
Abstract
The green tea polyphenol, epigallocatechin-3-gallate (EGCG), has been shown to prevent cancer; however, a precise mechanism responsible for tumor growth inhibition has not yet been clearly described. The endothelin (ET) A receptor (ETAR)/ET-1 autocrine pathway is overexpressed in ovarian carcinoma and triggers tumor growth, neoangiogenesis, and invasion. These latter tumor-promoting effects are mediated through the activation of cyclooxygenase (COX)-1– and COX-2–dependent pathways by ET-1. In the present study, pretreatment of HEY and OVCA 433 ovarian carcinoma cell lines with green tea and EGCG inhibited ET-1/ETAR expression, ETAR-mediated COX-1/2 mRNA expression, and COX-2 promoter activity. These effects were associated with a significant reduction in the COX-1/2–derived prostaglandin E2 (PGE2) production. These results provide a novel insight into the mechanism by which EGCG, by affecting ETAR-dependent COX-1/2 pathways may inhibit ovarian tumors suggesting that EGCG may be useful in preventing and treating ovarian carcinoma in which activation of ETAR by ET-1 plays a critical role in tumor growth and progression.
Introduction
Ovarian cancer represents the leading cause of death among gynecologic malignancies. Approximately 26,250 women are diagnosed yearly, with an overall 5-year survival rate of only 47%. Despite recent advances in surgery and chemotherapy, improvement in long-term survival of these patients has been slight (1). Development of new treatment options strongly relies on improved knowledge of the molecular mechanisms underlying ovarian carcinoma initiation and progression. Primary and metastatic ovarian carcinoma express high levels of endothelin (ET)-1 and ETA receptor (ETAR) compared with normal ovarian tissue (2). Engagement of ETAR by ET-1 promotes tumor cell proliferation (2, 3), apoptosis protection (4), invasiveness (5), and neovascularization by stimulating vascular endothelial growth factor (VEGF) secretion (6–9). The proangiogenic and invasive properties of ET-1 are amplified by the capacity of ET-1 to induce cyclooxygenase (COX)-1 and COX-2 enzymes and prostaglandin (PG) E2 production (10, 11). The COX-1 and COX-2 enzymes catalyze the rate-limiting step in the conversion of arachidonic acid to PGs (10), and are involved in tumor progression. We found that ET-1–induced PGE2 participates in these complex mechanisms by stimulating VEGF secretion cell migration and matrix metalloproteinase (MMP) activation. ETAR blockade by the selective receptor antagonist, ABT-627, inhibits the growth of ovarian carcinoma xenografts concomitantly with a reduction of microvessel density, MMP-2, VEGF, and COX-2 expression, indicating that the blocking of the ET-1/ETAR axis may represent a potential therapeutic target in ovarian carcinoma management (11, 12).
Several studies on human and animal models have demonstrated that green tea may be effective in preventing and treating cancer of the breast, prostate, esophagus, pancreas, and colon. The anticancer property of green tea polyphenol, (–)-epigallocatechin gallate (EGCG), is likely the result of blocking biochemical pathways involved in ovarian tumor growth (13).
Understanding the molecular mechanism of green tea polyphenol–mediated inhibition of ovarian tumor growth is essential in devising preventive and therapeutic approaches. In the present study, we show that green tea polyphenol induces inhibition of both ET-1 and ETAR expression, which, in turn, leads to the blockade of the COX-1/2–mediated signaling cascade. These results provide evidence that the ET-1/ETAR pathway is a critical target for green tea and EGCG in ovarian cancer growth and identify a potential role of green tea polyphenol in the prevention and treatment of this malignancy.
Materials and Methods
Cells and Cell Culture Conditions.
Human ovarian carcinoma cell lines, HEY and OVCA 433, previously characterized for ET-1 receptor expression and for ET-1 production (2, 6), were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal calf serum (FCS) and 1% penicillin and streptomycin at 37°C in 5% CO2/95% air. The cells were serum-starved by incubation for 24 hrs in serum-free DMEM. All culture reagents were from Invitrogen (Paisley, Scotland, UK). ET-1 (Peninsula Laboratories, Belmont, CA), was used at 100 nM and incubated with the cells for the indicated times. Green tea infusion was prepared from commercially available (China or Sri Lanka) green tea leaves steeped in 1 liter (12.4 g/l) of boiled distilled water for 1–2 mins. Pretreatment of cells with 1 μM BQ 123 (Peninsula Laboratories), or with the indicated concentrations of EGCG (Sigma Chemical Co., St. Louis, MO) or green tea infusion was performed for 30 mins before the addition of ET-1.
Reverse Transcription Polymerase Chain Reaction (RT-PCR).
Total RNA from HEY and OVCA 433 cells was extracted using TRIzol (Invitrogen). RT-PCR was performed using the AccessQuick RT-PCR System (Promega, Madison, WI) according to the manufacturer’s instructions. The primer sets were as follows: ET-1, 5′-TGCTCCTGCTCGTCCCTGATGGATAAAGAG-3′ and 5′-GGTCACATAACGCTCTCTGGAGGGCTT-3′; ETAR, 5′-CACTGGTTGGATGTGTAATC-3′ and 5′-GGAGATCAATGACCACATAG-3′; COX-1, 5′-TGCCCAGCTCCTGGCCCGCCGCTT-3′ and 5′-GTGCATCAACACAGGCGCCTCTTC-3′; COX-2, 5′-TTCAAATGAGATTGTGGGAAAATTGCT-3′ and 5′-AGATCATCTCTGCCTGAGTATCTT-3′; GAPDH, 5′-ACCACAGTCCATGCCATCAC-3′ and 5′-TCCAC-CACCCTGTTGCTGTA-3′. Thirty-five cycles of amplification were performed under the following conditions: melting at 95°C for 30 secs; annealing at 58°C for 30 secs; and extension at 72°C for 60 secs. The PCR products were analyzed by electrophoresis on a 2% agarose gel, and the relative intensities of signals were quantified using NIH Image (Scion Corporation, Frederick, MD).
Enzyme-linked Immunosorbent Assay (ELI-SA).
Subconfluent HEY cells were serum-starved for 24 hrs and incubated for the indicated times. The conditioned media were collected, centrifuged, and stored in aliquots at −20°C. Levels of PGE2 released into the cells medium were measured by the PGE2-Correlate-EIA (Assay Designs, Inc., Ann Arbor, MI) according to the manufacturer’s instructions. The sensitivity of the assay is 13.4 pg/ml for PGE2. Intraassay variation is 5.8% for PGE2 and interassay variation is 5.1% for PGE2.
Transfection of Reporter Construct and Luciferase Assay.
Reporter construct phPES2 (−1432/+59) containing the 5′-flanking region of the human COX-2 gene (11) were kindly provided by Dr. DuBois (Vanderbilt University Medical Center, Nashville, TN). For transient transfection, 1 × 105 cells were plated in 6-well plates 48 hrs before transfection. The cells were cotransfected with 0.5 μg of COX-2 luciferase plasmid construct and with 0.05 μg of the pCMV–β-galactosidase plasmid (Promega), using LipofectAMINE reagent (Invitrogen), as described in the manufacturer’s protocol. The cells were lysed and their luciferase activities were measured (Luciferase assay system; Promega). The results were normalized to β-galactosidase activity.
Statistical Analysis.
Results are representative of at least three independent experiments, each performed in triplicate. All statistical analysis was assessed using a two-tailed Student’s t test.
Results
Green Tea and EGCG Inhibit ETAR and ET-1 Expression.
Sustained ETAR signaling caused by an autocrine ET-1/ETAR loop has been implicated in ovarian tumor growth and progression (2, 6). To evaluate the effect of green tea polyphenols on the ET-1/ETAR axis, we examined ETAR and ET-1 expression in ovarian carcinoma cells, which secrete high levels of ET-1, in the range required for activation of ETAR in an autocrine fashion. HEY and OVCA 433 cells were treated with different concentrations of green tea and of the polyphenol EGCG, the principal mediator of the green tea antitumor effect. Total RNA or conditioned media were collected after 6 hrs and 24 hrs, respectively. RT-PCR and ELISA demonstrated that green tea and EGCG induced a dose-dependent inhibition of ETAR and ET-1 expression at the mRNA (Fig. 1A) and protein (Fig. 1B) levels. These results demonstrate that green tea and the polyphenol EGCG are able to reduce the expression of the ET-1/ETAR axis in ovarian carcinoma cells.
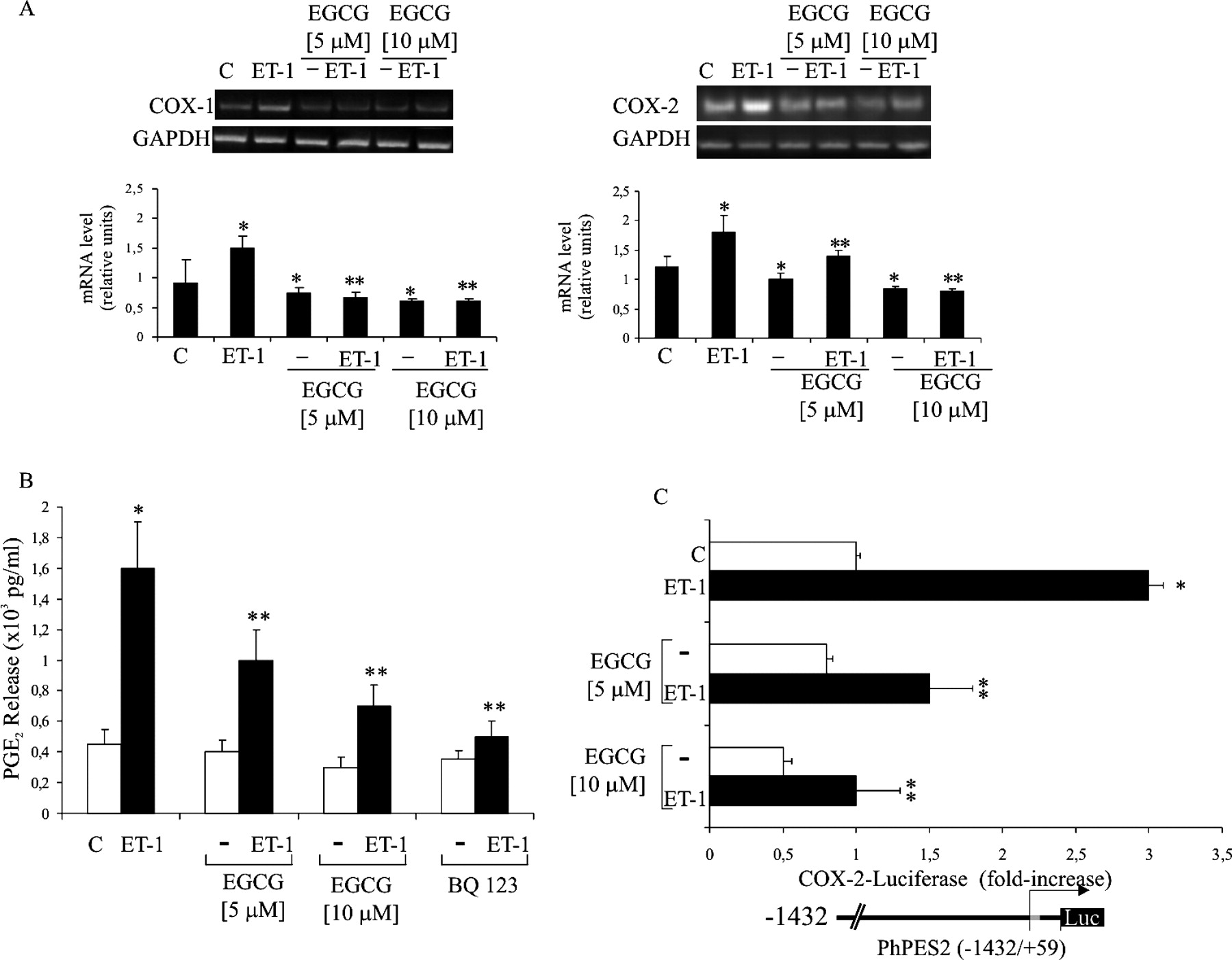
EGCG Inhibits ET-1–Induced COX-1 and COX-2 Expression and PGE2 Production.
Because in HEY and OVCA 433 cells, ET-1 induces expression of both COX-1 and COX-2, which contribute to PGE2 and VEGF production and cell invasion (10, 11), we examined the ability of EGCG to modulate the basal and the ET-1–induced COX-1 and COX-2 expressions. We, therefore, treated HEY cells for 6 hrs with different concentrations of EGCG alone or in combination with ET-1 and analyzed COX-1 and COX-2 mRNA expression by RT-PCR. Treatment with EGCG resulted in a dose-dependent reduction of the basal and ET-1–induced COX-1/2 levels (Fig. 2A). Moreover, to evaluate whether the reduction of COX-1 and COX-2 expression results in an inhibition of their enzymatic activity, we measured the basal and the ET-1–induced PGE2 production after treatment with EGCG. HEY cells were treated with different concentrations of EGCG and with the selective ETAR antagonist, BQ 123, alone or in combination with ET-1. As shown in Figure 2B, EGCG treatment resulted in a dose-dependent reduction of the basal and the ET-1–induced PGE2 production, comparable to that induced by BQ 123. These results demonstrate that EGCG is able to reduce the ETAR-mediated COX-2–derived PGE2 production in ovarian carcinoma cells.
EGCG Inhibits ET-1–Induced COX-2 Promoter Activity.
ET-1 controls COX-2 expression at transcriptional and posttranscriptional levels (11). To determine whether EGCG regulates the basal and the ET-1–induced COX-2 promoter activity, HEY cells transfected with the human COX-2 promoter (phPES2–1432/+59), were pre-treated with different concentrations of EGCG alone or in combination with ET-1. After 24 hrs, luciferase activity was measured and normalized to β-galactosidase activity. As shown in Figure 2C, ET-1 treatment induced a 3.1-fold increase in luciferase activity that was significantly reduced by EGCG in a dose-dependent manner. Moreover, EGCG was able to reduce the basal level of COX-2 promoter activity. These results indicate that EGCG inhibits the ET-1–mediated regulation of the COX-2 promoter in ovarian cancer cells.
Discussion
Epidemiologic and clinical studies have clearly shown that the COX-1 and COX-2 enzymes play a key role in the progression of ovarian carcinoma (12, 14). In these malignancies, COX-1– and COX-2–derived PGE2 have been implicated in tumor angiogenesis (11, 14), and in the invasive and migratory capacity (10). We have previously demonstrated that in vivo blockade of ETAR autocrine pathway using a highly specific ETAR antagonist results in tumor growth reduction associated with a significant inhibition of microvessel density, expression of VEGF, MMP-2, and COX-2, and increased tumor apoptosis (11, 12), suggesting that the ETAR signaling pathway may represent a potential preventive and therapeutic tumor target (15). The limited treatment options of ovarian cancer have prompted the need for developing alternative strategies for the management of this disease. Because tumor chemo-prevention and therapy with green tea or green tea polyphenols seem to offer new approaches to block tumor growth and progression, the study of green tea and EGCG mechanisms of action is becoming essential from the biologic as well as from the therapeutic point of view. Epidemiologic studies have shown that green tea consumption can enhance the survival of patients with epithelial ovarian cancer (16), and EGCG has been reported to have a growth inhibitory effect on ovarian tumor cell lines (17). In this study, we addressed whether ET-1/ETAR–induced autocrine signaling pathways are modulated by green tea polyphenols, and whether this treatment inhibits the expression of molecules involved in angiogenesis and invasiveness. There is a growing body of compelling evidence that targeted inhibition of abnormal or improper overexpression or activity of COX-2 and COX-1 is valuable for preventing ovarian cancer. In this study, we analyzed the capacity of green tea and EGCG to regulate the ET-1 axis and the ET-1–mediated COX-2/COX-1 pathways. We demonstrated that in ovarian carcinoma cells, treatment with green tea infusion or with the polyphenol EGCG significantly inhibited the expression of ET-1 and ETAR mRNA expression. These results are in agreement with several studies showing that the inhibition of growth factor receptor expression and pathways is one of the possible mechanisms of actions of green tea and EGCG (18). Concordant with these reports, we found that green tea and EGCG inhibited the ET-1–induced COX-1 and COX-2 expression that resulted in a reduction of PGE2 secretion. These results are, therefore, of biologic and clinical relevance for understanding the molecular mechanism underlying the antitumor activity of green tea polyphenol, which may be useful in developing preventive as well as therapeutic protocols.
Because high levels of ET-1 are associated with a number of malignancies (15), the impairment of ETAR signaling by green tea could be of broad therapeutic relevance.
Green tea and EGCG inhibit ETAR and ET-1 expression. (A) Serum-starved HEY
cells were treated with the indicated concentrations of green tea or EGCG for 6 hrs
and total RNA was extracted. ETAR and ET-1 expression was evaluated by
RT-PCR. Primers for the amplification of the GAPDH gene were used as
control for the RT-PCR experiments. Densitometric analysis of RT-PCR products was
evaluated by Scion image. Data were statistically analyzed and represented as the
average value of three independent experiments. Bars, ± SD. *P <
0.006; **P < 0.005, compared with the control. (B) HEY and OVCA
433 cells were treated with the indicated concentrations of green tea or EGCG for 24
hrs and conditioned media were collected. ET-1 secretion was measured by ELISA.
Results were representative of at least three independent experiments, each performed
in triplicate. Bars, ± SD. *P < 0.005; **P <
0.001, compared with the control. EGCG inhibits ET-1–induced COX-1 and COX-2 expression. Serum-starved HEY cells were
treated with different concentrations of EGCG alone or in combination with 100
nM ET-1 for 6 hrs or 24 hrs and total RNA (A) or conditioned media
(B) were collected. COX-1 and COX-2 mRNA were
analyzed by RT-PCR by using specific primers for COX-1 and
COX-2. Primers for the amplification of the GAPDH
gene were used as controls. (A) Densitometric analysis of RT-PCR products was
evaluated by Scion image. Data were statistically analyzed and represented as the
average value of three independent experiments. Bars, ± SD. *P <
0.006 compared with the control; **P < 0.005 compared with ET-1.
(B) PGE2 production was measured in conditioned media by ELISA. Bars, ± SD;
*P < 0.001, compared with control; **P <
0.005, compared with ET-1. (C) HEY cells were transfected with the phPES2 (−1432/+59)
construct of the human COX-2 promoter coupled with a luciferase reporter gene, as
indicated in the bottom of the figure, and incubated for 12 hrs with different
concentrations of EGCG alone or in combination with 100 nM ET-1.
COX-2 promoter activity was examined using the Luciferase Assay System in four
independent experiments and was expressed as fold-increase relative to control.
*P < 0.006 compared with control; **P <
0.004 compared with ET-1.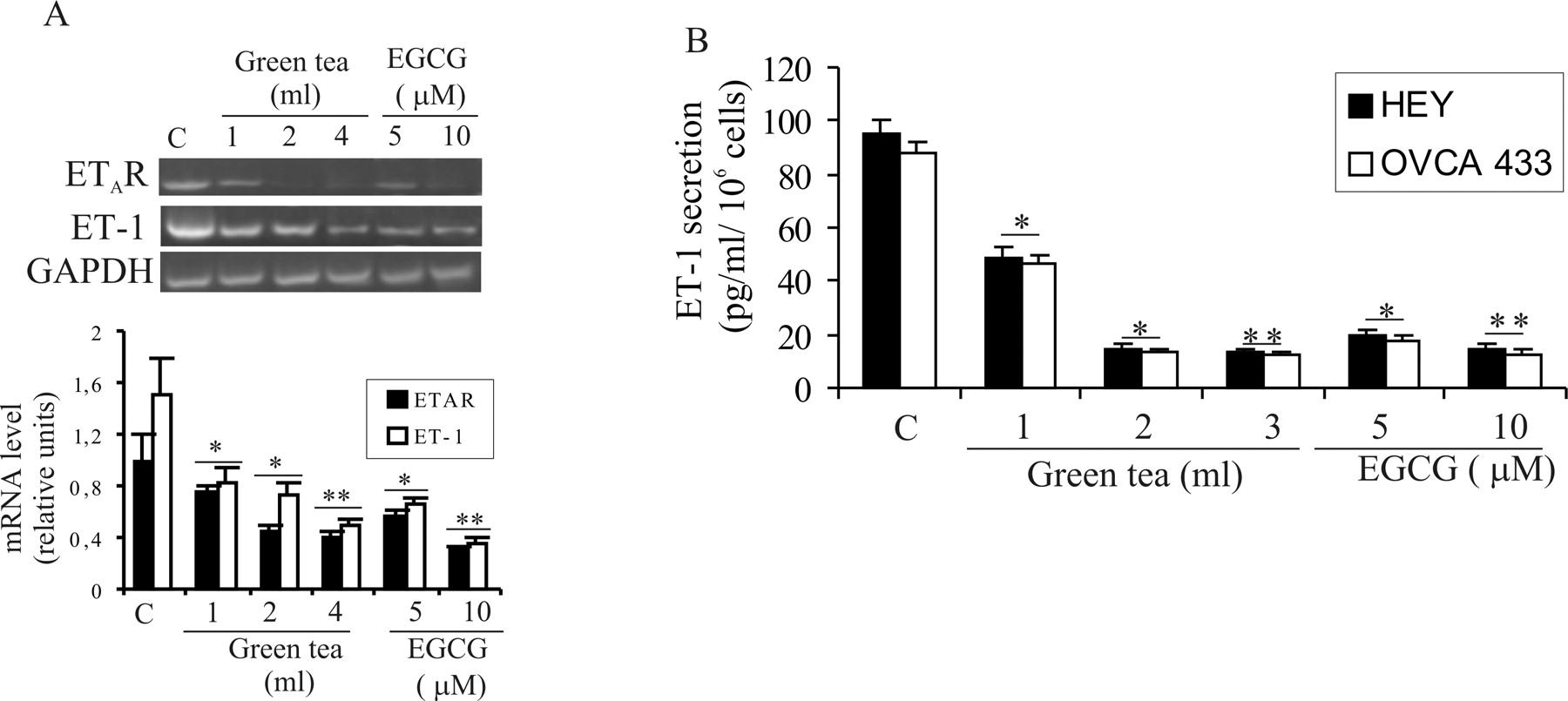
Footnotes
This work was supported by grants from the Associazione Italiana Ricerca sul Cancro, Ministero della Salute, and Ministero, Istruzione, Università, Ricera-Consiglio Nazionale delle Ricerche.
Acknowledgements
We thank M. V. Sarcone for secretarial support.