
Abstract
The substantial role of endothelin-1 (ET-1) in the development of cerebral vasospasm (CVS) after subarachnoidal hemorrhage (SAH) has been demonstrated by numerous experimental and, recently, clinical investigations. Whether the expression or function of the ET(B) receptor is altered in CVS is still unclear, however. The aim of the present study was, therefore, to characterize the cerebroarterial ET(B) receptor function during CVS. Experimental CVS was induced by the rat double-hemorrhage model. Reduction of the cerebral blood flow (CBF) was confirmed by magnetic resonance perfusion-weighted imaging. Animals were sacrificed on days 3 (d3) and 5 (d5) after CVS induction. The basilar arteries (BA) were dissected, cut into ring segments, and prepared for measurement of isometric force in an organ bath. Concentration-effect curves (CECs) were constructed by cumulative application of ET-1, acetylcholine (Ach), or sarafotoxin S6c (S6c). Segments with (E+) endothelial function were used. CECs were compared by the maximum effect (Emax), the pD2, and the shift calculated on the pD2 level. The pD2 is the negative decadic logarithm of the concentration producing the half maximal effect (−log10EC50). After SAH, the relative regional CBF in the d3 and d5 groups was reduced to 63% and 32%, respectively, of the CBF in controls. ET-1 induced a dose-dependent contraction of segments with and segments without CVS. In E+ segments, the Emax for ET-1 was not significantly changed after SAH (mean values [ ± SEM] of 104% ± 4% for the control group, 106% ± 4% for the d3 group, and 104% ± 3% for the d5 group). The CECs, however, were significantly shifted to the left versus the control by factors of 2.4 in the d3 group and 3.6 in the d5 group. Relaxation by S6c was significantly reduced after SAH (Emax: 73% ± 11% in the control group, 21% ± 13% in the d3 group, and 13% ± 8% in the d5 group), whereas relaxation associated with Ach was not significantly changed (Emax: 45% ± 7% in the control group, 56% ± 6% in the d3 group, and 43% ± 6% in the d5 group). Significant contraction by S6c was not observed in E+ and E − segments in any of the study groups. The present data indicate the loss of the ET(B) receptor–mediated relaxation of the cerebral arteries in cases of CVS, which is independent of the endothelial nitric oxide synthase level.
Introduction
Cerebral vasospasm (CVS) is one of the substantial causes of delayed neurological deficits, which are responsible for the high morbidity and mortality among patients with aneurysmal subarachnoid hemorrhage (SAH) (1). Despite extensive experimental and clinical research during the last decades, the pathogenesis of CVS is not completely understood, and no specific therapeutic approach has been established. However, several observations support the concept that endothelin-1 (ET-1) is an important factor in the pathophysiological cascade leading to CVS. First, ET-1 has been shown to induce a potent and long-lasting constriction of cerebral vessels in vivo and in vitro, which continues even after removal of ET-1 (2–5). Second, the concentration of ET-1 in the cerebrospinal fluid of patients with SAH correlates with the development of CVS (6–8). Finally, CVS has been successfully treated with ET-receptor antagonists in several animal models (9–17) and, recently, in a clinical phase IIa study (18).
ET-1 acts in the cerebral vasculature through two specific receptors, ET(A) and ET(B) (19–21). Physiologically, vasoconstriction is induced by the ET(A)-receptor, which is expressed by smooth-muscle cells (2, 21). Activation of ET(B) receptors located on endothelial cells causes vasodilatation, which is mediated by the release of nitric oxide (22, 23).
Possible changes in the function of ET(B) receptors after SAH and during the development of CVS have not been sufficiently characterized. Increased expression of ET(B)-receptor mRNA (24) and upregulation of the cerebrovascular ET(B) receptors (25–27) were observed after SAH, which were postulated to mediate vasoconstriction. However, ET(B) receptor–dependent neuroprotective effects were also reported by animal models of cerebral ischemia (18, 28–31). Functional data characterizing the effect of the activation of the cerebrovascular ET(B) receptors under pathophysiological conditions like CVS are scarce in the literature. Therefore, the aim of the present investigation was to characterize the functional changes of the ET(B) receptor during the development of CVS after experimental SAH.
Material and Methods
Animal Procedures.
All procedures were in compliance with German Federal Guidelines for Animal Experiments and were performed with the permission of the local ethics committee. Twelve male Sprague-Dawley rats (body wt, 300–350 g) were included in the study. On Day 1 of the study, an experimental SAH was performed as described previously (32). The rats were anesthetized with midazolam (1 mg/kg body wt ip) and ketamine (100 mg/kg body wt ip) and fixed in a stereotaxic frame. After withdrawal of 0.1 ml of cerebrospinal fluid, 0.2 ml of autologous arterial blood was injected into the cisterna magna. The operation was repeated on Day 2. In sham-operated rats (n = 4), physiological saline was inserted instead of blood, without further changes to the procedure.
After the second SAH, animals were observed until Day 3 or Day 5. During this period, the animals received 5 ml of crystalloid solution with 0.0125 mg of fentanyl twice per ip. On Day 3 or Day 5, magnetic resonance perfusion-weighted imaging (MR PWI) was performed, and thereafter animals were killed by bleeding from the external carotid arteries under carbon dioxide anesthesia. The brain was removed and immersed in cooled modified Krebs-Högestett solution (119 mM NaCl, 3 mM KCl, 1.2 mM NaH2PO4, 1.5 mM CaCl2, 1.2 mM MgCl2, 15 mM NaHCO3, and 10 mM glucose) as described previously (33).
MR PWI.
MR examinations were performed at 3.0 T (Magnetom Trio; Siemens, Germany) with a wrist coil. PWI postprocessing was implemented with interactive software (Stroketoole; Digital Image Solutions, Germany) that included deconvolution algorithms, as described previously (32). The arterial input function was analyzed for semi-quantitative calculation of the relative regional blood flow in different regions of interest (ROIs). In each animal, ROIs were placed in the cerebrum and the masseter muscle. The ratio of the relative regional cerebral blood flow to the relative regional masseter muscle blood flow was used to determine the reduction of CBF due to CVS. For each group, a mean value of this ratio was calculated. CVS was quantified by this ratio (32).
Tissue Preparation.
The basilar artery was carefully dissected from the removed brain under microscopic control and was cut into four ringed segments. The segments were meticulously mounted on two L-shaped stainless steel rods (diameter, 70 μm) in an organ bath. Special care was taken to avoid any damage to the intimal surfaces.
In a part of the segments, the endothelium was mechanically removed using a human hair. Isometric force was measured, and the changes in isometric contraction were displayed by VitroDatWin (FMI GmbH), a computer program that is used to analyze biological signals. Organ baths were filled with modified Krebs-Högestett solution and were bubbled continuously with a humified gas mixture (95% O2 and 5% CO2), resulting in a pH of approximately 7.35.
After mounting of the basilar artery ring segments, the organ bath temperature was increased gradually to 37°C. After an adaptation period of 60 mins, the segments were stretched repeatedly until a stable level of resting tension between 3 to 4 mN was reached.
A reference contraction (124 mM K+(A)) was elicited by exchanging the bath solution with 124 mM K+-Krebs solution (NaCl was replaced with KCl). At the end of the experiments, this procedure was repeated for each segment (124 mM K+ (E)). If a segment developed a force of < 2.5 mN or reached < 85% of the first contraction at the end of the experiment, it was excluded from assessment. The endothelial function was tested by application of acetylcholine (Ach; 1 × 10−5 M) after precontraction with 5-hydroxytryptamine (5-HT; 1 × 10−5 M).
Contraction force was measured in mN and expressed as a percentage of the 124-mM K+(A) reference contraction. Concentration-effect curves (CECs) for ET-1 (1 × 10−12 to 3 × 10−7) and Sarafotoxin S6c (S6c; 1 × 10−12 to 3 × 10−8) were performed by cumulative application. To investigate the relaxation due to S6c, the segments were precontracted with prostaglandin F2α (PGF2α; 10 −5 M) (incubation period, 30 mins) before S6c was added cumulatively.
Compounds and Solvents.
ET-1 and S6c were purchased from Calbiochem-Novabiochem (Bad Soden, Germany). Ach, 5-HT, and PGF2α were purchased from Sigma-Aldrich (Schnelldorf, Germany). All compounds were freshly dissolved in distilled water on the day of the experiment.
Data Analysis.
Results are expressed as mean ± SEM. For each CEC, the maximum contraction (Emax) and pD2 (−log10EC50) or EC50 (i.e., the concentration at which half of the maximal effect occurs) were calculated. The pD2 was calculated by linear regression analysis of the CEC, after logarithmic transformation of the concentrations greater than and less than the EC50. Thus, the higher the pD2, the smaller the concentration that is needed to reach the half-maximal effect. Relaxation was calculated as the percentage decrease from precontraction (i.e., the force induced by 5-HT for ACh or by PGF2α for S6c) and was determined using the lowest peak of vessel tone after application of each compound over a 20-min observation period (34).
Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey’s test for post hoc comparisons of mean values. Statistical significance was defined as a P value of < 0.05.
Results
Development of CVS was proven by MR PWI. According to MR PWI, the ratio of relative regional cerebral blood flow and blood flow of the masticatory muscles after SAH was reduced to 63% ± 9% in the d3 group and to 32% ± 11% in the d5 group, compared with control animals. This indicates a CVS with a significant reduction of CBF on Day 5.
Segments under resting tension were analyzed in an organ bath. Cumulative application of ET-1 in both segments, obtained from controls and animals after SAH, induced a dose-dependent contraction of rat basilar artery ring segments. A leftward shift was observed in the SAH groups. However, the Emax did not change significantly (Table 1). The shift observed for the Day 5 group seemed to be higher than that for the Day 3 group (Fig. 1 and Table 1).
Cumulative application of S6c did not induce a significant relaxation in segments from the Day 3 and Day 5 groups that were precontracted with PGF2α. In contrast, relaxation was observed in controls (Fig. 2a). The Ach-induced relaxation, however, was not significantly impaired after SAH, indicating a functionally intact endothelium (Fig. 2b).
Discussion
The aim of this study was to characterize changes in ET(B) receptor in cerebral vessels during the development of CVS after experimental SAH. CVS was confirmed in the present study by the quotient of CBF/masseter muscle blood flow, which showed a good correlation with the development of angiographic CVS in a previous study (32). This suggests that the ring segments from animals after SAH in this study had developed alterations, which are characteristic of CVS.
CECs for ET-1 were shifted to the left without changing the Emax, which indicates a higher sensitivity of cerebrovasculature to ET-1 (Table 1 and Fig. 1). This observation could be explained by an expression of a constricting ET(B) receptor under pathological conditions, which was postulated previously (25–27). However, an increase of the maximum contraction would also be expected under these conditions. Furthermore, the application of S6c on segments under resting tension did not induce any contraction of ring segments, neither in controls nor in animals after SAH (data not shown). Therefore, it seems to be unlikely that the higher sensitivity is mediated by the smooth-muscle ET(B2) receptor subtype.
Another possible explanation for a leftward shift in the present results could be the loss of the ET(B) receptor–dependent relaxation. The loss of this functional antagonism to the ET(A)-mediated contraction results in a leftward shift of the CEC (35). Accordingly, the pD2-values for ring segments from animals with CVS are comparable with results from BA ring segments without endothelial function in previous investigations (29, 35). The decrease of the vasorelaxation by selective ET(B) receptor activation using S6c further supports the concept of impaired endothelial ET(B) receptor–dependent vasodilatation in cases of CVS (Fig. 2A). However, the endothelium was not harmed and the nitric oxide–dependent pathway was not impaired by CVS in the present investigation (Fig. 2B and Table 1).
In summary, our data suggest a selective decrease of endothelial ET(B) receptor–dependent relaxation during the development of CVS. After SAH, increased ET-1 levels in cerebrospinal fluid are known. Additionally, the impairment of the ET(B)-receptor relaxation may be a further pathophysiologic mechanism in the development of CVS.
Time Course of the ET-1-Induced Contraction, the ET(B) Receptor–Dependent Relaxation, and the Endothelium-Dependent Relaxation After Experimental SAH. a
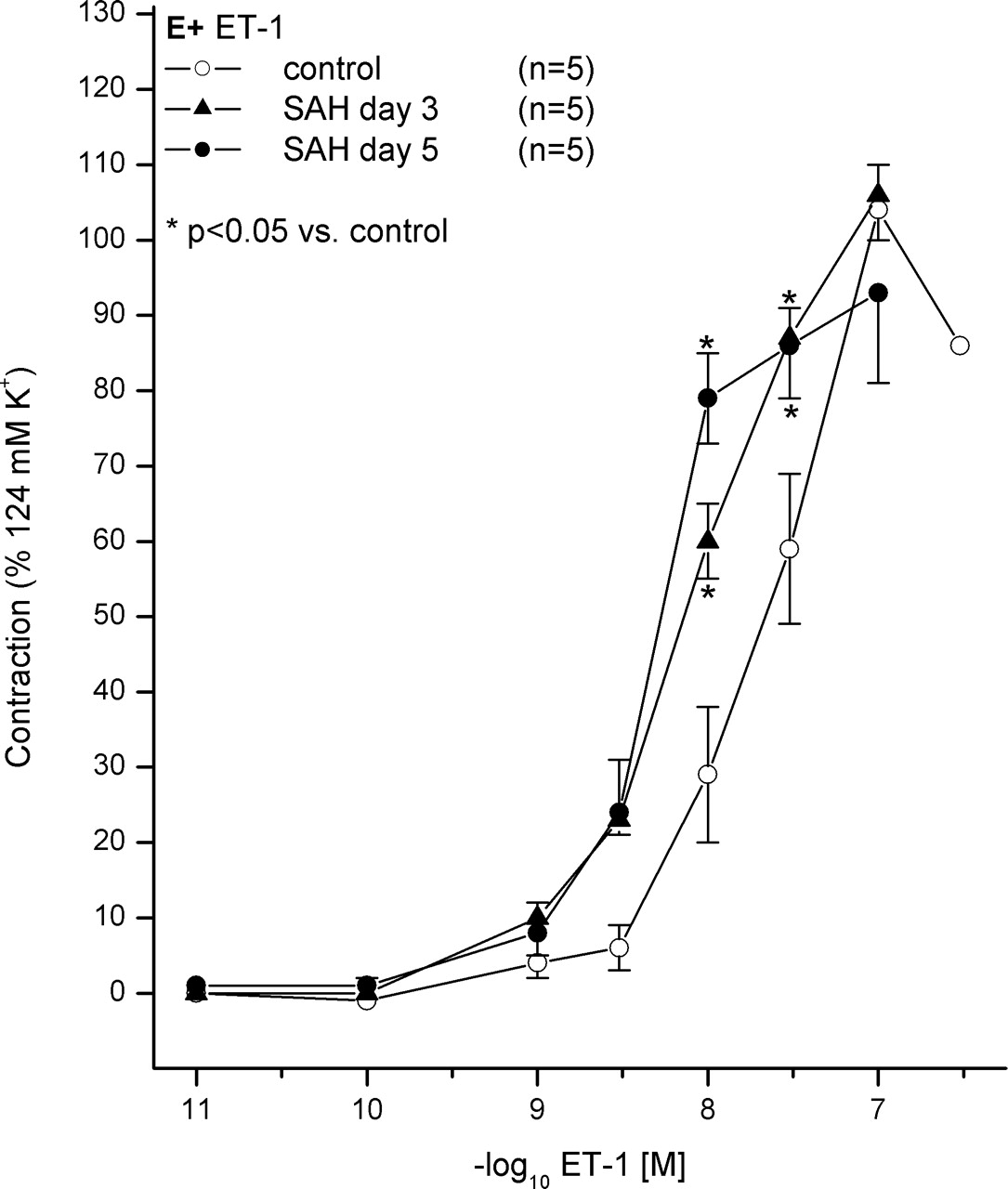
Contraction effect curve (CEC) for ET-1. Contraction of ET-1 is shown for controls and animals after SAH on Day 3 and Day 5. The CECs in SAH groups are shifted to the left dose dependently without reduction of maximum contraction, indicating an increased sensitivity of vessels during the development of CVS. Data mean ± SEM. *P < 0.05 vs. ET-1 control E+.
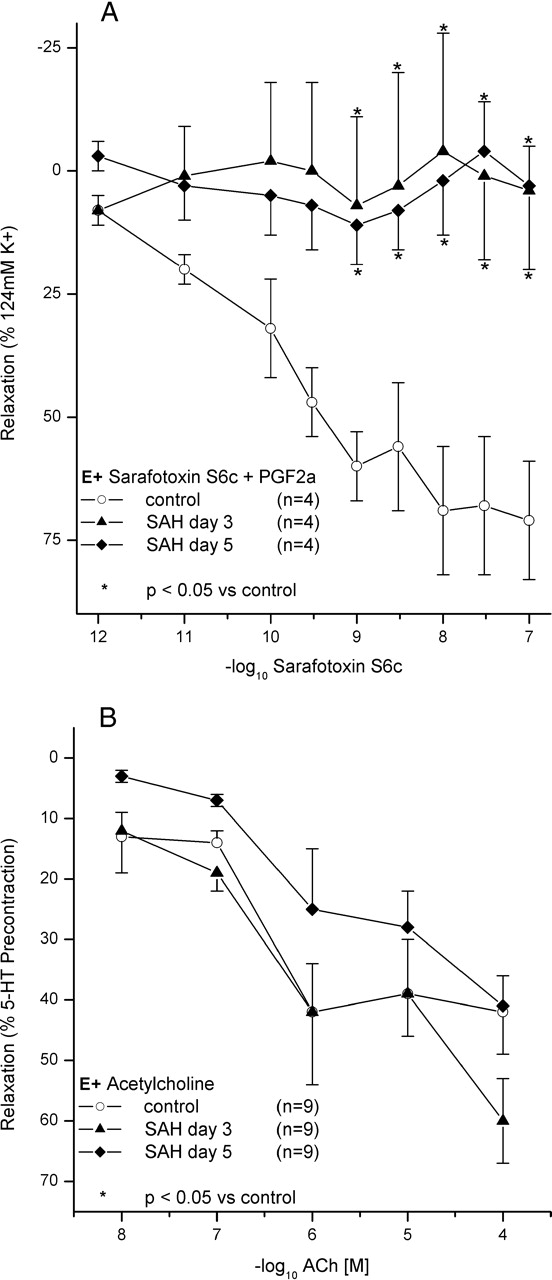
(A) CEC for S6c after precontraction by PGF2α. Relaxation by S6c is shown for controls and animals after SAH on Day 3 and Day 5. The relaxation is significantly reduced in both SAH groups. Data indicate a reduction of the ET(B) receptor–dependent relaxation. Data are mean ± SEM. *P < 0.05 vs. the control group. (B) CEC for Ach after precontraction by 5-HT. Relaxation by ACh is shown for controls and animals on Day 3 and Day 5 after SAH. Relaxation is not significantly impaired after SAH, indicating a functionally intact endothelium. Data are mean ± SEM. *P < 0.05 vs. ACh.
Footnotes
The present investigation was funded exclusively by the Department of Neurosurgery (Johann Wolfgang Goethe University-Hospital, Frankfurt/Main, Germany) and by the Adolf-Messer-Stiftung (Königstein/Taunus, Germany).
Acknowledgements
We thank M. Eberhardt for excellent technical assistance and Dr. A. Theissen for experimental support and animal care.