
Abstract
Endothelin-1 (ET-1) acts on two different G protein–coupled receptors, namely the endothelin A (ETA) and the endothelin B (ETB) receptors. Both receptor subtypes show differences in their tissue expression and signal transduction. In the present study, we compared the ability of ETA and ETB receptors to stimulate extracellular signal-regulated kinase 1/2 (ERK1/2). In addition, we analyzed the role of the extracellular N terminus for ERK1/2 activation, because the ETB receptor undergoes an agonist-dependent N-terminal proteolysis. ET-1 stimulation of HEK293 cells stably expressing the ETA receptor induced a monophasic, but sustained ERK1/2 activation, whereas the ETB receptor showed a biphasic ERK1/2 activation. A truncated mutant ETB receptor, lacking the proteolytically cleaved N terminus (Δ2-64 ETB) revealed only a monophasic and transient ERK1/2 activation. Treatment of HEK293 Δ2-64 ETB cell clones with ET-1 and a synthetic NT27-64 peptide, corresponding to the N-terminally cleaved fragment of the ETB receptor and ET-1, did not restore the biphasic activation of ERK1/2. A chimeric ETB receptor in which the N terminus was replaced by the N terminus of the ETA receptor elicited biphasic ERK1/2 activation. The presented data suggest that an intact N terminus of the ETB receptor is necessary for the second phase of ERK1/2 activation. However, it appears that the length of the N terminus rather than a specific sequence motif is required for biphasic ERK1/2 activation.
Introduction
Endothelins (ET-1, ET-2, and ET-3) are vasoactive peptides (1), which exert their action via two G protein–coupled receptors: the endothelin A (ETA) and the endothelin B (ETB) receptor (2, 3). The ETA receptor is mainly expressed in vascular smooth muscle cells, and its stimulation induces a long-lasting vasoconstriction (4). The ETB receptor is predominantly expressed in endothelial cells, and its activation causes a transient vasodilation (5). Although both endothelin receptors share 64% amino acid sequence identity, marked differences in the activation of G proteins exist. Whereas the ETA receptor activates G proteins of the Gq/11 and G12/13, the ETB receptor stimulates G proteins of the Gi and Gq/11 families (6–8). In addition, the ETB receptor displays several unique features that are not shared by the ETA receptor. For example, only the ETB receptor undergoes an agonist-dependent proteolysis of its N terminus. Although both endothelin receptor subtypes possess a cleavable signal peptide (ETA receptor: 16 aa, ETB receptor: 26 aa) that is removed by a signal peptidase in the endoplasmic reticulum lumen during receptor biosynthesis (9–11), only the ETB receptor reveals a further proteolytic cleavage within the extracellular N terminus (10). This cleavage occurs 38 amino acids C-terminally of the signal peptidase cleavage site, between arginine 64 and serine 65 (R64/S65) and is mediated by a metalloprotease in an agonist-dependent manner (12). Because the proteolytically released N-terminal peptide fragment harbors the only N-linked glycosylation site (N59/A60/S61), agonist-mediated proteolysis results in a truncated, nonglycosylated receptor. However, the physiologic significance of this process remains elusive.
Both, ETA and ETB receptors lead to an activation of extracellular signal-regulated kinase 1/2 (ERK1/2). Because ERK1/2 is important regulator of cellular proliferation, migration, and differentiation, we analyzed whether ETA and ETB receptors differ in their ability to activate ERK1/2. In addition, we studied whether N-terminal proteolysis of the ETB receptor is involved in ERK1/2 activation.
Materials and Methods
Peptide Synthesis.
Synthesis of ET-1 and of the NT27-64 peptide (corresponds to the N-terminal fragment of the ETB receptor released by a metalloprotease) were carried out by the solid-phase, standard 9-fluorenylmethoxy-carbonyl chemistry method as described recently (13).
Generation of ETB Receptor/GFP Expression Constructs.
The plasmids pcDNA3.ETB, pEGFP.ETB, and pΔ2-64.ETB encoding the human full-length
ETB receptor, the human ETB/GFP fusion protein, and a mutant
ETB receptor with a truncated extracellular N terminus (corresponding to the
N-terminally cleaved ETB receptor), respectively, have been described
previously (12, 13). Plasmids encoding a
chimeric ETA receptor in which the N terminus is replaced by the N terminus of
the ETB receptor (ANB·GFP), and a chimeric ETB receptor in which the
N terminus is replaced by that of the ETA receptor (BNA·GFP) were generated by
site-directed mutagenesis of the pEGFP.ETB and pEGFP.ETA plasmids (13). A SnaBI (underlined) site in the coding region of both
plasmids was introduced using the following primers: for the ETB receptor:
5′-GAGA CTTT CAAA
Cell Culture and Transient Transfections.
HEK293 cell clones stably expressing the ETB·GFP, BNA·GFP, ANB·GFP, or the Δ2-64 ETB·GFP receptor were maintained as described (12). Growth arrest was induced by culturing cells for 48 hrs in a serum-free quiescent medium. The protocol for the transient transfection of HEK293 cells was as described previously (14).
Immunoblots.
Detection of total ERK1/2 and phospho-ERK1/2 in immunoblots was performed as described (12, 15). In brief, HEK293 cells were grown on 35-mm Petri dishes, stimulated with ET-1 (100 nM) for up to 3 hrs and finally lysed in Laemmli buffer. Proteins were separated on 10% sodium dodecyl sulfate-polyacrylamide gels, blotted to nitrocellulose filters (Schleicher und Schuell, Dassel, Germany) and probed with affinity-purified polyclonal phospho-ERK1/2 (detects phosphoERK1/2) or ERK1/2 antibodies (detects phosphorylated and non-phosphorylated forms of ERK1/2) (both 1:1000) (Cell Signaling Technology, Danvers, MA).
Results and Discussion
To analyze whether ETA and ETB receptors differ in their ability to activate ERK1/2, we studied HEK293 cells stably expressing the full-length ETB·GFP receptor or the ETA·GFP receptor. HEK293 cells were chosen for these experiments because nontransfected cells lack endogenous expression of endothelin receptors (no specific binding of 125I-ET-1 and no ET-1–induced ERK1/2 activation; data not shown). Cell clones expressing ETA·GFP or ETB·GFP receptors were stimulated with ET-1 for up to 3 hrs, then lysed, and samples were subjected to immunoblot analysis using phosphoERK1/2 and ERK1/2 antibodies. Lysates derived from HEK293 cells expressing ETA·GFP showed a monophasic, but sustained increase in ERK1/2 phosphorylation (Fig. 1A). For lysates derived from cells expressing the ETB·GFP receptor, a biphasic ERK1/2 activation was found (Fig. 1B). The early phase of ERK1/2 activation reached a peak at 3–5 min and slowly declined thereafter within 30–60 min. The second phase was observed after 80 min of agonist application and persisted for up to 180 min (Fig. 1B). To study the role of the N-terminal proteolysis on ERK1/2 activation of the ETB receptor, we also analyzed ERK1/2 phosphorylation in HEK293 cells stably expressing the Δ2-64 ETB·GFP receptor. ET-1 stimulation of Δ2-64 ETB·GFP did not show biphasic, but only monophasic ERK1/2 activation (Fig. 1C).
Because only the full-length ETB receptor induced biphasic ERK1/2 activation, the presence of the N-terminal fragment could provide a key for switching from biphasic to monophasic ERK1/2 activation. To elucidate, whether the proteolytically released N-terminal peptide functions as a soluble ligand or coactivator in ET-1–induced ERK1/2 activation, HEK293 cell clones expressing the Δ2-64 ETB·GFP receptor were incubated with a synthetic peptide (NT27-64, 100 μM; Fig. 2A). This peptide corresponds to the proteolytically cleaved N-terminal fragment of the ETB receptor, with the exception that it lacks an N-linked glycosylation at asparagine 59. Treatment of HEK Δ2-64 ETB·GFP cell clones with the synthetic NT27-64 peptide and ET-1 did not restore the biphasic activation of ERK1/2 (Fig. 2A).
To study whether the length of the ETB receptor or a certain amino acid composition within the ETB receptor’s N terminus is required for biphasic ERK1/2 activation, we analyzed a chimeric ETB receptor in which the extracellular N terminus was replaced by that of the ETA receptor (BNA·GFP) and also a chimeric ETA receptor in which the N terminus was replaced by that of the ETB receptor (ANB·GFP). The BNA·GFP chimera induced a biphasic ERK1/2 phosphorylation on ET-1 stimulation (Fig. 2B), similar to that seen for the wild-type ETB receptor, and the ANB·GFP chimera elicited a monophasic, but sustained ERK1/2 activation as observed for the wild-type ETA receptor (Fig. 2C). The data suggest that the intact, extracellular N terminus of the ETB receptor is involved in biphasic ERK1/2 activation. Curiously, replacement of the extracellular N terminus of the ETB receptor by the N terminus of the ETA receptor did not alter biphasic ERK1/2 activation. This is notable because the extracellular N termini of the ETA and ETB receptors display only a little amino acid sequence identity.
The data demonstrate that ETA and ETB receptors differ considerably in their ability to stimulate ERK1/2 activation. Whereas the ETA receptor shows a monophasic, but sustained ERK1/2 activation, the ETB receptor elicits a biphasic ERK1/2 activation. Although an intact N terminus of the ETB receptor is necessary for biphasic ERK1/2 activation, the extracellular ETB receptor’s N terminus itself is not sufficient for the second phase of ERK1/2 activation, because it can be replaced by the N terminus of the ETA receptor. Regarding the low level of amino acid identity between the N termini of the ETA and the ETB receptor, it appears that the length of the N terminus rather than a specific sequence motif is required for biphasic ERK1/2 activation. In future experiments, the mechanisms involved in biphasic ERK1/2 activation of the full-length ETB receptor have to be studied in more detail. In addition, further studies are planned to demonstrate whether differences in the pattern of ERK1/2 activation between ETA and ETB receptors also result in different cellular responses, such as proliferation and differentiation. This is highlighted by studies on the pheochromocytoma cell line PC12: whereas treatment of these cells with epidermal growth factor resulted in monophasic ERK1/2 activation and proliferation, incubation of cells with nerve growth factor elicited biphasic ERK1/2 activation and differentiation (16).
ETA and ETB receptors show differences in ET-1–mediated ERK1/2
activation. HEK293 cell clones expressing either ETA·GFP (A) the
full-length ETB·GFP (B), or the N-terminally truncated ETB·GFP
receptor (Δ2-64 ETB·GFP; C) were stimulated with 100 nM
ET-1 for up to 180 min and finally lysed. Proteins were then separated by 10% sodium
dodecyl sulfate-polyacrylamide gels, transferred to filters, and then probed with
antiphosphoERK1/2 and anti-ERK1/2 antibodies. Shown are representative results of at
least three independent experiments. The second phase of ERK1/2 activation requires the presence of glycosylated
endothelin receptors N terminus. HEK293 cell clones expressing the N-terminally
truncated mutant Δ2-64 ETB·GFP (A) were incubated with a synthetic,
nonglycosylated N-terminal peptide (NT27-64, 100 μM) for 30 mins
before the stimulation of cells with ET-1 for up to 180 mins. HEK293 cells stably
expressing a chimeric ETB receptor, in which the N-terminus was replaced by
that of the ETA receptor (BNA·GFP) (B) or a chimeric ETA
receptor, in which the N terminus was replaced by that of the ETB receptor
(ANB·GFP) (C) were stimulated with 100 nM ET- 1 for the indicated
times. Lysates of cells were processed for immunoblotting as described in the legend
to Figure 1.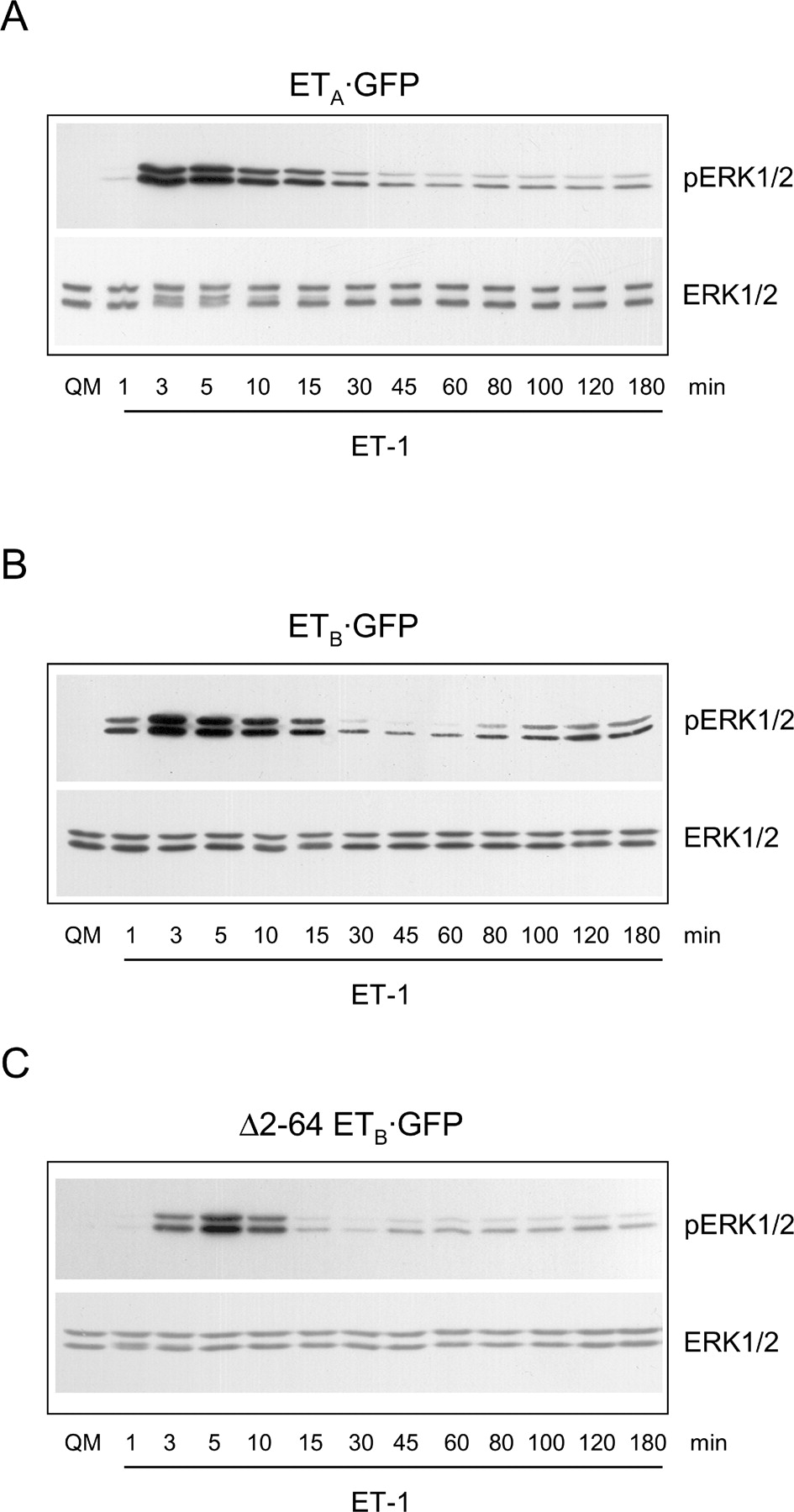

Footnotes
Supported by the Deutsche Forschungsgemeinschaft (FG 341), the Sonnenfeld-Stiftung, and the Fonds der Chemischen Industrie.
Acknowledgements
We thank Dr. Gisela Papsdorf for the generation of HEK293 clones and Monika Bigalke for technical assistance.