
Abstract
Endothelin-1 (ET-1) is present at high concentrations in ovarian cancer ascites and is overexpressed in primary and metastatic ovarian carcinomas. In these tumors, the presence of ET-1 correlates with tumor grade, enhanced neovascularization, and with vascular endothelial growth factor (VEGF) expression. ET-1 acts as an autocrine factor selectively through ETA receptor (ETAR), predominantly expressed in ovarian carcinoma cells resulting in increased VEGF production and VEGF-mediated angiogenic effects. Previous results demonstrated that in ovarian carcinoma cells, activation of the ET-1/ETAR axis promotes cell proliferation, neovascularization, and invasion, which are the principal hallmarks of tumor progression. The present study was designed to investigate the in vitro effects of trans, trans-2(4-methoxydhenyl)-4-(1–3-benzodiazol-5-yl)-1-(dibutylaminocarbonylmethyl)-pyrrolidine-3-carboxylic acid (ZD4054), an orally active specific ETAR antagonist, on the ET-1–induced mitogenic effect in OVCA 433 and HEY ovarian carcinoma cell lines secreting ET-1 and expressing ETAR and ETBR mRNA. We show that ETAR blockade by ZD4054 inhibits ET-1–induced mitogenic effects, while the ETBR antagonist, BQ 788, is ineffective. In conclusion, our data demonstrate that ZD4054 is capable in inhibiting the proliferative activity of ET-1, indicating that this specific ETAR antagonist may be a potential candidate in developing novel treatment of ovarian carcinoma.
Introduction
Ovarian cancer, the leading cause of death from gynecologic malignancy, is a highly metastatic disease characterized by widespread peritoneal dissemination and ascites at the time of initial diagnosis in most patients (1).
Endothelin-1 (ET-1) is a peptide produced primarily in vascular cells and in elevated amounts by a variety of cancer cells (2). ET-1 acts through two distinct subtypes of G-protein–coupled receptors, namely ETA and ETB. ETA receptor (ETAR) binds selectively to ET-1, whereas ETBR binds both ET-1 and ET-3. ET-1 has been implicated in the pathophysiology of a wide range of human tumors (3), including ovarian carcinoma (4, 5). We have previously demonstrated that ET-1 is overexpressed in primary and metastatic ovarian carcinomas compared with normal ovarian tissues (5). Moreover, ET-1 is present at high levels in ovarian tumor effusions, playing an important role in the progression of this disease (6). In ovarian tumor cells, ET-1 acts as an autocrine growth, survival, and angiogenic factor selectively through the ETAR (4, 6–9), activating different signaling pathways (10). The pathways include mitogen-activated protein kinase (MAPK); phosphoinositide 3-kinase (PI3K)–dependent Akt activation; epidermal growth factor receptor (EGFR) transactivation, which is partly responsible for MAPK phosphorylation (11); and p125 focal adhesion kinase and paxillin activation, which are thought to transduce signals involved in tumor cell invasion (10, 12).
We also demonstrated that, in addition to controlling growth and survival of primary tumor, ET-1/ETAR is a potential target to prevent the spread and survival of tumor metastasis. ET-1 in fact induces overexpression and activation of a panel of proteinases known to be associated with highly invasive and metastatic ovarian carcinoma with a concomitant marked increase in ovarian tumor cell migratory and invasive activities. The enhanced proteolytic capability of ovarian carcinoma cells induced by ET-1 indicates a biochemical mechanism by which disruption of local tissue architecture (to allow tumor growth and spread of ovarian carcinoma) may be mediated by ET-1 through ETAR binding. In view of these findings, the ETAR has been proposed as a potential target for anticancer therapy (14). The recent identification of small molecules that inhibit ligand-induced activation of the ETAR now offers the possibility of testing this therapeutic approach in a clinical setting. Importantly, pharmacologic blockade of ETAR inhibits tumor xenograft growth and the molecular determinants involved in tumor progression (15).
Among various ETAR inhibitors, trans, trans-2(4-methoxydhenyl)-4-(1–3-benzodiazol-5-yl)-1-(dibutylaminocarbonylmethyl)-pyrrolidine-3-carboxylic acid (ZD4054) is an orally active ETAR antagonist in early clinical development for the treatment of cancer. The synthesis and molecular characterization of ZD4054, a nonpeptide ETAR antagonist, have been described previously (16). ZD4054 binds to ETAR with high affinity and has no detectable affinity for ETBR. In preclinical studies, ZD4054 selectively inhibits ETA-mediated antiapoptotic effects, but not ETB-mediated proapoptotic effects, in human smooth muscle cells (17); blocks ETA-mediated activation of p44/42 MAPK in murine osteoblast cells; and inhibits ET-1–induced proliferation of human immature pre-osteoblast cells (17). Because activation of ETAR is involved in the proliferation, migration, and invasion of ovarian carcinoma cells, we tested whether the ETAR antagonist ZD4054 may inhibit ovarian carcinoma cell growth in vitro.
Materials and Methods
Cells and Cell Culture Conditions.
Human ovarian carcinoma cell lines HEY and OVCA 433, previously characterized for ET-1 receptor expression and for ET-1 production (2, 6), were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal calf serum and 1% penicillin-streptomycin at 37°C under 5% CO2 to 95% air. The cells were serum-starved by incubation for 24 hrs in serum-free DMEM. All culture reagents were from Invitrogen (Paisley, Scotland). ET-1 (Peninsula Laboratories, Belmont, CA) was used at 100 nM and incubated with the cells for the indicated times. Pretreatment of cells with 1 μM ZD4054 (AstraZeneca, Macclesfield, UK) was performed for 30 mins before the addition of ET-1.
Reverse Transcription–Polymerase Chain Reaction (RT-PCR).
Total RNA from HEY and OVCA 433 cells was extracted using TRIzol (Invitrogen). RT-PCR was performed using AccessQuick RT-PCR System (Promega, Madison, WI), according to the manufacturer’s instructions. The primers sets were as follows: ET-1, 5′-TGCTCCTGC-TCGTCCCTGATGGATAAAGAG-3′ and 5′-GGTCACATAACGCTCTCTGGAGGGCTT-3′; ETAR, 5′-CACTGGTTGGATGTGTAATC-3′ and 5′-GGAGATCAATGACCACATAG-3′; ETBR, 5′-TCAACACGGTTGTGTCCTGC-3′ and 5′-ACTGAATAGCCACCAATCTT-3′; and GAPDH, 5′-ACCACAGTCCATGCCATCAC-3′ and 5′-TCCACCACCCTGTTGCTGTA-3′. Thirty-five cycles of amplification were performed under the following conditions: melting at 95°C for 30 sec; annealing at 58°C for 30 sec; and extension at 72°C for 60 sec. The PCR products were analyzed by electrophoresis on a 2% agarose gel.
Enzyme-Linked Immunosorbent Assay (ELISA).
Subconfluent HEY cells were serum-starved for 24 hrs and incubated for the indicated times. ET-1 release was measured in triplicate on microtiter plates using an ELISA kit (R&D Systems, Minneapolis, MN), according to the manufacturer’s instructions. ET-1 may be measured in the range of 0–120 pg/ml. The sensitivity is less than 1.0 pg/ml.
Thymidine Incorporation Assay.
Cells were seeded in 96-well plates at approximately 80% confluence (2 × 106 cells/well) and incubated in serum-free medium for 24 hrs to induce quiescence and then 100 nM ET-1 was added. After 24 hrs, when cells were confluent, 1 μCi of [methyl-3H] thymidine (5.0 Ci/mmol) was added to each well. Six hours later, culture media were removed and cells were washed three times with phosphate-buffered saline, treated with 10% trichloroacetic for 15 mins, washed twice with 100% ethanol, and solubilized in 0.4 N sodium hydroxide. The cell-associated radioactivity was then determined by liquid scintillation counting. Responses to all treatments were assayed in sextuplicate, and results were expressed as the means of three separate experiments.
Results
Using RT-PCR, we first detected the expression of mRNA of the various components of the endothelin system ET-1, ETAR and ETBR, in two cell lines of ovarian carcinoma, HEY and OVCA 433. As shown in Figure 1A, RT-PCR–amplified cDNA fragments for ET-1 (462 base pairs [bp]), ETAR (367 bp) and ETBR (529 bp), were detected in both cell lines. The levels of ET-1 mRNA are mirrored in their ET-1 secretion (Fig. 1B). In fact, the ET-1 levels released from HEY and OVCA 433 cells were within the physiologic range needed for activation of ETAR in an autocrine fashion.
To examine whether the mitogenic activity of ET-1 was mediated by ETAR, we coincubated the cells with ET-1 and ZD4054. ZD4054 is a potent antagonist of ETAR, with no evidence of ETBR blockade. The lack of affinity for ETBR indicates that ZD4054 has the potential to block the multiple pathologic processes related to malignancy that are mediated by ETAR while allowing the beneficial processes mediated by ETBR, such as apoptosis and clearance of ET-1 (17). The stimulatory activity of 100 nM ET-1 on [3H]thymidine incorporation by HEY and OVCA 433 cells was completely blocked in the presence of 1 μM ZD4054. Furthermore, blocking the interaction of endogenous ET-1 with ETAR using ZD4054, we obtained a significant (P ≤ 0.0001) inhibition of cell proliferation (Fig. 2), demonstrating that the potent ETAR antagonist ZD4054 is capable of reversing mitogenic responses induced by ET-1.
Discussion
The ET-1/ETAR autocrine pathway contributes to ovarian cancer progression by inducing cell proliferation, apoptosis protection, angiogenesis, and invasiveness, indicating that the pharmacologic inhibition of the ETAR signaling pathway may improve current treatment of these tumors. In the search for pharmacologic agents capable of inhibiting endothelin receptors, ZD4054 is of particular interest. In receptor binding studies, in fact, ZD4054 selectively binds ETAR with high affinity, whereas no binding was detected to ETBR (which, therefore, does not interfere with ET-1 clearance) (17). In a randomized placebo-controlled trial, a single oral dose of ZD4054 reduced forearm vasoconstriction in response to brachial artery infusion of ET-1, thus providing further in vivo evidence of ETAR blockade. ETBR blockade was also assessed in an ascending, single-dose, placebo-controlled trial in volunteers. For all doses of ZD4054, mean plasma ET-1 concentration was within the reference range, confirming the specificity of ZD4054 for ETAR, with no activity for ETBR. In view of the above, in the present study we employed two ovarian cell lines characterized by the ET-1/ETAR autocrine loop to assess the in vitro antiproliferative activity of ZD4054. We demonstrated that in the two cell lines, which express both ETAR and ETBR mRNA and produce ET-1, ZD4054 is capable of significantly inhibiting both the baseline and the ET-1–induced proliferation. All together, the results identify ZD4054 as a specific blocking agent of ETAR with the potential of inhibiting the multiple tumor-promoting processes elicited by the activation of this receptor in ovarian cancer and other malignancies expressing ETAR.
(A) ET-1, ETAR, and ETBR mRNAs detected by RT-PCR in HEY and
OVCA 433 cells. PCR products in HEY cells of 462 bp for ET-1, 367 bp for
ETAR, and 535 bp for ETBR are shown as visualized by ethidium
bromide. (B) Time-dependent ET-1 production in HEY and OVCA 433 cell lines. Cells were
incubated for 24 hrs in serum-free medium before the experiment. The conditioned media
were collected at the indicated times, and the ET-1 secretion was determined using an
ELISA kit. Data are presented as mean of results from three experiments, each of which
was performed in triplicate. Bars, ±SD. Effect of ZD4054 on [3H]thymidine incorporation. Quiescent ovarian
carcinoma cell lines (HEY and OVCA 433) were treated with ET-1 at the indicated
concentrations for 24 hrs before measuring [3H]thymidine incorporation.
Cells were incubated for 15 mins with 1 μM ZD4054 in the absence or
in the presence of 100 nM ET-1. Bars, mean ± SD of data from three
independent experiments, each of which was performed in sextuplicate. *,
P < 0.0001 compared to control; **, P <
0.0001 compared to ET-1.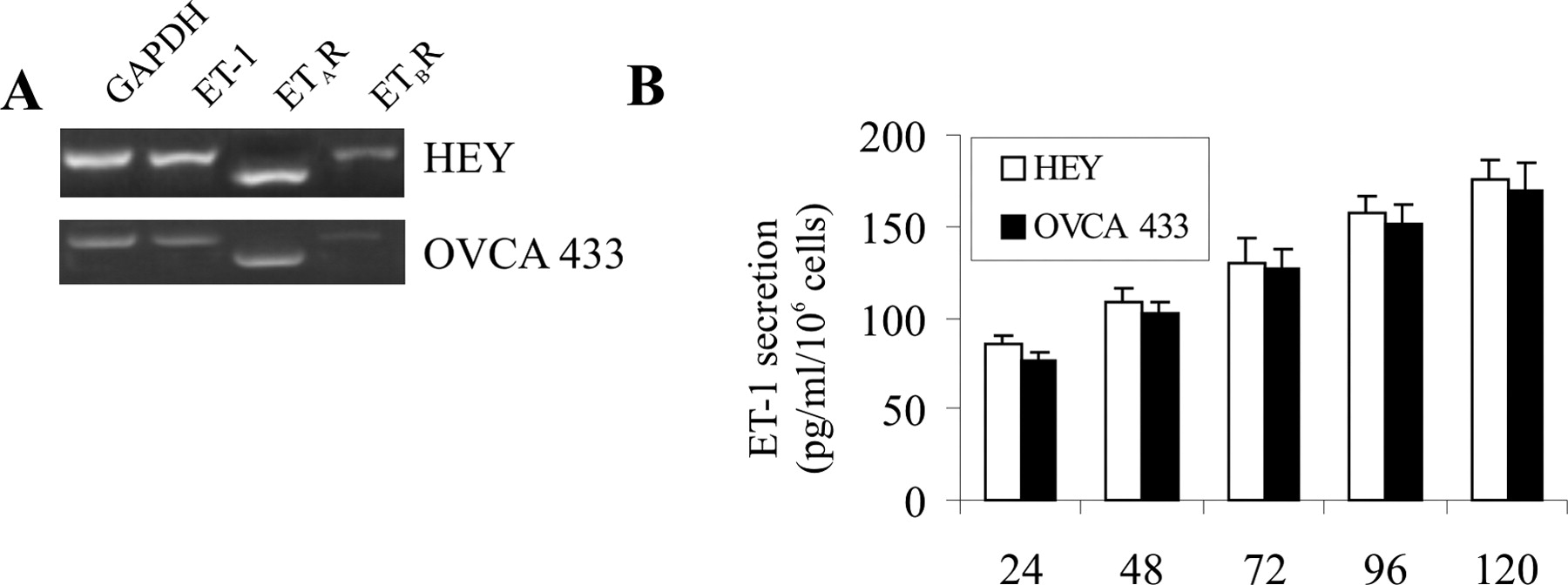

Footnotes
This work was supported by grants from the Associazione Italiana Ricerca sul Cancro, Consiglio Nazionale delle Ricerche-MIUR, the Ministero della Salute and AstraZeneca.