
Abstract
Long-term use of morphine in pain management leads to adverse effects, such as development of antinociceptive tolerance. We have previously shown the involvement of central endothelin (ET) mechanisms in morphine analgesia and development of tolerance in vivo. The present study was conducted to investigate the in vitro mechanism of interaction of the ETA receptor antagonist, BMS182874, and morphine during acute and chronic morphine tolerance in SH-SY5Y cells. SH-SY5Y cells were exposed to acute and chronic treatment with vehicle, morphine, ET-1, BMS182874, or morphine plus BMS182874. Activation of G-protein–coupled receptors in SH-SY5Y cells was determined using [35S]GTPγS binding assays. Acute morphine treatment produced a concentration-dependent increase in GTP binding. Median effective concentration (EC50) values were significantly decreased after acute morphine treament, suggesting sensitization of opioid receptors. Chronic morphine treatment produced a lower maximal response of GTP binding compared with both control (vehicle treated) and acute morphine treatment, indicating uncoupling of G-proteins. Acute and chronic exposure of cells to ET-1 did not affect changes in ET-1–induced GTP binding. BMS182874 treatment alone (acute or chronic) did not produce G-protein activation. However, in cells chronically cotreated with 10 μM morphine and 1 μM BMS182874, morphine-induced GTP stimulation was significantly higher than control (vehicle treated). The EC50 value after control treatment was 414 nM, and was significantly increased in chronically morphine-treated cells (>1000 nM ). However, the EC50 value in cells receiving a chronic treatment of BMS182874 and 63 nM morphine was significantly reduced compared with control (vehicle treated) and chronic morphine treatment. ETA antagonists significantly enhance the coupling of G-protein to opioid receptors. Therefore, we propose that restoration of morphine antinociception by ETA antagonists in morphine-tolerant animals is likely via a G-protein mediated mechanism.
Introduction
Long-term opioid therapy for providing sustained analgesia is associated with tolerance and physical dependence. Although compounds of extremely high potency with varying affinity and efficacy at all opioid receptor subtypes have been produced, the problem of tolerance to and dependence on these agonists persists (1). The multiplicity of opioid receptors (μ, δ, and κ) and the functional interaction between their coupling mechanisms leads to complex cellular and biochemical changes in opioid tolerance (2, 3). G-protein–coupled receptor desensitization and intracellular changes in second messengers in morphine tolerance also play a role in analgesic tolerance to opioids (4).
Despite intensive research documenting the occurrence of antinociceptive tolerance, the mechanisms underlying this phenomenon still remain unclear. A variety of diverse agents have been reported to modify opioid tolerance (5–8). However, many of these agents interact with various neurotransmitters, producing unwanted side effects and limiting their clinical use in treatment of tolerance. Because of the multiple mediators in pathogenesis of opiate tolerance, current therapeutic strategy focuses on using combination therapy with other analgesics to produce enhanced analgesia. Inhibition of excitatory receptors that may link with opioid receptors after chronic exposure to opiate analgesics is another emerging area of research. We propose that endothelin (ET), an endogenous neuropeptide, may be an important factor in mediating opiate tolerance. We have previously shown that ETA receptor antagonists enhance morphine analgesia (9) and restore the analgesic response of morphine in morphine-tolerant rats (10).
Chronic administration of morphine leads to modulation of Gi/Go-coupled receptors; however, effects of chronic morphine exposure and corresponding changes in the ET system have not been investigated. We propose that there is a possible role of the ET system in pharmacologic actions of opioid analgesics. Chronic morphine treatment results in functional uncoupling of μ-opioid receptors and G-proteins in cell cultures and animal models. Neuroblastoma cell lines have been used to study opioid tolerance. Neuroblastoma SH-SY5Y cells expressing opioid as well as ETA receptors were used for this study. The aim was to determine the activation of G-proteins coupled to opioid receptors in response to acute and chronic treatment with morphine, ET-1, and the ETA receptor antagonist, BMS182874, in SH-SY5Y cells, using a [35S]GTPγS binding assay.
Materials and Methods
Drugs and Chemicals.
Morphine sulfate (Mallinckrodt Chemical Co., St. Louis, MO) was dissolved in distilled deionized pyrogen-free water. BMS182874 (Tocris Pharmaceuticals Inc., Ellisville, MO) was dissolved in 20% dimethylsulfoxide prepared in sterile saline. ET-1 (American Peptide Company Inc., Sunnyvale, CA) was dissolved in 0.1% bovine serum albumin. [35S]GTPγS (specific activity, 1000 Ci/mmol; Amersham Pharmacia Biotech, Piscataway, NJ) was diluted in 50 mM Tris buffer, pH 7.4. Guanosine-5′-diphosphate (GDP) and unlabeled GTPγS (Sigma, St. Louis, MO) were dissolved in 50 mM Tris buffer, pH 7.4.
Estimation of [35S]GTPγS Binding in SH-SY5Y Neuroblastoma Cells.
Human neuroblastoma SH-SY5Y cells (American Type Culture Collection, Manassas, VA) were used for this study because this cell line expresses both opioid and ETA receptors. This is a prototypical model system frequently used to study cellular aspects of opioid tolerance (11). Cells were cultured in monolayers in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum, 100 IU/ml penicillin, and 100 μg/ml streptomycin, and maintained at 37°C in a humidified 5% CO2 atmosphere. For acute treatment, separate cell plates were treated with different concentrations of morphine, ET-1, BMS182874, and morphine plus BMS182874 for 1 hr. For chronic treatment, cells were treated for 48 hrs with morphine, ET-1, BMS182874, or morphine plus BMS182874. Cells were harvested after drug treatments in ice-cold phosphate-buffered saline containing 1 mM EDTA and dispersed using a glass homogenizer. After centrifugation at 500 g, the cell pellet was suspended in 50 mM Tris-HCl buffer, pH 7.4; 100 mM NaCl; and 10 mM MgCl2·H2O (Buffer A) and homogenized again using a glass homogenizer. The resultant homogenate was centrifuged at 20,000 g, and the pellet was collected, washed in Buffer A, and centrifuged again. The pellet was finally resuspended in Buffer A, to give a protein concentration of 100–200 μg/ml, and the membrane preparation was prepared as described previously (12). All procedures were performed in triplicate. The final pellet was resuspended to a protein concentration of approximately 50–75 μg. Membranes (50–75 μg protein) were incubated in Buffer A containing 80 pM [35S]GTPγS, 30 μM GDP, and varying concentrations of ET-1 and BMS182874, in a volume of 1 ml for 60 mins at 30°C. Nonspecific binding was determined using unlabeled 10 μM GTPγS. Bound and free [35S]GTPγS were separated by vacuum filtration using GF/B filters and quantified by liquid scintillation counting. Specific binding (determined by subtracting nonspecific binding from total binding) was expressed as femtomoles per milligram of protein (mean ± SEM). The concentration of protein in the samples was determined by Lowry’s method (13).
Statistical Analysis.
Data was analyzed by one-way analysis of variance followed by a Bonferroni test. A level of P < 0.05 was considered significant.
Results
Stimulation of [35S]GTPγS Binding After Acute Treatment of Neuroblastoma Cells with Morphine, ET-1, and BMS182874.
Morphine increased [35S]GTPγS binding in SH-SY5Y cells in a concentration-dependent manner. The median effective concentration (EC50) values for the different treatments with morphine indicate that the curve shifted to the left with increasing concentrations (Fig. 1a). The EC50 values of morphine for GTP binding after acute treatment with 100 nM, 1 μM, and 10 μM morphine were 426.67 nM, 186.61 nM, and 106.45 nM, respectively, indicating that acute treatment with morphine may increase the sensitivity of the opioid receptors and produce a higher G-protein activation. ET-1–induced GTP stimulation also increased in a concentration-dependent manner (Fig. 1b). The EC50 values for acute treatment with ET-1 did not differ significantly (P > 0.05) between treatments (126.98 nM with 100 nM ET-1; and 95.47 nM with 1 μM ET-1). These findings indicate that ET-1 did not cause sensitization of ET receptors. The EC50 values after acute treatment with increasing concentrations of BMS182874 (1 nM, 100 nM, and 1 μM) were greater than 1000 nM. This indicates that BMS182874 did not activate GTP binding in SH-SY5Y cells.
Maximal stimulation after combined acute treatment with 1 μM morphine plus 100 nM BMS182874 (76.55 ± 1.08%) was similar (P > 0.05) to control-treated cells (79.07 ± 2.86%; Fig. 1c). The EC50 value was significantly lower in morphine plus BMS182874–treated cells (73.95 nM), compared with control treatment (416.81 nM). Activation of G-proteins was observed even with lower concentrations of morphine. Therefore, BMS182874 enhanced the morphine-induced activation of G-proteins in SH-SY5Y cells.
Stimulation of [35S]GTPγS Binding After Chronic Treatment of Neuroblastoma Cells with Morphine, ET-1, and BMS182874.
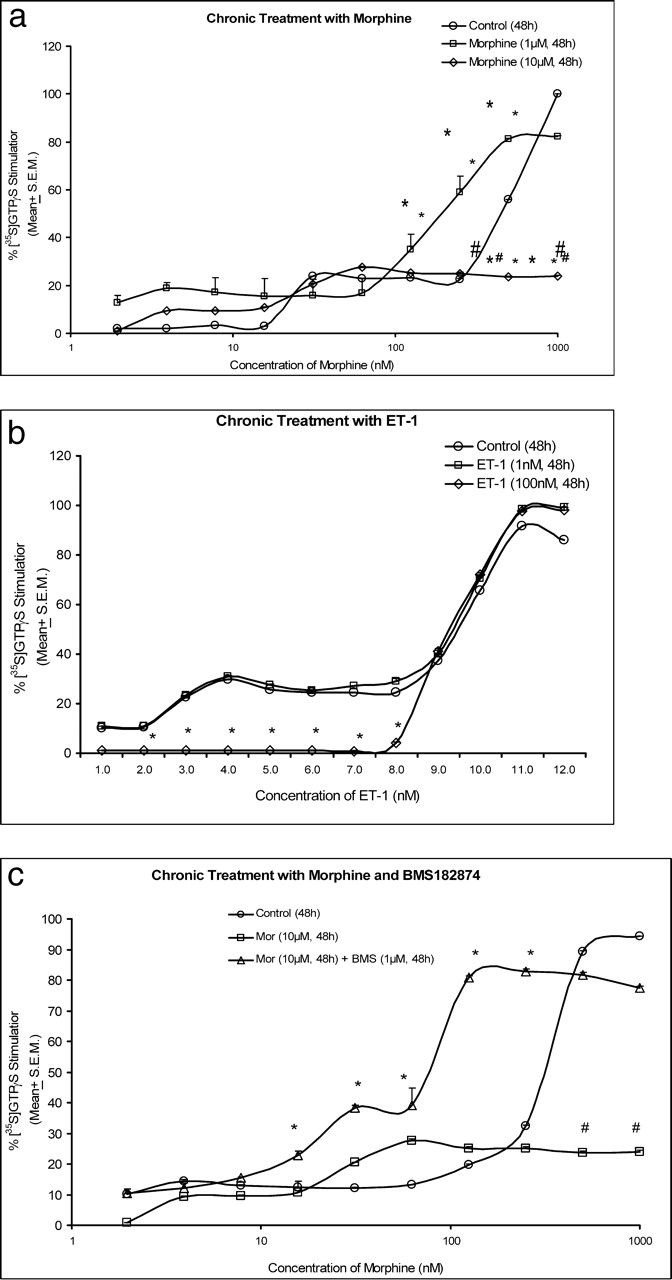
Chronic morphine (1 μM) did not change the basal [35S]GTPγS binding (Fig. 2a). However, the EC50 (127.85 nM) was greater (P < 0.05) compared with cells receiving the control treatment. Chronic exposure to 10 μM morphine significantly reduced (P < 0.05) GTP binding compared with control treatment, and the maximal activation in cells treated with 10 μM morphine for 48 hrs was 28.06 ± 1.57% (Fig. 2a).
Results obtained with chronic exposure to 1 μM ET-1 indicate that, at lower ET-1 concentrations, GTP binding was significantly reduced (P < 0.05) compared with control cells, and compared with 100 nM treatment. This suggests that ET receptors were also uncoupled from G-proteins (Fig. 2b). Chronic treatment with BMS182874 did not activate GTP binding in SH-SY5Y cells with increasing concentrations of the antagonist.
Stimulation of [35S]GTPγS Binding After Chronic Treatment of Neuroblastoma Cells with Morphine and Morphine Plus BMS182874.
Two treatments were investigated to determine the effect of chronic treatment of morphine with ETA receptor antagonists. Percent stimulation of [35S]GTPγS binding in neuroblastoma (SH-SY5Y) cells treated with morphine and BMS182874 is shown in Figure 2c. Basal binding was statistically similar (P > 0.05) in control (vehicle treated) and morphine plus BMS182874–treated cells. Maximal stimulation after chronic treatment with 10 μM morphine (23.82 ± 0.68%) was significantly lower (P < 0.05) compared with control treatment. The EC50 value in 10 μM morphine–treated cells was >1000 nM. This was significantly higher (P < 0.05) compared with control-treated cells (520.46 nM). The maximal stimulation in 1 μM morphine plus 100 nM BMS182874–treated cells was 92.53 ± 0.23%. The EC50 value of morphine in these cells was 66.43 nM. The EC50 value for morphine-stimulated [35S]GTPγS binding in 10 μM morphine and 1 μM BMS182874–treated cells was 62.78 nM. These values were significantly lower (P < 0.05) than the EC50 value for morphine-stimulated [35S]GTPγS binding in control-treated cells (413.78 nM; Fig. 2c). These results indicate that G-proteins were activated to a significantly greater extent when cells were exposed chronically to the combined treatment with morphine plus BMS182874 compared with morphine alone. As observed previously, the maximal stimulation with 1 μM BMS182874 in control cells was 5.16 ± 0.71%, indicating that BMS182874 did not stimulate GTP binding. However, in combination with morphine, the maximal stimulation was 96.54 ± 3.69%, which was significantly higher (P < 0.05) than in control cells (Fig. 2c).
Discussion
The agonist-mediated increase in G-protein function is a critical step in receptor activation and function. The extent of G-protein activation is proportional to the agonist-bound receptors, and the attenuation of G-protein activation is a significant factor in development of opiate tolerance. SH-SY5Y cells endogenously express opioid receptors, and chronic morphine attenuates the μ-opioid receptor–activated [35S]GTPγS binding (14). We investigated the effects of morphine, ET-1, and ETA receptor antagonists in SH-SY5Y cells. A dose-dependent increase in [35S]GTPγS binding in response to increasing doses of morphine was observed after acute treatment of SH-SY5Y cells with morphine. We observed a sensitization of G-protein activation by morphine, as seen by a leftward shift of the curve. Morphine-induced [35S]GTPγS binding was significantly reduced after chronic treatment with morphine. Therefore, activation of G-proteins was significantly attenuated in chronic morphine–treated cells.
Acute treatment of SH-SY5Y cells with ET-1 showed a dose-dependent increase in [35S]GTPγS binding. The response was similar to ET-1–induced [35S]GTPγS binding in control cells. These data suggest that acute treatment with ET-1 did not sensitize ET receptors. In cells treated chronically with ET-1, the maximum response was similar to control cells. Increasing doses of the ETA receptor antagonist, BMS182874, showed no stimulation of GTP binding in these cells after acute or chronic treatment. Therefore, BMS182874 alone did not activate G-proteins. In cells treated chronically with morphine plus ETA receptor antagonist, GTP binding was significantly higher than in cells treated chronically with morphine alone, but similar in response to control treatment. There was significant increase in G-protein activation in response to morphine after combined treatment with BMS182874. This suggests that tolerance did not develop to chronic morphine exposure in presence of the ETA receptor antagonist, and that G-protein activation was produced by morphine, even at lower concentrations. This enhanced binding and signaling by combined treatment with ETA receptor antagonists indicates the ability of these antagonists to restore G-protein coupling to opioid receptors. The findings of the present study suggest a possible mechanism of interaction between morphine and ETA receptor antagonists during tolerance.
(a) Morphine-stimulated [35S]GTPγS binding in SH-SY5Y cells after acute
treatment with morphine. Values are mean ± SEM; n = 3 per group. (b)
ET-1–stimulated [35S]GTPγS binding in SH-SY5Y cells after acute treatment
with ET-1. Values are mean ± SEM; n = 3 per group. (c)
Morphine-stimulated [35S]GTPγS binding in SH-SY5Y cells after acute
treatment with morphine plus BMS182874. Values are mean ± SEM; n = 3
per group. *P < 0.05 compared with control vehicle–treated
cells. (a) Morphine-stimulated [35S]GTPγS binding in SH-SY5Y cells after chronic
treatment with morphine. Values are mean ± SEM; n = 3 per group.
*P < 0.05 in comparison with control vehicle–treated cells.
#P < 0.05 in comparison with 1 μM
morphine–treated cells. (b) ET-1–stimulated [35S]GTPγS binding in SH-SY5Y
cells after chronic treatment with ET-1. Values are mean ± SEM; n = 3
per group. *P < 0.05 in comparison with 100 nM
ET-1–treated cells. (c) Morphine-stimulated [35S]GTPγS binding in SH-SY5Y
cells after chronic treatment with morphine plus BMS182874. Values are mean ± SEM;
n = 3 per group. *P < 0.05 compared with
control vehicle-treated cells; #P < 0.05 compared with 10
μM morphine–treated cells.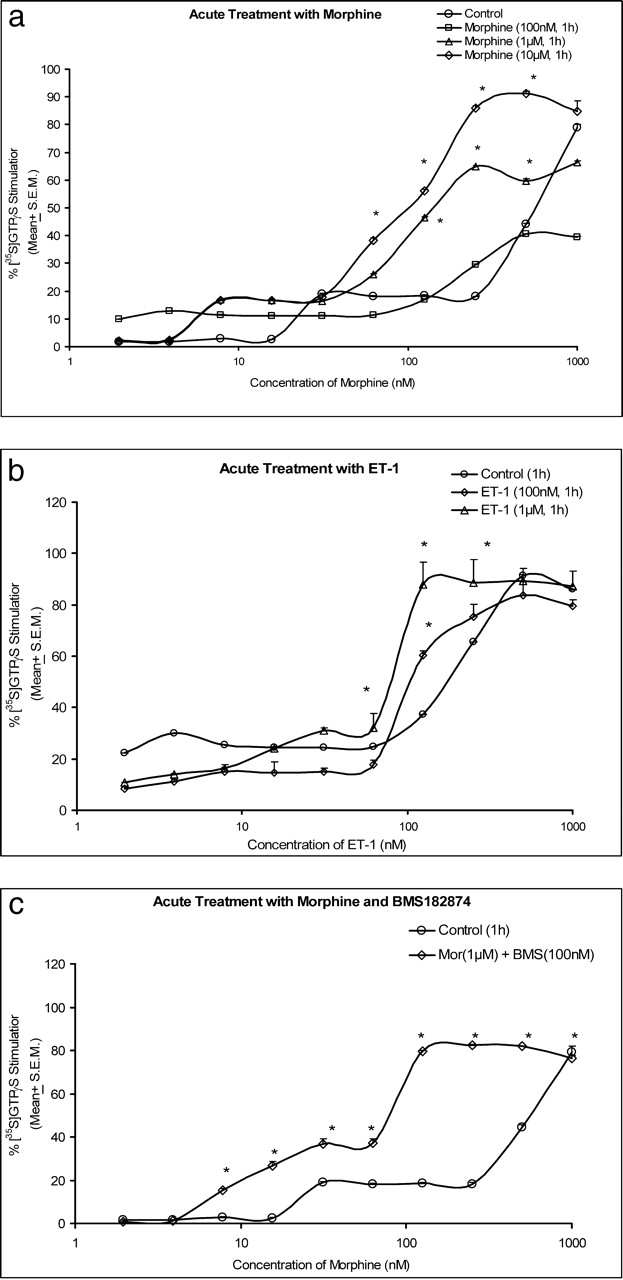
Footnotes
The present study was funded by Chicago Labs Inc., Chicago, IL.