
Abstract
The effects of calcium channel blockers (CCBs) on complications associated with diabetes mellitus (DM) have been well studied in clinical and basic science investigations. Cardiovascular complications are a common feature of type 2 DM, and insulin resistance is an early clinical manifestation of type 2 DM. CCBs are widely used to treat cardiovascular diseases in patients with DM. In this study, we used a spontaneous type 2 diabetic rat model, Otsuka Long-Evans Tokushima Fatty (OLETF) rats, at a highly insulin-resistant stage with modest hyperglycemia. We examined cardiac expression of transforming growth factor–β1 (TGFβ1) and endothelin-1 (ET-1) in male OLETF rats. At 8 weeks of age, OLETF rats were treated for 12 weeks with the long-acting CCB benidipine (1 mg/kg/day or 3 mg/kg/day, po, n = 12), with hydralazine hydrochloride (3 mg/kg/day, po, n = 12), or with vehicle (OLETF, n = 12), and male age-matched genetic control Long-Evans Tokushima Otsuka (LETO, n = 12) rats were used. Blood pressure was significantly higher in OLETF rats than in LETO rats, and benidipine treatment at both dosages in OLETF rats for 12 weeks did not significantly reduce blood pressure, whereas hydralazine treatment significantly lowered blood pressure in OLETF rats. Hydralazine and both dosages of benidipine significantly reduced upregulated cardiac ET-1 levels in OLETF rats. Plasma and cardiac TGFβ1 levels were remarkably higher in OLETF rats compared with LETO rats and were normalized by treatment with benidipine (3 mg/kg/day). Our results suggest that CCBs are effective in normalizing upregulated cardiac TGFβ1 and ET-1 levels at the insulin-resistant stage in OLETF rats, which may improve cardiac morphology and function in this rat model without altering blood pressure and plasma glucose levels. In contrast, hydralazine treatment also normalizes cardiac ET-1 levels while significantly reducing blood pressure.
The Otsuka Long-Evans Tokushima Fatty (OLETF) rat is an established genetic model of human Type II diabetes mellitus (DM) (1). Cardiac remodeling, including cardiac fibrosis, occurs during an early stage of DM development in this strain, and altered left ventricular (LV) diastolic function has been demonstrated (2). The alteration includes an increase in deceleration time and a decrease in peak velocity amplitude of the early diastolic filling wave (2). This alteration can be tracked longitudinally using transthoracic Doppler echocardiography and is accompanied by serologic and pathologic changes in the heart (2). These changes may be a result of metabolic abnormalities or may be simultaneous alterations occurring parallel to the metabolic abnormalities in prediabetes. Previous studies using this rat strain demonstrated that cardiac fibrosis and hypertrophy are evident at the insulin-resistant stage in this model and that these changes are reversed by type-1 angiotensin II receptor blockers and by calcium channel blockers (CCBs) (3–5). LV diastolic function is impaired beginning at age 20 weeks in untreated OLETF rats (6). Early treatment with pravastatin sodium prevents cardiovascular remodeling in OLETF rats by retarding progression of glucose intolerance, by overexpressing cardiac endothelial nitric oxide synthase, and by inhibiting overexpression of fibrogenic and proinflammatory cytokines (6).
The endothelin (ET) system is composed of 3 active ETs (7). ET-1 is the main isoform, which is synthesized in endothelial cells, in the muscular coating of the arterial wall, and in the heart, kidney, and central nervous system (8). ET-1 is the predominant isoform in the cardiovascular system (8). Evidence suggests that ET-1 plays an important role in the pathogenesis of cardiovascular diseases, including congestive heart failure and coronary artery disease, through multiple mechanisms (9). ET-1 has mitogenic properties, causing proliferation and hypertrophy of vascular smooth muscle, cardiac myocytes, and fibroblasts (7).
Transforming growth factor–β1 (TGFβ1) facilitates fibrous tissue formation, upregulates collagen expression by stimulating extracellular matrix synthesis (10), and mediates perivascular and myocardial fibrosis. The increased gene expression of extracellular matrix components, such as collagen, and of TGFβ1 causes cardiac fibrosis and promotes cardiac stiffness, leading to diastolic dysfunction (11). TGFβ1 upregulation was recently demonstrated in OLETF rat heart at 30 weeks of age, and this upregulation was normalized by treatment with pravastatin (6).
Calcium channel blockers (CCBs) are widely used to treat cardiovascular diseases. In DM-related vascular hypertrophy in the absence of hypertension, these agents confer vascular protection similar to that observed with renin-angiotensin system blockers (12). Compared with placebo, dihydropyridine CCBs (long-acting nifedipine and nitrendipine) reduce the risk for cardiovascular end points and significantly decrease the risk for strokes and cardiovascular and total mortality relative to other hypertension treatment (13). A reduced risk has also been observed in patients with DM when treated with CCBs compared with placebo (13). Treatment with the antihypertensive drug hydralazine hydrochloride does not affect the hyperglycemic state of streptozotocin-induced diabetic rats (14). Moreover, although lacidipine (a CCB) and hydralazine normalized blood pressure, only lacidipine was effective in reducing renal and cardiac damage in rats fed a high-fructose diet with hypertriglyceridemia, hyperinsulinemia, and hypertension (15).
The present study was designed to investigate the effects of benidipine at different dosages (1 mg/kg/day and 3 mg/kg/day) and of hydralazine (3 mg/kg/day) on cardiac ET-1 levels in OLETF rats at age 20 weeks. Moreover, plasma and cardiac expression of TGFβ1 was examined in cases in which there was already coronary perivascular fibrosis and modest cardiomyocyte hypertrophy, and the effects of benidipine on plasma and cardiac TGFβ1 levels were evaluated. Finally, the effects of benidipine and hydralazine on blood pressure changes were studied.
Materials and Methods
Animals and Drug Treatment.
The experimental design was approved by the Hokkaido University School of Medicine Animal Care and Use Committee. Animals were obtained from the Tokushima Research Institute of Otsuka Pharmaceutical (Tokushima, Japan) and were maintained under constant temperature and lighting conditions, with free access to food and water. At 8 weeks of age, male OLETF rats were randomly divided into four groups with 12 animals each. Two groups were administered benidipine (Kyowa Hakko Kogyo Co., Ltd., Tokyo, Japan) at a per-oral dosage of 1 mg/kg/day or 3 mg/kg/day in 0.3% carboxymethyl cellulose solution, a third group received hydralazine (3 mg/kg/day, po), and a fourth group received vehicle. The treatment period was 12 weeks, and the animals were evaluated at age 20 weeks, when they were in the early stage of type 2 DM with modest hyperglycemia and prominent insulin resistance. Twelve male aged-matched genetic control Long-Evans Tokushima Otsuka (LETO) rats, which developed from the same colony by selective mating but did not develop DM, served as controls. On the day of the experiments, the animals were humanely killed under gaseous ether anesthesia. The hearts were rapidly excised, and the whole-heart and LV weights were measured. An LV portion was dipped into optimum cutting temperature (OCT) compound (Sakura Finetechnical, Tokyo, Japan) and immediately frozen in liquid nitrogen. The remaining portion was preserved at −80°C without OCT compound.
Enzyme Immunoassay for LV ET-1.
The concentration of ET-1 in LV tissue extracts was determined using an ET-1 enzyme immunoassay kit (Immuno-Biological Laboratories, Fujioka, Japan). This kit is a solid-phase sandwich enzyme-linked immunoabsorbent assay (ELISA) using 2 kinds of high-specific antibodies. Tetraethyl benzidine is used as a coloring agent (chromogen), the strength of which is in proportion to the quantities of ET-1. In LV tissue extract, an equal volume of 0.1% trifluoroacetic acid (TFA) in water was added. The sample was then centrifuged at 3000 g for 15 mins at 4°C for clarification, and the supernatant was saved. After a 200-mg C18 Sep-Pak column (Part 235D1; Waters Inc., Milford, MA) was washed with 4×1 ml of 60% acetonitrile in 0.1% TFA followed by 4×5 ml of 0.1% TFA in water, the supernatant was applied to the column and was washed with 4×5 ml of 0.1% TFA in water. The sample was eluted slowly by applying 3×1 ml of 60% acetonitrile in 0.1% TFA in water, and the eluant was collected in a plastic tube. The eluant was then evaporated to dryness using a centrifugal concentrator under vacuum. The sample was reconstituted with assay buffer and was measured immediately according to the manufacturer’s instructions.
Western Blot Analysis.
Immunoblotting was performed in the LV tissues of experimental animals. Samples of tissue homogenate were subjected to electrophoresis on polyacrylamide gels, and proteins were transferred to polyvinylidine difluoride filter membrane. After blocking with 5% nonfat milk in phosphate-buffered saline, the membranes were incubated with human anti-TGFβ1 (R&D Systems, Minneapolis, MN). After extensive washing with phosphate-buffered saline containing 0.1% Tween 20 to remove any nonspecifically bound primary antibodies, the membranes were incubated with horseradish peroxidase–conjugated antirabbit antibody. Blots were visualized using an enhanced chemiluminescence detection system (Amersham Biosciences UK, Little Chalfont, Buckinghamshire, UK), exposed to x-ray film, and analyzed using free image software from the National Institutes of Health (Bethesda, MD).
Enzyme Immunoassay for TGFβ1.
The concentration of TGFβ1 in plasma and tissue extracts was determined using a multispecies TGFβ1 ELISA kit (Bio-Source International, Camarillo, CA). This assay recognizes only active TGFβ1, and no significant cross-reactivity or interference was observed. The assay procedure was carried out according to the manufacturer’s instructions.
RNA Preparation and Quantitative Real-Time Polymerase Chain Reaction (RT-PCR).
Total tissue RNA was isolated by acid guanidinium thiocyanate-phenol-chloroform extraction with Isogen (Nippon Gene Ltd., Toyama, Japan) as previously described in detail (16). Briefly, the LV tissue was homogenized in Isogen with a Polytron tissue homogenizer (Model PT10SK/35; Kinematica, Lucerne, Switzerland). Precipitated RNA was extracted with chloroform, precipitated with isopropanol, and washed with 75% (vol/vol) ethanol. Total tissue RNA was treated with DNase using an RNase-free DNase kit (Qiagen, Tokyo, Japan). The samples were then purified using a Qiagen RNeasy mini kit to improve the quality of total tissue RNA. RNA concentration was determined spectrophotometrically at 260 nm, and RNA quality was judged based on the ratio of absorbance at 260 nm and 280 nm. Total tissue RNA was primed with 0.05 μg of oligo d (pT)12–18 and reverse transcribed by omniscript reverse transcriptase using a first-strand cDNA synthesis kit (Qiagen). The reaction was performed at 37°C for 60 mins.
mRNA expression levels of TGFβ1 in the heart were analyzed by quantitative RT-PCR with TaqMan probe using an ABI Prism 7700 Sequence Detector (Perkin-Elmer Applied Biosystems, Foster, CA) as previously described (16). The gene-specific primers and TaqMan probes were synthesized using Primer Express v. 1.5 software (Perkin-Elmer Applied Biosystems). The sequences of the oligonucleotides were as follows:
TGFβ1 forward: 5′-CGGGAAGCAGTGCCAGAA-3′
TGFβ1 reverse: 5′-TGCTCCACAGTTGACTTGAATCTC-3′
TGFβ1 probe: 5′-CAGAGCTGCGCCTGCA-3′
GAPDH forward: 5′-GTGCCAAAAGGGTCATCATCTC-3′
GAPDH reverse: 5′-GGTTCACACCCATCACAAACATG-3′
GAPDH probe: 5′-TTCCGCTGATGCCCC-3′
The expression of glycolytic enzyme glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA was used as an internal control. The PCR mixture (25 μl total volume) consisted of 450 nM of forward and reverse primers for TGFβ1 and GAPDH, 200 nM of FAM-labeled primer probes, and TaqMan Universal PCR Master Mix (all from Perkin-Elmer Applied Biosystems). Each PCR amplification was performed in triplicate, using the following profile: 1 cycle of 95°C for 10 mins and 40 cycles of 94°C for 15 secs and 60°C for 1 min. For the standard curve in the quantitative RT-PCR, serial dilutions of rat heart cDNA were performed within the range of various concentrations (1×, 2×, 4×, 8×, and 16×). A no-template (water) reaction mixture was prepared as a negative control.
Statistical Analysis.
Values are means ± SE. Statistical assessment of the data was performed using one-way analysis of variance, with multiple comparisons using the Fisher protected least significant difference test. Nonparametric data were analyzed using the Mann-Whitney U test or the Wilcoxon signed rank test. P < 0.05 was considered significant.
Results
The general features of OLETF rats at age 20 weeks and those of age-matched LETO rats have been previously described (3–5). The whole-heart and LV weights of OLETF rats were significantly greater (about 1.4-fold) than those of age-matched LETO rats. Benidipine and hydralazine treatment had no effect on increased whole-heart and LV weights in OLETF rats. Plasma glucose and insulin levels were significantly higher in OLETF rats than in LETO rats regardless of whether benidipine or hydralazine was given. Plasma glucose levels were significantly higher in OLETF rats than in LETO rats at age 20 weeks (10.93 ± 0.54 vs 9.12 ± 0.20 mM, P < 0.01); therefore, OLETF rats used in the study were modestly hyperglycemic. Plasma insulin levels were much higher in OLETF rats than in LETO rats (743 ± 81 vs. 401 ± 33 pmol/l, P < 0.01).
Systolic blood pressure of OLETF rats was slightly but significantly higher than that of LETO rats (140 ± 3 vs. 129 ± 3 mm Hg, P < 0.01) (Fig. 1A). Benidipine at a dosage of 1 mg/kg/day did not significantly reduce blood pressure (137 ± 3.9 mm Hg) in OLETF rats, while benidipine at a dosage of 3 mg/kg/day lowered blood pressure in OLETF rats (132 ± 4 mm Hg), but this depressor effect was not statistically significant. In contrast, hydralazine treatment significantly reduced blood pressure in OLETF rats (122 ± 2.6 mm Hg, P < 0.001.
Immunoreactive ET-1 levels in LV tissues were significantly higher (about 1.7-fold) in OLETF rats than in LETO rats, and benidipine treatment at both dosages reduced immunoreactive ET-1 levels in LV tissues to the same extent in OLETF rats as in LETO rats (Fig. 1B). Moreover, hydralazine treatment normalized upregulated cardiac ET-1 levels in OLETF rats.
A significant increase in TGFβ1 levels as determined by ELISA was observed in plasma and LV tissue samples from OLETF rats, and benidipine (3 mg/kg/day) reversed the levels to those obtained in nondiabetic LETO rats (Fig. 2A and B). Immunoblots obtained with TGFβ1 antiserum showed a single band that migrated, with an apparent molecular mass of 44 kDa (Fig. 3A). The amount of immunodetectable TGFβ1 was increased 212% ± 10% in OLETF rats relative to that of nondiabetic controls. Benidipine (3 mg/kg/day) resulted in significant reversal of the DM-induced increase in TGFβ1 protein levels (120% ± 8%). Compared with those in LETO rats, TGFβ1 mRNA levels were significantly greater in OLETF rats and were normalized in benidipine-treated OLETF rats, as determined by quantitative RT-PCR (Fig. 3B).
Discussion
In the present study, we demonstrated that upregulated cardiac ET-1 levels were normalized by benidipine at two different dosages and by hydralazine. Systolic blood pressure was significantly higher in OLETF rats compared with control LETO rats. Benidipine at both dosages did not significantly reduce blood pressure in OLETF rats, while hydralazine significantly lowered blood pressure in OLETF rats. Benidipine and hydralazine did not affect plasma glucose and insulin levels in diabetic rats. In addition, plasma and cardiac TGFβ1 levels were upregulated in OLETF rats at a highly insulin-resistant stage, and these upregulations were normalized by a subdepressor dosage of the long-acting CCB benidipine (3 mg/kg/day).
Previous work indicates that OLETF rats exhibit significant insulin resistance and impaired glucose tolerance with hyperglycemia at age 20 weeks (3–5). OLETF rats at this age showed modest hyperglycemia and prominent hyperinsulinemia after 24 hrs of fasting. Therefore, our subjects were in the type 2 DM stage of insulin resistance seen early in this strain.
In the present study, a significant increase in ET-1 levels was found in LV tissues of OLETF rats. It was previously demonstrated that (compared with LETO rats) plasma ET-1 levels were significantly higher in OLETF rats at insulin-resistant stages and were normalized by benidipine treatment (17). In our study, upregulated cardiac ET-1 levels were reversed to the levels of control LETO rats by both dosages of benidipine. In addition, the antihypertensive drug hydralazine normalized upregulated cardiac ET-1 levels in OLETF rats. ET-1 is powerfully hypertrophic in cardiac myocytes (18, 19). It also activates the Type I procollagen promoter (20) and collagen synthesis in fibroblasts (21). Via ETB receptors, ET-1 potently stimulates aldosterone release (22) and may increase collagen synthesis by enhanced aldosterone secretion. Therefore, ET-1 might also contribute to cardiac remodeling in OLETF rats.
In our study, benidipine was used at subdepressor dosages. Benidipine at a dosage of 1 mg/kg/day did not reduce blood pressure in OLETF rats, while 3 mg/kg/day tended (statistically insignificantly) to reduce blood pressure. Both dosages of benidipine normalized cardiac ET-1 levels in OLETF rats in a similar extent and pattern. In a previous study, a subdepressor dosage of benidipine (1 mg/kg/day) ameliorated the increased expression of ET-1 in LV tissues in Dahl salt-sensitive hypertensive rats fed a high-salt diet (23). Hydralazine normalized cardiac ET-1 levels in OLETF rats to the same extent as benidipine, but hydralazine also significantly reduced blood pressure. Based on these findings, the reversal effect of benidipine on upregulated cardiac ET-1 levels may be independent of blood pressure change. Normalization of upregulated cardiac ET-1 levels in OLETF rats by hydralazine might be due to its blood pressure–lowering effects.
Our study also showed that TGFβ1 was highly expressed in OLETF rat heart at the protein and mRNA levels. TGFβ1 is a powerful stimulant of cardiomyocyte hypertrophy (24) and collagen synthesis in the heart (25). This suggests that superinduction of TGFβ1 transcription may be involved in cardiac remodeling at the early insulin-resistant stage of DM in OLETF rats. TGFβ1 promotes the deposition of extracellular matrix protein and acts as an anti-inflammatory cytokine. These biologic effects might be implicated in LV remodeling and treatment failure after myocardial infarction. In a recent study, significant upregulation of TGFβ1 was observed in 30-week-old OLETF rat heart (6). This upregulation was associated with cardiac fibrosis and was reversed by pravastatin (6). The most important finding in the present study is that benidipine (3 mg/kg/day) normalized upregulated TGFβ1 levels in plasma and in cardiac tissues at a subdepressor dosage. This is similar to the previous finding that angiotensin II may inhibit reactive myocardial fibrosis in pressure-overloaded hearts, through a mechanism independent of hemodynamic change (26). Moreover, 1 mg/kg/day of benidipine inhibited the increased expression of TGFβ1 in LV tissues in Dahl salt-sensitive hypertensive rats fed a high-salt diet (23).
TGFβ1 substantially increases the expression of pre-proET-1 mRNA and ET-1 in human pulmonary arterial smooth muscle cells (27) and contributes to pulmonary hypertension. Therefore, ET-1 may partially mediate the pulmonary vascular actions of TGFβ1. Hepatic ET-1 and TGFβ1 levels increased during a similar time frame in hepatopulmonary syndrome (28). TGFβ1 stimulates cholangiocyte ET-1 expression and production, and cholangiocyte-derived ET-1 may be an important endocrine mediator in rat experimental hepatopulmonary syndrome (28). On the other hand, ET-1 has been reported to stimulate production of TGFβ1 in rat liver stellate cells (29). Furthermore, ETA receptor antagonism can prevent the elevation of cardiac TGFβ1 mRNA and protein levels in deoxycorticosterone acetate–salt hypertensive rats (30). Therefore, there may be an interregulatory system between TGFβ1 and ET-1 in diabetic heart that would contribute to the development of cardiac remodeling in OLETF rat heart at the insulin-resistant stage.
The present study showed that treatment of OLETF rats with benidipine, a long-acting CCB, at two subdepressor dosages (1 mg/kg/day and 3 mg/kg/day) inhibited increased cardiac ET-1 levels. In addition, the antihypertensive agent hydralazine normalized upregulated cardiac ET-1 levels and significantly reduced blood pressure in OLETF rats. Benidipine treatment at 3 mg/kg/day also reversed upregulated cardiac TGFβ1 levels in insulin-resistant OLETF rats. Future studies should investigate whether these normalizations of TGFβ1 and ET-1 levels by benidipine are independent of hemodynamic changes in OLETF rats or whether there might be a specific effect of CCBs on TGFβ1 and ET-1 expression in OLETF rat heart.
Blood pressure changes (A) (*P < 0.01 compared with the
corresponding values in OLETF rats without any treatment) and ET-1 levels in LV
tissues as determined by ELISA (B) (*P < 0.01 compared with the
corresponding values in OLETF rats without any treatment) in LETO, OLETF,
benidipine-treated (1 mg/kg/day and 3 mg/kg/day) OLETF rats, and hydralazine-treated
OLETF rats. Data are means ± SE (n = 12). TGFβ1 levels determined by ELISA in plasma (A) and in LV tissues (B) in
LETO, OLETF, and benidipine-treated (3 mg/kg/day) OLETF rats. Data are means ± SE
(n = 12). *P < 0.01 compared with the
corresponding values in LETO rats. #P < 0.01 compared with the
corresponding values in OLETF rats without any treatment. (A) TGFβ1 protein levels by immunoblot in LV tissues of LETO, OLETF, and
benidipine-treated (3 mg/kg/day) OLETF rats. A typical Western blot indicating an
increased expression of the 44-kDa band in DM is shown in the upper part of the panel.
(B) mRNA expression of TGFβ1 was quantitatively evaluated using RT-PCR.
Data are means ± SE (n = 12). *P < 0.01 compared
with the corresponding values in LETO rats. #P < 0.01 compared
with the corresponding values in OLETF rats without any treatment.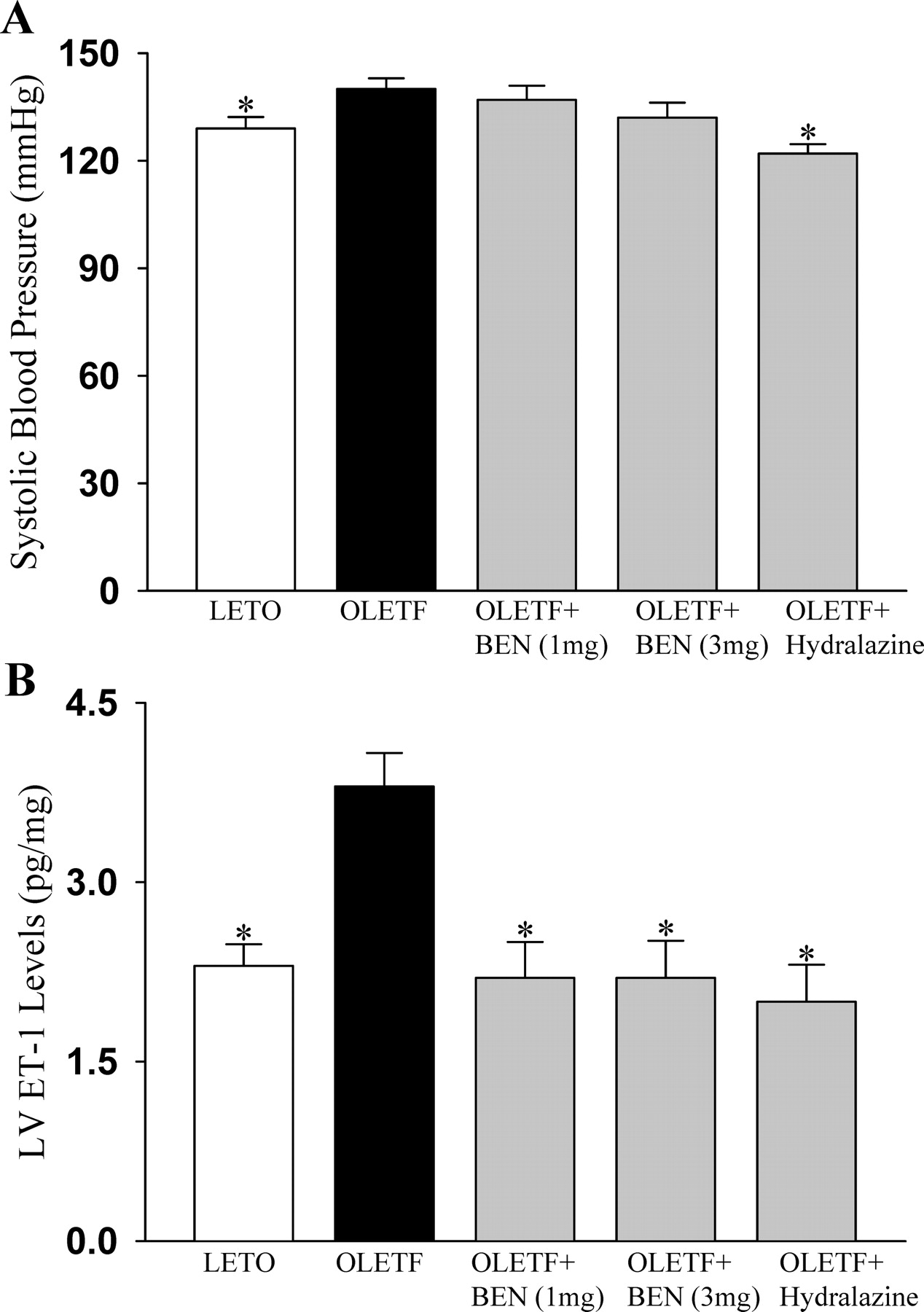


Footnotes
This study was supported in part by a grant-in-aid for scientific research from the Ministry of Education, Science, Sports and Culture of Japan; and by health sciences research grants for comprehensive research on aging and health from the Ministry of Health, Welfare and Labor of Japan.