs of treatment. By contrast, the expression of PACAP type I receptor (PAC1-R) mRNA in the astrocytes was not responsive to PACAP38 treatment. The tPACAP38 expression also was activated by the cAMP analog, dibutyryl-cAMP, in a dose-dependent manner. Adding high salinity (170 mM NaCl, 500 mOsm/kg osmolarity) to cultured medium substantially increased astroglial tPACAP38 expression over 4 hrs to a level that was maintained for 16 hrs. This observation was not found when mannitol (270 mM) was supplemented as an osmolarity-enhancing agent (500 mOsm/ kg). Taken together, tPACAP expression in tilapia astrocytes was well regulated by exogenous PACAP, cAMP, and salinity and might be involved in the adaptation to high salinity when the fish is in a seawater environment.
Introduction
Pituitary adenylate cyclase-activating polypeptide (PA-CAP) is a neuropeptide that was originally purified from ovine hypothalamus tissues because of its capacity to stimulate cAMP production in cultured rat anterior pituitary cells (1). Based on its high amino acid sequence homology with vasoactive intestinal peptide (VIP), this peptide is one of the glucagons/secretin/growth hormone–releasing hormone (GHRH) superfamily. PACAP is present in two biologically active forms with 38 and 27 amino acid residues, designated PACAP38 and PACAP27, respectively (1, 2). GHRH is encoded in the same gene with PACAP in protochordates (3), fish (4–8), amphibians (9, 10), and birds (11, 12). At some point before or during the emergence of mammals, separate genes encoded PACAP and GHRH. The biologic functions of PACAP in mammals have been widely studied (14). The diverse functions of PACAP include the regulation of cell proliferation, differentiation, and apoptosis and the regulation of metabolism and the cardiovascular, endocrine, and immune systems. However, the physiologic events that coordinate PACAP responses have not yet been identified (15). PACAP is widely dispersed in the central nervous system (CNS) and may act as a hypophysiotropic factor in adenohypophysial hormone release and a neurotransmitter and neuromodulator in the CNS (13). Three PACAP receptors (PAC1-R, VPAC1-R, and VPAC2-R) have been identified. All of them are G protein–coupled receptors with a classical structure of seven transmembrane domains (16). PACAP type I receptor (PAC1-R) has a high binding affinity for PACAP38 and PACAP27 but a much lower affinity toward VIP, whereas PACAP type II receptors (VPAC1-R and VPAC2-R) have similar binding affinities toward PACAP27, PACAP38, and VIP (20–22). All three receptors signal by stimulating adenylate cyclase activity, increasing levels of intracellular cAMP (20–22). Astrocytes, the major macroglial cells in the CNS, are crucially involved in maintaining homeostasis. PACAP stimulates the proliferation of astrocytes at a physiologic concentration of 10−12 M (23). The presence of specific PACAP binding sites on rat astrocytes has been demonstrated (24, 25). Pharmacologic studies reveal that astrocytes predominantly express the PAC1-R that mediates cAMP accumulation by PACAP 103 times more effectively than VIP (17, 23, 24). PACAP also stimulates intracellular calcium mobilization in astrocytes, indicating that astrocytes might be the main target cells for PACAP in the CNS (23, 24, 26, 27).
The actions of PACAP in fish are more interesting, especially when PACAP serves as a hypophysiotropic agent (8). It effectively stimulates the secretion of GH (8, 28–32) and gonadotropin (33); it functions in the gut by affecting contractions of the intestines (34, 35); it can trigger catecholamine secretion from chromaffin cells (36); and it controls the differentiation and proliferation of cells during zebrafish ontogeny (37). PACAP was presented recently as a potential ovarian factor for mediating the activity of gonadotropin in the manner of paracrine/autocrine in zebrafish follicle cells (38). Mossambic tilapia, a euryhaline teleost, can adapt to various salinities of sea water (SW). Studies have shown that the physiologic responses involve the coexistence of two isoforms of creatine kinase in gills (39). Studies have demonstrated that creatine kinase provides energy in gills (40, 41) and the brain (42) during the transfer of fresh water (FW) to 25 parts per thousand (ppt) SW in tilapia. However, the underlying mechanism of tilapia brain in response to osmolarity challenge has not yet been examined. Astrocytes, the dominant component of glial cells in brain, are known to support a normal CNS by optimizing ion and pH homeostasis. Previous studies have demonstrated that cultured astrocytes under various osmotic conditions exhibit various biochemical and physiologic responses, including the augmentation of mRNA and protein expression of water channel (43), osmolytes, synthetic enzymes, and transporter expression (44). Astrocytes respond to hyperosmotic stress by activation of p38 kinase (45) and change in the regulated volume through the chloride channel (46). By contrast, astrocytes trigger a calcium response (47) in response to a hypo-osmotic challenge. Our preliminary study has found that the mRNA level of tPACAP38 in tilapia brain was significantly stimulated at 8 hrs after a transfer from FW to SW. However, the presence and involvement of PACAP and its receptor in tilapia brain, especially in specific types of cells, such as astrocytes, that are under osmotic stress, remains unknown. Therefore, the present study examined the effects of PACAP in cultured tilapia astrocytes. We isolated a new GHRH-PACAP gene from tilapia brain (designated tGHRH-tPACAP38, where t denotes tilapia). The tGHRH-tPACAP38 cDNA expressed in tilapia pituitary and astrocytes also were found. The role of tPACAP38 in astrocytes was investigated by considering (i) the expression of tPACAP38 and PAC1-R in tilapia astrocytes, (ii) the regulation of tPACAP38 expression by exogenous ovine PACAP38 and the signaling mechanism involved, and (iii) the effect of hyperosmolarity (NaCl and mannitol) on the expression of tPACAP38 and PAC1-R.
Materials and Methods
Chemicals.
Hydrocortisone and dibutyryl cyclic AMP (db-cAMP) were purchased from Sigma (St. Louis, MO), and ovine PACAP38 was purchased from Calbiochem (La Jolla, CA). NaCl and mannitol were purchased from Sigma. First, db-cAMP and ovine PACAP38 were dissolved in water and then diluted with L-15 medium to the desired concentrations before use. NaCl (170 mM) and mannitol (270 mM) were added directly to L-15 medium (300–310 mOsm/kg osmolarity) with 10% fetal bovine serum (FBS) to make hyperosmolarity at 500 mOsm/kg (according to previous studies, the osmolarity of cerebrospinal fluid is 450–500 mOsm/kg at 4–8 hrs following exposure to 25 ppt seawater) before use. Rabbit anti-human (ovine, rat) PACAP38 polyclonal antibody (dilution 1:500) was purchased from Phoenix Pharmaceuticals (Belmont, CA), rabbit anti-human PAC1-R polyclonal antibody (dilution 1:500) was purchased from LifeSpan Biosciences (Seattle, WA), and rabbit anti-human glial fibrillary acidic protein (GFAP) polyclonal antibody (dilution 1:1000) was purchased from Chemicon (Temecula, CA).
Animals.
Tilapia (Oreochromis mossambicus) fish were obtained originally from the Tainan Fish Culture Station of the Taiwan Fisheries Research Institute. All fish were maintained in a freshwater recirculating tank at 25°C–28°C on a 12:12-hr light:dark photoperiod and fed daily at the Institute of Biotechnology, National Dong Hwa University (Hualien, Taiwan). Euryhaline tilapia are able to live in both sea water (SW) and fresh water (FW).
Primary Culture of Astrocytes.
Brains of adult tilapia were used for primary astrocyte culture. Tilapia 40–60 g body wt were put on ice, immersed briefly (30 secs) in 70% alcohol (sterile), and decapitated. Brains were dissected out into 35-mm culture dishes containing ice-cold phosphate-buffered saline (PBS) with 10 mM glucose and were rinsed twice with PBS. They were digested with 0.5% (v/v) trypsin in L-15 medium (Gibco, Gaithersburg, MD) for 15 mins at 28°C, followed by mechanical dispersion with 25-G needles in L-15 medium supplemented with 10% FBS and filtration with a 40-μM Falcon cell strainer (Becton Dickinson Labware, Franklin Lakes, NJ) to remove cell debris. After centrifugation at 500 g for 10 mins, the supernatant was discarded, and approximately 2 × 106 cells were seeded in 25-T flask at 28°C without CO2 supplement. The medium was changed twice per week. Experiments were conducted after third-passage cultures, which showed greater than 95% positive staining for GFAP. These cultures were not immunoreactive to other major CNS cell markers, including microglial markers (GSA-IB4 and ED-1), neuronal markers (MAP-2 and NF-L), and oligodendrocyte (MBP and GalC).
Total RNA Isolation.
Male or female tilapia (80–100 g body wt) were put on ice and decapitated. The brains and pituitaries were removed and immediately homogenized for RNA isolation. Total RNA was isolated from the brains, pituitaries, or cultured astrocytes using RNA-Bee RNA isolation reagent (Tel-Test, Friendswood, TX) according to the manufacturer’s instructions.
Cloning of PACAP and PAC1-R Fragments from Tilapia Brain and Astrocytes.
The sequences of gene-specific primers in PACAP and PAC1-R were designed from EST clone of our tilapia brain cDNA library and VIP receptor of Takifugu rubripes (accession no. AJ408877), respectively. The cDNA was synthesized using a reverse transcriptase–polymerase chain reaction (RT-PCR) kit (Invitrogen Co., Carlsbad, CA). Five micrograms of total RNA from the brain was dissolved in 9 μl diethyl pyrocarbonate (DEPC) water with 1 μl oligo (dT)12–18 (0.5 μg/μl), and 1 μl dinucleoside triphosphate (dNTP) mix (10 mM) was denatured at 65°C for 5 mins and then chilled on ice immediately for 1 min. The denatured RNA was mixed with 4 μl 5× first-strand buffer, 2 μl of 0.1 M 1,4-dithiothreithol (DTT), and 1 μl RNAse Out (40 U) and denatured at 42°C for 2 mins. Subsequently, 1 μl random hexamer (50 ng/μl) and 1 μl Superscript II transcriptase (200 U) were added and incubated at 25°C for 2 min. The condition was set at one cycle at 25°C for 10 mins, one cycle at 42°C for 90 mins, and 70°C for 15 mins. After reverse transcription (RT), the product was incubated with 1 μl (2 U) Escherichia coli RNAse H at 37°C for 20 mins and stored at −20°C until analysis. The PCR products were purified with Gel/PCR DNA Fragments Extraction Kit (Geneaid Co., Bade City, Taiwan) and cloned into the pGEM-T Easy vector (Promega, Madison, WI) for sequencing. The same gene-specific primers of PACAP (EtPA-CAP38; Table 1) also were used to investigate the characteristics of exon skipping on the brain, pituitary, and astrocytes, whereas the gene-specific primers of PAC1- R (Table 1) were newly designed according to the data after sequencing in this study.
5′ RACE and 3′ RACE.
The gene-specific primers of EtPACAP38 (Table 1) were used for 5′ and 3′ rapid amplification of cDNA ends (RACE) to amplify the 5′ and 3′ cDNA ends using the SMART-RACE cDNA amplification kit (Clontech, Palo Alto, CA) with the same protocol as previously described (39). The amplified cDNA ends were cloned into the pGEM-T Easy vector for sequencing to complete the full length of tilapia GHRH/PACAP gene.
Validation of Semiquantitative RT-PCR Assays for tPACAP38 and β-Actin.
The gene-specific primers for tPACAP38 and β-actin are listed in Table 1. One microliter of cDNA solution (1:1 diluted with sterile water from stock) and two sets of primers for tPACAP38 and β-actin were used for PCR amplification simultaneously in 50 μl of PCR reaction samples. The parameters of the PCR reaction were 35 cycles at 95°C for 30 secs, 55°C for 30 secs, and 72°C for 1 min, and then a single cycle at 72°C for 7 mins. PCR products were separated in 1.2% agarose gel. After ethidium bromide (EtBr, 0.5 μg/mL) staining and photographing, the signals were analyzed by Phoretix ID standard software (PHORETIX, 61397/28052 MemoHASP1; Feng Jin Biomedical & Instruments Co., Ltd,, Taipei, Taiwan). Data were expressed as the mRNA ratio of PACAP or PAC1 receptor to β-actin following the measurement of the optical density for each cDNA fragment detected. The ratio to control presented the comparisons between control and treatments after β-actin (as an internal control) normalization. For validation of semiquantitative RT-PCR assays, the amplified cycle numbers of PCR were determined by checking the linear intensity of amplified product in various cycles, and the ratio of two set of primers for tPACAP38 and β-actin used in one reaction was measured the same way. In the meantime, inserted tPACAP38 plasmid as a template was used to generate a standard curve for the semiquantitative of tPACAP38 to do comparisons.
Southern Hybridization.
The PCR-amplified DNA (50 ng) was separated in a 1.2% agarose gel and transferred to a nylon membrane (NEN Life Science Products, Boston, MA), then analyzed by Southern hybridization as described previously (75). Gel-purified inserts of cDNA clones of tGHRH-tPACAP38 were used as probes (DIG Southern Starter Kit; Roche, Mannheim, Germany). The PCR product of RNA without RT was concurrently performed as a negative control.
Immunocytochemistry.
The experiment was performed using the VECTASTAIN ABC system kit (avidin and biotinylated horseradish peroxidase macromolecular complex; Vector, Burlingame, CA). Astrocytes grown on 12-well tissue culture plates that had flat bottoms and low evaporation lids (Falcon, Bedford, MA) were washed three times with 10 mM PBS for 3 mins and fixed in ice-cold methanol for 5 mins before being washed three more times with PBS for 3 mins. The fixed astrocytes were blocked at nonspecific binding sites with 1%–2% normal mouse serum (blocking serum) in PBS for 1 hr at 37°C. Astrocytes were identified with antibodies against GFAP (1:1000). The tPACAP38 and PAC1-R were identified using antibodies against PACAP38 (1:500) and PAC1-R (1:500), respectively. Astrocytes were incubated with primary antibodies or rabbit serum (as a negative control) at 4°C overnight and washed three times with PBS, followed by incubation with biotinylated secondary antibody at 37°C for 1 hr. After washing, the astrocytes were visualized by the addition of VECTASTAIN ABC. The cells were examined by phase-contrast microscopy.
Western Blotting.
Brain or cell homogenate (total protein 40 μg) was mixed with 6× electrophoresis sample buffer containing DTT (Sigma). The proteins were separated by electrophoresis on a 4%–12% gradient polyacrylamide slab gel and electrophoretically transferred to a polyvinylidene fluoride membrane (NEN Life Science Products). The blots were incubated for 2 hrs in blocking buffer (3% bovine serum albumin in TBST buffer) and were washed twice with TBST buffer (0.8% NaCl, 0.02% KCl, 0.3% Tris base, pH 7.5, 0.024% KH2PO4, and 0.2% Tween-20). Two membranes were incubated overnight at 4°C with rabbit anti-human PACAP38 (1:500) and rabbit anti-human PAC1- R (1:500) and internal control β-actin antibody (1:4000; Sigma), respectively. After incubation with secondary antibody (alkaline phosphatase–conjugated goat anti-rabbit antibody, 1:4000), the protein bands were developed by BCIP/NBT staining. Bands were quantified using computer software (PHORETIX, 61397/28052 MemoHASP1).
Statistical Analysis.
The experimental results were presented as mean ± standard error (SE). The significant difference (P < 0.05) between treatment and control was compared by Duncan multiple range test after one-way analysis of variance (ANOVA).
Results
Identifying PACAP (tPACAP38) Expressed in the Tilapia Astrocyte.
The full-length cDNAs that encode tGHRH and tPACAP38 in the brain of tilapia were constructed from overlapping 5′ and 3′ RACE. The cDNA was 798 bp, encoding 175 amino acids (Fig. 1A). The result demonstrates that the nucleotide sequence of tGHRH starts at 333, ends at 467, and encodes 45 amino acids, and tPACAP38 starts at 474, ends at 587, and encodes 38 amino acids. In this study, the exon skipping of GHRH/PACAP is evident in tilapia brain and pituitary, including astrocytes (Fig. 1B). This result was verified by Southern blot. The structural scheme elucidates the difference between the long and short forms and the characteristics of exon skipping (Fig. 1C). Based on the cloning and sequencing of various-size PCR products, the nucleotide sequence reveals that the long form (271 bp) comprises both tGHRH and tPACAP38. However, the short form (166 bp) consists of tPACAP38. Comparing these two forms indicates that the small product has a complete region of coding of tPACAP38 and a region that encodes 13 amino acids of tGHRH at the C terminus, but the region that encodes most of tGHRH mRNA, from nucleotide 323 to nucleotide 427 (105 bp), was absent. Additionally, viewing from the deduced amino acid sequence revealed that the long form, KRFERHAEEEL-DRALREILGQLTARHYLHSLMTIRAGDD, was substituted by the short form, KSDD, and 35 amino acids were absent (Fig. 1C). To investigate the underlying mechanism of PACAP regulation, the short form was designed to explore the alterations of tPACAP38 following various treatments in subsequent experiments.
Exogenous Ovine PACAP38 and db-cAMP Regulation of tPACAP38 in Cultured Astrocytes.
Whether exogenous PACAP regulates the expression of endogenous PACAP (tPACAP38) in the astrocytes of tilapia brain was investigated. Experiments were conducted using semi-quantitative RT to elucidate whether tPACAP38 is expressed in tilapia astrocytes in response to exogenous ovine PACAP38 PCR. The RT-PCR assays were validated carefully by determining the number of amplifying cycles and from the ratio of the two sets of primers for tPACAP38 and β-actin used in a single reaction. Meanwhile, tPACAP38 plasmid was inserted as a template to generate a standard curve to ensure the semiquantitative accuracy of tPACAP38 (data not shown). The cultured astrocytes were treated with exogenous ovine PACAP38 (1 nM) for various periods. The data showed that the expression of tPACAP38 by treated astrocytes increased significantly for 2 hrs, reaching a peak at 4 hrs (Fig. 2A). The stimulation of tPACAP38 expression in astrocytes by exogenous ovine PACAP38 depended on concentration (Fig. 2B). However, the effect of exogenous ovine PACAP38 at 5 nM on tPACAP38 expression was significant, reaching a peak at 2 hrs and then declining to 4 hrs (data not shown). Cyclic AMP is well known to be the main second messenger that is involved in exogenous PACAP stimulation through the protein kinase A (PKA) signaling pathway (27), so experiments were designed to determine whether intracellular cAMP level affects the expression of tPACAP38. Treatment of cultured astrocytes with 1.5 mM db-cAMP, a cAMP analog, was associated with strong stimulation of tPACAP38 for 2 hrs, after which the response declined (Fig. 3A). Stimulation with db-cAMP for 2 hrs significantly promoted the expression of tPACAP38 in a concentration-dependent manner (Fig. 3B).
Expression of PAC1-R in the Tilapia Astrocyte.
Primary cultures were used to explore whether the PACAP receptor was expressed in the tilapia astrocyte and thus to determine whether PACAP is a potential paracrine/ autocrine factor in astrocytes. First, T. rubripes PAC1-R gene (accession no. AJ408877) primers were designed for use in RT-PCR of tilapia brain, and a 510-bp partial sequence of PAC1-R from tilapia brain was cloned (data not shown). Subsequently, based on the tilapia PAC1-R sequence, gene-specific primers of tilapia PAC1-R (446 bp) and one more nested primer of tilapia PAC1-R (267 bp) were designed to analyze and verify the existence of PAC1- R in various tissues of tilapia. The partial nucleotide sequence of PAC1-R (446 bp) was obtained (Fig. 4A). PAC1-R was present in tilapia brain, pituitary, and astrocytes, as determined using RT-PCR with tilapia PAC1-R primer (Fig. 4B). Adding exogenous ovine PA- CAP38 to the cultured tilapia astrocyte did not affect the expression level of PAC1-R mRNA (Fig. 4C). The downstream activation of PACAP may involve the cAMP-PAK signaling pathway to promote astrocytic PACAP expression.
Colocalization of tPACAP38 and PAC1-R in Astrocyte.
In the present work, cultured astrocytes were subcultured with at least three passages and were used to perform experiments. The light field view and anti-GFAP immunostaining of the cultures demonstrated that more than 95% of the cells were positive stained for astroglial marker GFAP (Fig. 5A). They were not immunoreactive to microglial markers (ED1 or GSA-IB4) or oligodendrocyte markers (MBP and GalC). Particularly, they were virtually devoid of neurons (not immunoreactive to MAP-2 or neurofilament L; data not shown), which is consistent with the results of Franke et al. (48). The anti-PACAP38 and anti-PAC1-R antibodies were used to test the protein levels of tPACAP38 and PAC1-R in tilapia astrocytes (Fig. 5B and C). Based on our earlier findings, astrocytes also were separately treated with exogenous ovine PACAP38 and db-cAMP for 4 hrs and 2 hrs, respectively, before immunocy-tochemistry. Astrocytes stained with anti-PACAP38 antibody had a denser signal in the cytoplasm than the control group, indicating an increase in the expression of tPACAP38 peptide. These results were confirmed by Western blot analysis (Fig. 5C). However, the control did not differ from the treated astrocytes following anti–PAC1-R staining, revealing that treatment with exogenous ovine PACAP38 did not affect the expression of PAC1-R, which is consistent with the findings of RNA expression experiments.
Expression of tPACAP38 Under Hyperosmotic Stress.
The cultured astrocytes were treated with NaCl-and mannitol-hyperosmolarity medium for various periods. The data indicated that the PACAP expression of NaCl-treated astrocytes increased significantly for 4 hrs to a level sustained for up to 16 hrs (Fig. 6A). Mannitol treatment did not significantly change the expression of PACAP in tilapia astrocytes. Both NaCl and mannitol treatments did not alter the expression of PAC1-R in tilapia astrocytes, even after 16 hrs (Fig. 6B). The anti-PACAP38 and anti–PAC1-R antibodies were used to determine the protein levels of tPACAP38 and PAC1-R in tilapia astrocytes, respectively (Fig. 7A and B). The astrocytes also were separately treated with NaCl and mannitol for 4 hrs before the immunocytochemical experiment was performed. A comparison with control group revealed that astrocytes stained with anti-PACAP38 antibody had a denser signal in the cytoplasm, suggesting an increase in the expression of tPACAP38 peptide following NaCl treatment but not mannitol treatment. This result was confirmed by Western blot analysis. However, the absence of any difference between the control and treated astrocytes after anti–PAC1-R antibody staining demonstrated that the treatments of NaCl and mannitol did not influence the expression of PAC1-R, which is consistent with the results of the mRNA expression experiments.
Discussion
VIP and PACAP have been shown as neuroprotective agents in vivo and exert several actions on neurons, glia, and inflammatory cells in vitro (49). Numerous studies have shown that PAC1-R is present in rat astrocytes (17, 25, 50). Previous immunohistochemical works have identified the presence of PACAP38 peptide in various areas of teleostian brain (28, 51, 52). However, the coexistence of PACAP and PAC1-R in the brain and in specific neural or nonneural cells is uncertain. In our previous study, isolated full-length cDNAs encoded GHRH-PACAP from the brain of tilapia. The results of this present investigation verify the expression of GHRH-PACAP gene in the brain and pituitary of tilapia. Moreover, this is the first study to demonstrate the presence of GHRH-PACAP cDNA (tGHRH-tPACAP38) in tilapia astrocytes. This work also shows that PAC1-R is expressed concurrently in tilapia astrocytes. Three distinct subtypes of receptors possess differential affinities for PACAP and VIP: PAC1 receptors (PAC1-R) show high affinity for PACAP and a much lower affinity for VIP, whereas VPAC1 and VPAC2 receptors (VPAC1-R and VPAC2-R) recognize PACAP and VIP with high affinity (20–22). Recently, both VPAC1-R and VPAC2-R were demonstrated to be present in various tissues, including brain of goldfish, zebrafish, fugu, and trout (53–55). We cannot rule out the possibility that effects of PACAP on astrocytes in the present study are mediated partially through VPAC-R. It also is possible that exogenous PACAP induced cytokine production in astrocytes and subsequently acted back to the astroglial cytokine receptor to regulate PACAP expression. In mammals, most studies have focused on PACAP and VIP mediation of cytokine (such as interleukin-6) production and release (54). Relatively few experiments have addressed the regulation of PACAP by cytokines. Several studies indicate that VIP and certain other neuropeptides are regulated by leukemia inhibitory factor and other cytokines (56). The effects of exogenous or endogenous PACAP, exerting in an autocrine/paracrine manner, on the VPAC-R of tilapia astrocytes or the interaction of PACAP and cytokine in tilapia astrocytes deserves to be further explored.
The tGHRH-tPACAP38 cDNA regions are those in other nonmammals as determined and described elsewhere (15). They are 5′ UTR, signal peptide, cryptic peptide, GHRH, PACAP, and 3′ UTR. RT-PCR analysis revealed two products of different sizes in tilapia brain, pituitary, and astrocyte. Sequence analysis of the PCR products showed that the large encodes both tGHRH and tPACAP38, whereas the small product encodes only complete tPACAP38 and some C-terminal amino acids of tGHRH, from which most of the tGHRH molecule is deleted. The shorter PCR product of tGHRH-tPACAP38 is formed by exon skipping, a particular form of alternative splicing in which an exon is deleted during the formation of mRNA. This process has been shown to occur in lower vertebrates, including various species of fish (4, 6, 29, 57, 58), frog (10), and bird (11, 12). Mammals control the production of GHRH and PACAP by encoding these two peptides in separate genes. In the tilapia of interest here, exon 4 contains the tGHRH(1–32 aa) that is deleted, allowing tPACAP38 to be expressed independently of tGHRH. The deleted region corresponds to exon 4 of the rainbow trout GRF-PACAP gene (58). Exon skipping in the GHRH-PACAP mRNA probably represents the ancestral regulation of GHRH and PACAP production, increasing the ratio of PACAP mRNA to GHRH mRNA. However, the factors that regulate exon skipping of the GHRH-PACAP transcripts are not well understood. Several studies have reported that this mechanism operates as early as Embryonic Day 4 in fish (58) and developing turkeys (12), so the ratios of PACAP to GHRH-PACAP transcripts vary. These results also may support the findings that PACAP in fish more strongly promotes the secretion of GH and gonadotropins than does GHRH (28, 29). Alternative processing is an effective means of regulating the ratio of peptides from multipeptide coding genes in the PACAP/glucagon superfamily. In fish and birds, the proglucagon gene is alternatively spliced in a tissue-specific manner to generate either glucagon or glucagonlike peptides 1 and 2 (59). In birds, the vasoactive intestinal polypeptide can be formed without peptide histidine isoleucine (PHI) when the exon encoding of the latter is skipped (60). However, no evidence supports exon skipping in the mammalian vasoactive intestinal polypeptide transcript (61). These examples reveal that tissue-specific factors may activate exon skipping.
The expression of tilapia tPACAP38 in astrocytes suggests a local role for peptides in regulating astrocytic functions. Locally generated PACAP acts as an autocrine and paracrine hormone such as a neurotransmitter or a neuromodulator in various organs (14). In follicle cells in zebrafish, PACAP acts as autocrine/paracrine in follicular maturation (38). As more evidence shows that PACAP is a potential local factor, understanding how it is regulated in astrocytes becomes increasingly important. In this study we verified that PACAP coexisted with PAC1-R in tilapia astrocytes, suggesting that PACAP acts on tilapia astrocytes in an autocrine/paracrine manner. Evidence obtained here demonstrates that exogenous ovine PACAP38 strongly upregulates tPACAP38 in the tilapia astrocytes, which is consistent with the assumption that PACAP mediates neuromodulators. Like exogenous ovine PACAP38, db-cAMP significantly promoted the expression of tPACAP38 in a dose-dependent manner in tilapia astrocytes. Unlike tPACAP38, PAC1-R in tilapia astrocytes did not respond to exogenous ovine PACAP38 at the mRNA level, as in a previous report that found that a reproductive hormone (gonadotropin) in rats does not affect PAC1-R expression in granulosa cells (62). However, the expression of PAC1-R in this work verified that PAC1-R is present in astrocytes (63). Similar time course response (Fig. 5), as short as 2 hrs, the upregulations of PACAP mRNA in tilapia astrocytes were observed by dbcAMP and PACAP treatment. The upregulation of tPACAP38 in response to db-cAMP treatment suggests that the activation of the cAMP-PKA signaling pathway dominantly mediates the upregulation of tPA-CAP38 expression by exogenous ovine PACAP38. These data are consistent with the results of other studies that found the presence of two cAMP response elements in the promoter of rat and mouse PACAP genes (64, 65) as well as with works that found that PACAP-induced differentiation is mediated by the production of cAMP in such cells as neuroblastoma (66), pheochromocytoma (67), cerebella granule (68), and embryonic stem (69) cells. This investigation verified that PACAP also may generate cAMP in tilapia astrocytes (data not shown) and in rat astrocytes (70). The strong upregulation of tPACAP38 by added exogenous ovine PACAP38 and db-cAMP reveals that PACAP in astrocytes acts as a paracrine communicator between an astrocyte (PACAP) and another astrocyte (PAC1-R) or in an autocrine manner in astrocyte itself, (PACAP and PAC1-R colocalization) to mediate the function of vertebrate astrocytes.
In rodents, PACAP has been found to have multiple functions, including the regulation of pituitary hormone release, intestinal motility, neuronal action, and insulin release, as well as cell division and differentiation (14). In addition, PACAP affects many defined behaviors, such as feeding behavior, exploratory activity, locomotor activity, face washing, body grooming, drinking and rearing behaviors, and rapid eye movement sleep when injected into the hypothalamic paraventricular nucleus or third ventricle, and there are more studies of feeding behavior in fish (55). Our preliminary data showed that the expression level of tPACAP38 mRNA in tilapia brain was significantly increased at 8 hrs after transfer from FW to SW. We have postulated that PACAP may act in a paracrine and/or autocrine manner to mediate the in vivo effects of hyperosmolarity on brain PACAP expression. However, the role of PACAP in the osmoregulation of tilapia, particularly in brain and astrocytes, is becoming more interesting. Earlier investigations have reported that cells respond to changes in osmotic stress with compensatory molecular adaptations, enabling them to reestablish the homeostasis of osmotically disturbed aspects of cell structure and function (71). Several studies have demonstrated that cultured astrocytes under various osmotic conditions exhibit augmentation of water channel mRNA and protein expression (43); expression of osmolytes, synthetic enzymes, and transporter (44); and activation of p38 kinase (45) in response to hyperosmotic stress accompanied by a change in the regulated volume through the chloride channel (46). Whereas the triggering of a calcium oscillation (47) in response to a hypoosmotic challenge has been reported. The effects of NaCl and mannitol on tilapia astrocytes were further examined to elucidate the involvement of osmolarity and ions in the osmoregulation of tilapia during SW transfer. As shown in Figures 6 and 7, the expressed level of PACAP mRNA in astrocytes was increased significantly up to 16 hrs after exposure to NaCl. A significant increase in protein content of PACAP was seen at 4 hrs. At 8 hrs and 16 hrs after NaCl exposure, the PACAP protein content in astrocytes remained higher, although it was not significantly different from the control. This discrepancy could be due to an alteration at translational level. Previous reports from experiments with Xenopus oocytes have demonstrated that RNA splicing can influence subsequent events in the RNA processing pathway and result in change of translational yields (72). When mammalian cells (COS7 cells) are exposed to a variety of environmental stresses, translation-ally inactive messenger ribonucleoprotein particle (mRNP), 40S ribosomal subunits, and the mRNA binding proteins TIA-1 and TIAR in cytoplasm is formed and results in translational arrest of transcripts (73, 74). However, there no such reports have demonstrated stress-related changes of cytoplasmic processing bodies in teleostian (tilapia). The underlying mechanism of our observation remains to be determined. This study is the first to show that NaCl treatment but not mannitol treatment significantly promoted the expression of PACAP in tilapia astrocytes, demonstrating that ions (Na+ and Cl−) under hyperosmolarity conditions induce the expression of tPACAP38.
In conclusion, PACAP expresses in tilapia astrocytes and is upregulated by hyperosmolarity, and it subsequently acts on its own receptor by an autocrine and/or paracrine method. This fact suggests that PACAP may be involved in regulating the adaptation of tilapia to SW.
Primer Sequence and Expected Lengths of Fragments in RT-PCR Analysis of Selected Genes in Tilapia and β-Actin a

Tilapia GHRH/PACAP cDNA. (A) Full-length nucleotide sequence (GenBank accession no. AY522580) and deduced amino acid sequence. The deduced amino acids constitute the regions that encode tGHRH (double underlined) and tPACAP38 (bold underlined). The gray background area in the coding region is the sequence deleted from the short spliced form of the transcript. The poly-adenylation signal (attaaa) in the 3′ untranslational region is underlined with a wavy line, and an asterisk marks the terminal codon. (B) Distribution of tPACAP38 in tilapia brain, pituitary, and astrocyte. EtPACAP38 primers were used to perform RT-PCR, yielding two bands. The large band (271 bp) encodes tGHRH and tPACAP38, and the small band (166 bp) encodes only tPACAP38. +, RT;-, RNA alone, without RT. The PCR-amplified DNA was separated and transferred to a nylon membrane hybridized with a cDNA clone of tGHRH-tPACAP38 to form the probe. (C) Structures of long and short forms. The long form comprises GHRH and PACAP, and the short form comprises PACAP alone. From the shadow region 105 nucleotides were deleted, including most of the GHRH, during exon skipping. SP and Cryptic are abbreviations for the signal peptide and the cryptic peptide, respectively.
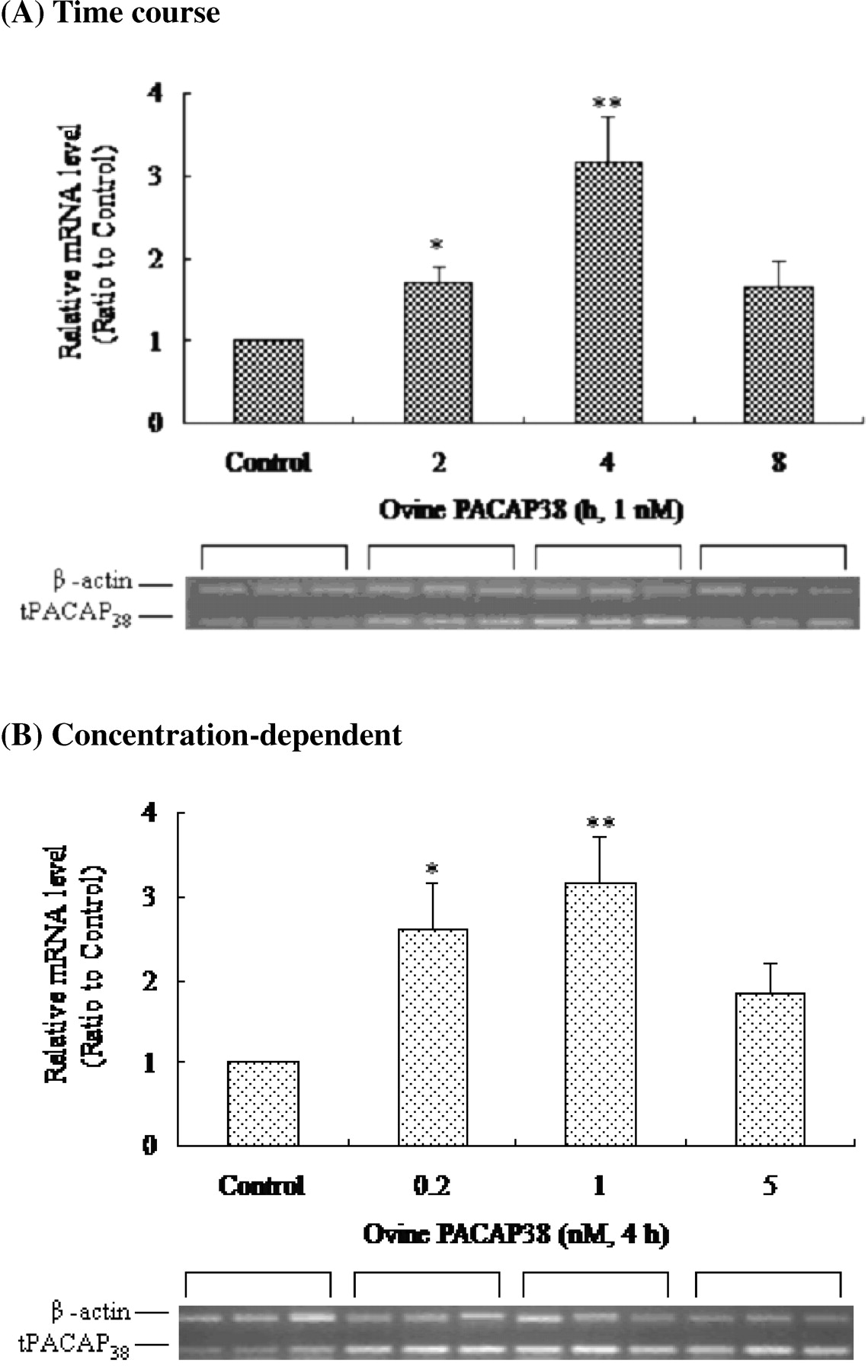
Effects of exogenous ovine PACAP38 on the expression of endogenous tPACAP38 in cultured astrocyte of tilapia brain. (A) In the time course experiment, exogenous ovine PACAP38 (1 nM) was applied to the cultured astrocyte for various periods (0–8 hrs) before RNA was extracted. (B) In the concentration-response experiment, the cells were treated for 4 hrs with various concentrations of exogenous ovine PACAP38 (0–5 nM). The data obtained after β-actin normalization were presented as ratios with the data for the control. Three independent experiments were performed in triplicates. Asterisks (* and **) represent significant differences (P < 0.05 and P < 0.01) among treatments.

Effects of db-cAMP on the expression of endogenous tPACAP38 in cultured astrocyte of tilapia brain. (A) In the time course experiment, the cells were treated by db-cAMP (1.5 mM ) for various periods (0–8 hrs). (B) In the concentration-response experiment, the cells were treated for 2 hrs with various concentrations of db-cAMP. The data obtained after β-actin normalization were presented as ratios with the data for the control. **A significant difference (P < 0.01) among treatments.

(A) Partial nucleotide sequence (446 bp) of PAC1-R. (B) Presence of PAC1-R in tilapia brain, pituitary, and astrocyte. (C) Effect of exogenous ovine PACAP38 on the expression of PAC1-R in cultured astrocyte. The data obtained after β-actin normalization were presented as ratios with the data for the control. Three independent experiments were performed in triplicate.

(A) Morphology of astrocyte in light field with GFAP staining. Magnifications: ×300 and ×600. (B) Immunocytochemistry of PACAP and PAC1-R in cultured astrocytes treated with exogenous ovine PACAP38 (1 nM, 4 hrs) and db-cAMP (1.5 mM, 2 hrs). Magnification: ×600. The arrows indicate signals with various densities. (C) Western blot analysis of PACAP (3.8 kDa) and PAC1-R (50 kDa) in cultured astrocytes treated with exogenous ovine PACAP38 (1 nM) and db-cAMP (1.5 mM) for various periods (0, 2, 4, or 8 hrs). Three independent experiments were performed in triplicate. Asterisks (* and **) represent significant differences (P < 0.05 and P < 0.01) among treatments. Color version of this figure is available in the on-line journal.

Effects of NaCl and mannitol (hyperosmolarity) on the expression of (A) tPACAP38 and (B) PAC1-R in cultured astrocytes of tilapia brain. The cells were treated with hyperosmolarity medium by adding NaCl and mannitol for various periods (0, 4, 8, or 16 hrs). The data obtained after β-actin normalization were presented as ratios with the data for the control. Three independent experiments were performed in triplicate. Asterisks (* and **) represent significant differences (P < 0.05 and P < 0.01) among treatments.

(A) Immunocytochemistry of PACAP and PAC1-R in cultured astrocytes treated with NaCl- and mannitol-hyperosmolarity medium (4 hrs). Magnification: ×600. The arrows indicate signals with different densities. (B) Western blot analysis of PACAP (3.8 kDa) and PAC1-R (50 kDa) in cultured astrocytes treated with NaCl and mannitol for various periods (0, 4, 8, or 16 hrs). Three independent experiments were performed in triplicate. *Significant difference (P < 0.05) among treatments. Color version of this figure is available in the on-line journal.
Footnotes
This work was supported by the National Science Council of the Republic of China, Taiwan, through contract NSC 91-2311-B-259-001.