
Abstract
During severe sepsis, several immunological defense mechanisms initiate a cascade of inflammatory events leading to multiorgan failure, including septic encephalopathy and ultimately death. Endothelin-1 (ET-1) has recently been investigated in different cerebral pathologies. Some reports suggest the involvement of ET-1 in sepsis. However, no study to date has reported the alterations in expression of the genes encoding preproET-1 and ET receptors in the frontal cortex of the septic brain. Male Sprague-Dawley (SD) rats 8 weeks of age were administered either saline or 15 mg/kg lipopolysaccharide (LPS) at different time points (1, 3, 6, and 10 hrs). Rats that did not receive LPS were considered to be controls. The rats were sacrificed with ether, and the brain tissues were harvested. Systolic and diastolic blood pressure decreased 1 hr after LPS administration and then gradually returned to normal, without any change in the heart rate. We confirmed the induction of endotoxemia in the brains of SD rats by measuring the expression of nitric oxide synthase (NOS) mRNA induced in the cerebrum. The expression of inducible NOS (iNOS) mRNA in the brains of SD rat after LPS administration was 30-fold higher than that in the brains of control rats. mRNA expression of preproET-1 in the frontal cortex of SD rats after LPS administration was 2-fold higher than that in control rats. A time-dependent increase in the expression of the gene encoding the ETA receptor (vasoconstrictive property) after LPS administration was observed in SD rat brain, whereas expression of the gene encoding the ETB receptor (vasodilatatory property) showed an initial upregulation and then gradually decreased as sepsis progressed. In conclusion, we report for the first time that expressions of the genes encoding ET-1 and ET receptors are altered in the endotoxemic brain and that these alterations are time-dependent in SD rats. The alterations in the ET system in brain tissue observed in the present study may contribute to the understanding of the pathophysiological changes in the endotoxemic brain.
Introduction
Septic shock is a severe form of sepsis that is characterized by progressive hypotension, hyporeactivity to vasopressor agents, and vascular leakage (1). Many inflammatory mediators (e.g., tumor necrosis factor-α [TNF-α], interleukin-1 [IL-1], and IL-6) are involved in the pathogenesis of shock, among them, nitric oxide (NO) (2). Because most of the studies conducted focused on peripheral organs, such as the lungs, liver, gut, and kidney, the participation and the pathophysiological alterations of the brain during sepsis seems to be less clear and poorly investigated (2, 3).
Septic shock is caused by lipopolysaccharide (LPS) and other bacterial products (4). The administration of LPS, a constituent of the gram-negative bacterial cell wall, induces inflammatory responses in cells or animals that are similar to the responses seen in septic shock, a major cause of death worldwide (4). When endotoxic shock develops, exaggerated amounts of the proinflammatory cytokines are released, resulting in refractory hypotension, tissue hypoperfusion, and organ dysfunction. Systemic LPS administration is a powerful immune challenge to brain tissue and is associated with an increase in the number of different proinflammatory cytokines in the brains of adult and neonatal rats, because different cell populations in the brain can synthesize their own set of cytokines during endotoxemia (5).
Endothelin-1 (ET-1) is an endothelium-derived peptide that exerts a wide range of effects on various tissues and cell types through interaction with the G-protein–coupled receptors for endothelin A (ETA) and endothelin B (ETB) (6). During sepsis, plasma ET-1 level has been shown to be significantly higher, and endotoxin and other microbial products that are released into the bloodstream trigger endothelial cells to increase the production of ET-1, which causes local vasoconstriction (7). The possible involvement of the ET system in human septic shock is further supported by a clear correlation between ET plasma levels and morbidity and mortality in septic patients (8). ET-1 has also been suggested to contribute to the dysfunction of several vital organ systems in septic shock (9). Increased expression of preproET-1 mRNA in the heart and lungs has been reported in endotoxemia (1). LPS administration increased preproET-1 mRNA expression by 5.5-fold in bovine aortic endothelial cells, compared with that in controls (9). Sepsis caused downregulation of ETA and ETB receptors at the level of gene expression, with increased TNF-α in the lungs but not the kidneys of Sprague-Dawley (SD) rats (10). Different studies are already underway to investigate the expression of the ET system in different organs in sepsis, both in vivo and in vitro (1).
ETs are widely distributed in the brain and peripheral organs (1). The brain tissue expresses both types of ET receptors on both neuronal and nonneuronal cells (11). The global distribution of ET and its binding sites in the brain suggests that, in addition to being a vasoconstrictor, it may be acting as an important neuropeptide in the central nervous system. ET-1 is produced in the cerebrovascular bed and plays a role in the pathophysiology of cerebral circulation in different cerebrovascular diseases, such as nonhemorrhagic cerebral infarct and cerebral vasospasm after subarachnoid hemorrhage (12). Intracisternal ET-1 in conscious rats induces a marked pressor response that causes an intense widespread reduction in cerebral blood flow (13). Intracerebral injection of ET-1 or infusion of ET-1 into cerebral arteries causes a focal necrosis in brain (14). However, little is known about the alterations in the ET system in the endotoxemic brain. Because ET-1 seems to be involved in the regulation of cerebral blood flow, we examined whether this potent endothelial vasoconstrictor peptide plays a pathophysiological role in the cerebral pathogenesis in endotoxemia.
In the present study, male SD rats were used, and a model of endotoxemia was produced by means of LPS administration. Specimens from the frontal cortex of the brain were analyzed for time-dependent alterations in the expression of the genes encoding preproET-1 and ET receptors and the gene encoding inducible nitric oxide synthase (iNOS). Before the animals were sacrificed, the systolic and diastolic blood pressure was recorded.
Materials and Methods
Animal Preparation.
Ten-week-old male SD rats (weight, 300–350 g) were obtained from Charles River Japan Inc. (Yokohama, Japan) and cared for according to the Guiding Principles for the Care and Use of Animals specified in the 1964 Helsinki Declaration. Endotoxemic shock was induced by ip administration of 15 mg/kg LPS dissolved in sterile saline from Escherichia coli 055:B5. The systolic and diastolic blood pressure were recorded before the animals were killed. Groups of animals (10 per group) were sacrificed by means of pentobarbital injection at different time points after LPS administration (1, 3, 6, and 10 hrs). The control group received an equal volume of sterile saline (2 ml/kg) instead of LPS. At the indicated time, brain tissues were gently harvested, immediately frozen in liquid nitrogen, and stored at −80°C. Brain tissue was postfixed in 4% paraformaldehyde overnight and processed routinely for paraffin embedding. All animals received care that complied with guidelines of the University of Tsukuba, and the university’s Graduate School of Medicine Animal Care and Use Committee approved the experimental procedure.
Hemodynamic Analysis.
On the day of the experiment, the rats were anesthetized with pentobarbital sodium (40 mg/kg ip), and a microtip pressure transducer catheter (SPC-320; Millar Instruments, Houston, TX) was inserted into the left carotid artery. Arterial blood pressure and heart rate were monitored and were recorded with a polygraph system (an AP-601G amplifier [Nihon Kohden, Tokyo, Japan], an AT-601G tachometer [Nihon Kohden], and a WT-687G thermal-pen recorder [Nihon Kohden]).
Quantitative Real-Time Polymerase Chain Reaction (PCR).
Total RNA from brain tissue specimens from the frontal cortex of SD rats was isolated by acid guanidinium thiocyanate-phenol-chloroform extraction with Isogen (Nippon Gene Ltd., Toyama, Japan) according to the manufacturer’s instructions. cDNA synthesis was performed as described elsewhere (15, 16).
The levels of preproET-1, ETA receptor, ETB receptor, and iNOS mRNA expression in the brain were analyzed by quantitative real-time PCR with TaqMan probe with an ABI Prism 7700 Sequence Detector (Perkin-Elmer Applied Biosystems, Foster, CA), as described elsewhere (15, 16). The gene-specific primers and TaqMan probes were designed using Primer Express software, version 1.5 (Perkin-Elmer), and were specific for preproET-1, ETA receptor, ETB receptor, iNOS, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA. The sequences of the oligonucleotides of preproET-1 have been published elsewhere (16). The primer sequences used for ETA receptor, ETB receptor, iNOS, and GAPDH mRNA in the present study were as follows: ETA receptor forward: 5′-GAATCTCTGCGCTCTCAGTGT-3′, ETA receptor reverse: 5′-GAGACAATTTCAATGGCGGTAATCA-3′, and ETA receptor probe: 5′-CAGGAAGCCACTGCTCT-3′; ETB receptor forward: 5′-GCTGGTGCCCTTCATACAGA-3′, ETB receptor reverse: 5′-CTTAGAGCACATAGACTCAACACTGT-3′, and ETB receptor probe: 5′-ATCCCCACAGAAGCCT-3′; iNOS forward: 5′-GTGGGTGGCCTCGAGTTC-3′, iNOS reverse: 5′-CCAATCTCGGTGCCCATGTAC-3′, and iNOS probe: 5′-CTGCCCCTTCAATGGTT-3′; and GAPDH forward: 5′-GTGCCAAAAGGGTCATCATCTC-3′, GAPDH reverse: 5′-GGTTCACACCCATCACAAACATG-3′, and GAPDH probe: 5′-TTCCGCTGATGCCCC-3′.
The expression of GAPDH mRNA was used as an internal control. Each PCR amplification was performed in triplicate, using the following profile: 1 cycle of 95°C for 10 mins, and 40 cycles of 94°C for 15 secs and 60°C for 1 min. For the standard curve in the real-time quantitative PCR, serial dilutions of a rat brain cDNA were performed using range of various concentrations (1×, 2×, 4×, 8×, and 16×). A water-based reaction mixture was prepared as a negative control.
Statistical Analysis.
Results are expressed as mean ± SD, and the sample number equals the number of animals in each group. Mean values were compared by means of one-factor analysis of variance, followed by the Scheffé’s test to account for the problem of multiple comparisons. Differences were considered to be statistically significant at a P value of <0.05.
Results
Hemodynamic Evaluation.
Systolic blood pressure decreased to 80 ± 7.6 mm Hg (P < 0.05 vs. controls) at 1 hr after LPS administration and gradually returned to normal (122 ± 8.4 mm Hg) after 10 hrs (Fig. 1A). Diastolic blood pressure also decreased to 56 ± 5.9 mm Hg (P < 0.05 vs. controls) at 1 hr after LPS administration and then returned to 108 ± 7.1 mm Hg (Fig. 1B). There were no significant changes in heart rate during sepsis (Fig. 1C).
Induction of Endotoxemia in the Frontal Cortex of the SD Rat Brain.
We confirmed the induction of endotoxemia in the brains of SD rats by measuring the expression of iNOS mRNA. The expression of iNOS mRNA in SD rat brain after LPS administration was 30-fold higher than that in the control rats (Fig. 2).
Expression of the Genes Encoding PreproET-1 and ET Receptors in the Frontal Cortex of the SD Rat Brain After LPS Administration.
The expression of preproET-1 mRNA was significantly increased (by approximately 2-fold) in the frontal cortex of SD rats at all time points after LPS administration, compared with the control rats (Fig. 3A). A time-dependent increase in the expression of the gene encoding ETA receptor was seen after LPS administration in the frontal cortex of SD rats, compared with the control rats (Fig. 3B). However, the expression of ETB receptor mRNA was at first upregulated and then gradually decreased as sepsis progressed (Fig. 3C).
Discussion
The present study demonstrates that the expression of the gene encoding preproET-1 in frontal cortex tissue is increased in LPS-treated SD rats at all time points, compared with expression in control rats. The expression of the gene encoding the ETA receptor is associated with the preproET-1 mRNA level in frontocortical tissue after LPS administration in SD rats, whereas the expression of the gene encoding the ETB receptor shows an initial upregulation followed by downregulation as the septic condition advances. iNOS, which is regarded as a marker of LPS-induced sepsis (17), is also remarkably upregulated in the brain tissue of SD rats after LPS administration. Both the systolic and diastolic blood pressure are significantly decreased early after administration of LPS but gradually normalized with the onset of endotoxemia.
A large number of inflammatory mediators and vasoactive substances are involved in the complex pathophysiology of sepsis and endotoxemia (17). Excessive NO from overexpression of iNOS causes vascular damage in endotoxemia and extravasation as an obligatory sequela to endotoxemia (18). In the present study, we also found a 30-fold increase in production of the gene encoding iNOS in the brain of SD rats after LPS administration. The detrimental characteristics of overexpression of iNOS have been associated with various organ abnormalities during LPS-induced endotoxemia. We speculate that the overexpression of iNOS may initiate several inflammatory changes in the brain of SD rats after LPS administration.
ET-1, a 21-amino-acid peptide, is a novel and long-lasting vasoconstrictor that is abundantly expressed in brain tissues (19). ET-1 activity has been demonstrated in 17 regions of the brain (19, 20). The rodent cerebral cortex has the highest levels of ET-1, with decreasing levels in the cerebellum, medulla, and pituitary gland (19). Thus, the present investigation was conducted using the frontal cortex of the brain of SD rats. Some studies demonstrated that ET-1 could not cross or alter the permeability of the rodent blood-brain barrier, implying that ETs are synthesized within the rodent brain (21). In the human cerebral cortex, immunoreactive ET-1 has been detected (20). After various types of brain injury, alterations in ET-1 synthesis occur (20). ET-1 concentration in cerebrospinal fluid is elevated in stroke patients, as well as in patients with subarachnoid hemorrhage (22). Similarly, ischemia or trauma in experimental animals results in an elevation of the ET-1 level in the central nervous system (23). Moreover, direct administration of ET-1 to brain blood vessels induces both vasoconstriction and cellular damage, whereas in vivo suppression of the gene encoding ET-1 has been reported to attenuate experimentally induced vasospasm in brain blood vessels (24). Thus, a clear role of ET-1 has been demonstrated in different cerebral pathological conditions.
In the present study, we found that expression of preproET-1 was increased in the frontal cortex of SD rats after LPS administration and that this upregulation was maintained at all time points. ET-1 is produced from prohormones known as preproET-1. In the septic brain, ET-1 may cause direct vasoconstriction, which impairs cerebrovascular circulation. Overexpression of ET-1 may cause brain damage by inducing the release of inflammatory cytokines after LPS administration. The regulative interaction between NO and ET-1 contributes to the maintenance of microvascular function in various organ systems. ET-1 may contribute to brain abnormalities during sepsis through enhanced NO synthesis. ET-1 may act in autocrine, paracrine, and endocrine fashions to stimulate cerebral NO synthesis through the induction of iNOS expression. Enhanced ET production in brain tissue in response to LPS injection can activate ET receptor and lead to an increase in intracellular cAMP synthesis and to activation of the protein kinase C pathway, both of which are known to cause transcriptional upregulation of iNOS expression (25). Moreover, induction of iNOS isoform by ET-1 in cultured endothelial cells and monocytes has recently been demonstrated (26).
ET-1 may also influence cerebral NO production and iNOS expression through the release of proinflammatory cytokines, such as TNF-α and IL-1. Moreover, superoxide produced by ET-1 may also participate in cerebral pathogenesis after LPS administration. Generally, ETA receptor causes vasoconstriction, and the upregulation of ETA receptor in the present study may play a role in the vasoconstricting effect of ET-1 in cerebral circulation after LPS administration to SD rats. On the other hand, the ETB receptor shows an initial upregulation followed by a downregulation in gene expression, which suggests that ETB receptor has a differential role in cerebral pathogenesis during sepsis that depends on the progression of endotoxemia. However, more studies should be performed to clarify the specific role of ET receptors in cerebral pathogenesis after LPS administration.
In the present study, preproET-1 was upregulated in frontal cortex tissue of SD rats after LPS administration. In addition, after LPS administration, expression of the genes encoding ET receptors was also significantly altered in the brains of SD rats. Expression of the gene encoding iNOS was also increased in brain tissue after LPS administration. Again, LPS caused a decrease in both systolic and diastolic blood pressure initially, and blood pressure tended to return to normal toward the end of LPS administration. Thus, there might be a loss of balance in the complex and tight interaction and regulation of ET-1 and iNOS in brain tissues in SD rats after LPS administration, and these alterations may play a role in cerebral pathogenesis in endotoxemia.
Changes in systolic blood pressure (A), diastolic blood pressure (B), and heart rate
(C) in SD rats after LPS administration and in control rats (n = 10).
Results are mean ± SEM. *P < 0.05, with respect to control. Changes in mRNA expression of iNOS in frontocortical tissue in SD rats after LPS
administration and in control rats. Expression of the iNOS gene in cerebral tissue was
determined by real-time PCR (n = 10). Results are expressed as
relative values (% of control). Results are mean ± SEM. *P < 0.01,
with respect to control. Changes in mRNA expression of preproET-1 (A), ETA receptor (B), and
ETB receptor (C) in the frontocortical tissue in SD rats after LPS
administration and in control rats. Levels of expression of preproET-1, ETA
receptor, and ETB receptor in cerebral tissue were determined by real-time
PCR (n = 10). Results are expressed as relative values (% of
control). Results are mean ± SEM. *P < 0.05, with respect to
control.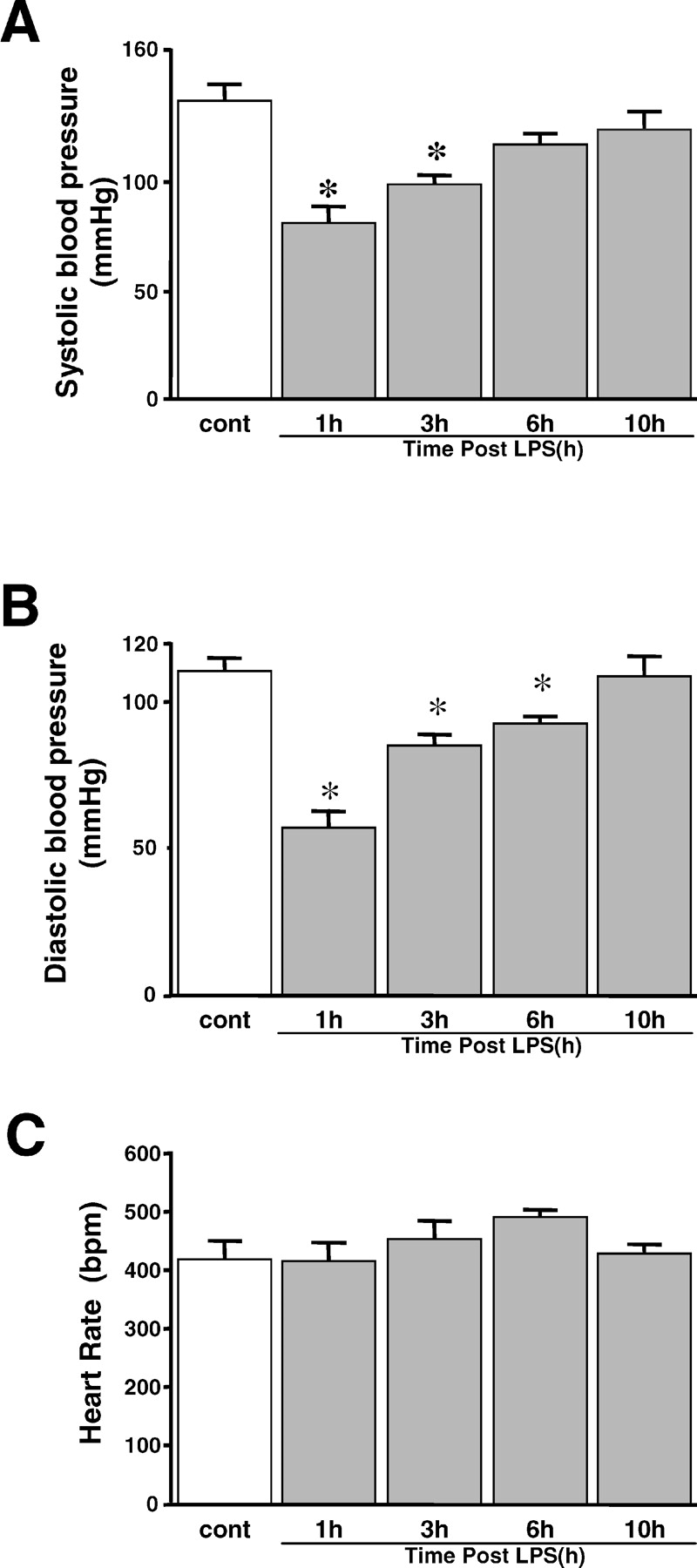


Footnotes
This work was supported by grants-in-aid for scientific research from the Ministry of Education, Science, Sports, and Culture of Japan (15390077 and 15650130) and by a grant from the Miyauchi project of Tsukuba Advanced Research Alliance at the University of Tsukuba.