
Abstract
Sepsis involves a heterogeneous class of syndromes, and septic shock, a severe form of sepsis, is associated with the development of progressive damage in multiple organs. The present study examined the time-dependent alterations of endothelin-1 (ET-1) and vascular endothelial growth factor (VEGF) levels in liver tissue in a septic rat model. Healthy male Wistar rats aged 15 weeks received 15 mg/kg lipopolysaccharide (LPS) and were sacrificed at different time points (1, 3, 6, and 10 hrs after treatment). Rats that did not receive LPS were considered to be controls. A 28-fold increase in the ET-1 level was observed in liver tissue 10 hrs after LPS administration. VEGF was also altered in hepatic tissue in a time-dependent manner. A gradual increase of VEGF expression in liver tissue after LPS administration was observed. Expression of Flt-1, the vascular permeability receptor of VEGF, was also increased in liver tissue after LPS administration. ET-1 is a potent vasoconstrictor and, therefore, may play a role in the regulation of hepatic perfusion in a sepsis model. On the other hand, VEGF may be involved in capillary leakage in liver tissue after LPS administration. The present findings suggest that there might be a loss of balance between the ET-1 and VEGF levels in the septic liver at different time points, which could contribute to the pathogenesis of acute liver injury in endotoxemia.
Sepsis involves a life-threatening and heterogeneous class of syndromes caused by a systemic inflammatory response to infection. Septic shock is the severe form of sepsis, is associated with the development of progressive damage in multiple organs, and is the leading cause of patient mortality in intensive care units (1). Septic shock is caused by lipopolysaccharide (LPS) and other bacterial products. Accumulating evidence indicates a role for vasoactive substances and cytokines in this disease process.
Vascular endothelial growth factor (VEGF) is a major angiogenic factor, as well as a potent vascular permeability factor (2). VEGF plays a crucial role in a variety of diseases through the promotion of angiogenesis and by means of its vasopermeability effect. It also plays a critical role as a potent proinflammatory cytokine in a variety of physiologic and pathologic immune responses (2). Because the development of capillary leak is a common occurrence in septic patients, one might reasonably speculate that several sepsis-associated mediators might induce VEGF production. Recently, a potential role for VEGF in sepsis has been evaluated. For example, plasma VEGF levels are elevated during severe sepsis (3). Plasma VEGF levels are also associated with disease severity and mortality (3). However, the organ-specific expression of VEGF in sepsis has not been clarified.
Endothelin-1 (ET-1) is a potent vasoconstrictor with vasoproliferative activity that has been postulated to participate in the pathogenesis of a number of diseases. During sepsis, the plasma ET-1 level has been shown to be significantly higher (4). The possible involvement of the ET system in human septic shock is further supported by a clear correlation between ET plasma levels and morbidity and mortality in septic patients (5, 6). Infusion of ET-1 to humans causes cardiovascular changes that resemble those seen during sepsis (i.e., decreased cardiac output and vasoconstriction in the pulmonary, renal, and splanchnic circulation) (6). Thus, we speculate that ET-1 might have some role in the regulation of liver perfusion in endotoxemia, because infusion of ET-1 into the portal vein markedly and dose-dependently reduces sinusoidal blood flow (7).
In the present study, a rat model of endotoxemia was produced by LPS administration. Time-dependent alterations of VEGF and ET-1 levels were examined in liver tissue in the septic rat model. Flt-1, the vascular permeability receptor of VEGF, was also investigated, and the extent of liver injury induced by LPS was assessed histologically.
Materials and Methods
Animal Preparation.
Male Wistar rats were used in all experiments. Endotoxemic shock was induced by ip administration of 15 mg/kg LPS from Escherichia coli O55:B5 dissolved in sterile saline. Groups of 12 animals were killed by pentobarbital at different time points after LPS administration (1, 3, 6, and 10 hrs). The control group received an equal volume of sterile saline (2 ml/kg) instead of LPS. At the indicated time, liver tissues were gently harvested, immediately frozen in liquid nitrogen, and stored at −80°C. For paraffin sections, the liver tissue was postfixed in 4% paraformaldehyde overnight and processed routinely for paraffin embedding.
All animals received care that was in compliance with University of Tsukuba guidelines, and the University of Tsukuba Graduate School of Medicine Animal Care and Use Committee approved the experimental procedure.
Histopathological Analysis.
For histopathological analysis, the tissues were fixed in 4% buffered formalin solution, dehydrated, embedded in paraffin, and then sliced into 5-μm–thick sections. After being deparaffinized, hematoxylin-eosin–stained slides were prepared by using standard methods.
Hemodynamic Analysis.
On the day of the experiment, the rats were anesthetized with pentobarbital sodium (40 mg/kg ip), and a microtip pressure transducer catheter (SPC-320; Millar Instruments, Houston, TX) was inserted into the left carotid artery. Arterial blood pressure and heart rate were monitored and recorded with the use of a polygraph system (an AP-601G amplifier [Nihon Kohden, Tokyo, Japan], an AT-601G tachometer [Nihon Kohden], and a WT-687G thermal-pen recorder [Nihon Kohden]).
Enzyme Immunoassay for Hepatic ET-1.
The concentration of ET-1 in hepatic tissue extracts and in plasma was determined using an ET-1 Enzyme Immuno Assay Kit (Immuno-Biological Laboratories, Fujioka, Japan). This kit is a solid-phase sandwich enzyme-linked immunoabsorbent assay (ELISA) that uses 2 kinds of highly specific antibodies. Tetraethyl benzidine (TMB) is used as a coloring agent (chromogen), the strength of which is in proportion to the quantity of ET-1. In hepatic tissue extracts, an equal volume of 0.1% trifluoroacetic acid (TFA) in water was added. Then the sample was centrifuged at 3000 g for 15 mins at 4°C for clarification, and the supernatant was saved. After a 200-mg C 18 Sep-Pak column (part number 235D1; Waters Inc., Milford, MA) was washed four times with 1 ml of 60% acetonitrile in 0.1% TFA and four times with 5 ml of 0.1% TFA in water, the supernatant was applied to the column and washed four times with 5 ml of 0.1% TFA in water. The sample was eluted slowly by washing it three times with 1 ml of 60% acetonitrile in 0.1% TFA in water, and the eluant was collected in a plastic tube. The eluant was then evaporated to dryness with a centrifugal concentrator under vacuum. The sample was reconstituted with assay buffer and measured immediately according to the manufacturer’s instructions.
VEGF Immunoassay.
The VEGF protein level in hepatic tissue and in plasma was measured by immunoassay (VEGF Immunoassay kit; R and D Systems, Minneapolis, MN). This assay uses the quantitative sandwich enzyme immunoassay technique.
Quantitative Real-Time Polymerase Chain Reaction (RT-PCR).
Total tissue RNA was isolated by acid guanidinium thiocyanate-phenol-chloroform extraction with Isogen (Nippon Gene, Toyama, Japan). In brief, the liver tissue was homogenized in Isogen (100 mg tissue per 1 ml Isogen) with a Polytron tissue homogenizer (model PT10SK/35; Kinematica, Lucerne, Switzerland). The precipitated RNA was extracted with chloroform, precipitated with isopropanol, and washed with 75% (vol/vol) ethanol. The resulting RNA was resolved in diethyl pyrocarbonate–treated water, treated with DNase I (Takara, Shiga, Japan), and extracted again with Isogen to eliminate the genomic DNA. The RNA concentration was determined spectrophotometrically at 260 nm. Total tissue RNA was primed with 0.05 μg of oligo d (pT)12–18 and reverse transcribed by omniscript reverse transcriptase by means of a first-strand cDNA synthesis kit (Qiagen, Tokyo, Japan). The reaction was performed at 37°C for 60 mins.
The mRNA expression level of Flt-1 in the liver was analyzed by quantitative RT-PCR with a TaqMan probe using an ABI Prism 7700 Sequence Detector (Perkin-Elmer Applied Biosystems, Foster, CA). The gene-specific primers and TaqMan probes were synthesized from Primer Express software, version 1.5 (Perkin-Elmer), according to the published cDNA sequences for Flt-1 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA. The sequences of the oligonucleotides were as follows: Flt-1 forward: 5′-TCGGCTGTCCATGAAAGTGAAGT-3′, Flt-1 reverse: 5′-GCGGGTACGCCATCTTTTAAC-3′, and Flt-1 probe: 5′-CCTCGCCAGAAGTCGTATG-3′; GAPDH forward: 5′-GTGCCAAAAGGGTCATCATCTC-3′, GAPDH reverse: 5′-GGTTCACACCCATCACAAACATG-3′, and GAPDH probe: 5′-TTCCGCTGATGCCCC-3′.
The expression of GAPDH mRNA was used as an internal control. The PCR mixture (25 μl total volume) consisted of 450 nM of both forward and reverse primers for Flt-1 and for GAPDH (Perkin-Elmer), 200 nM of FAM-labeled primer probes (Perkin-Elmer), and TaqMan Universal PCR Master Mix (12.5 μl; Perkin-Elmer). Each PCR amplification was performed in triplicate, using the following profile: 1 cycle of 95°C for 10 mins, and 40 cycles of 94°C for 15 secs and 60°C for 1 min. To obtain the standard curve in the quantitative RT-PCR, serial dilutions of rat lung cDNA were performed using a range of concentrations (1×, 2×, 4×, 8×, and 16×). A water-based reaction mixture was prepared as a negative control.
Statistical Analysis.
Results are expressed as mean ± SD, and the sample number equals the number of animals in each group. Means were compared by one-factor analysis of variance, followed by the Scheffé’s test to account for the problem of multiple comparisons. Differences were considered to be statistically significant at a P value of < 0.05.
Results
Both the systolic and diastolic blood pressure levels were significantly lower at different time points than those in control rats after LPS administration. Livers from control animals showed no morphological abnormalities. However, in rats that received LPS, hepatocytic atrophy and necrosis, as well as neutrophil infiltration, were observed in liver tissues.
ET-1 levels determined by ELISA gradually became significantly elevated in liver tissue after LPS administration (Fig. 1). Furthermore, the immunoreactive ET-1 level in plasma was significantly higher after LPS administration, compared with that in the control group (0.77 ± 0.37 pg/ml in the control group, 8.30 ± 2.23 pg/ml 1 hr after receipt of LPS, 15.28 ± 12.65 pg/ml after 3 hrs, 10.11 ± 5.86 pg/ml after 6 hrs, and 3.96 ± 1.93 pg/ml after 10 hrs).
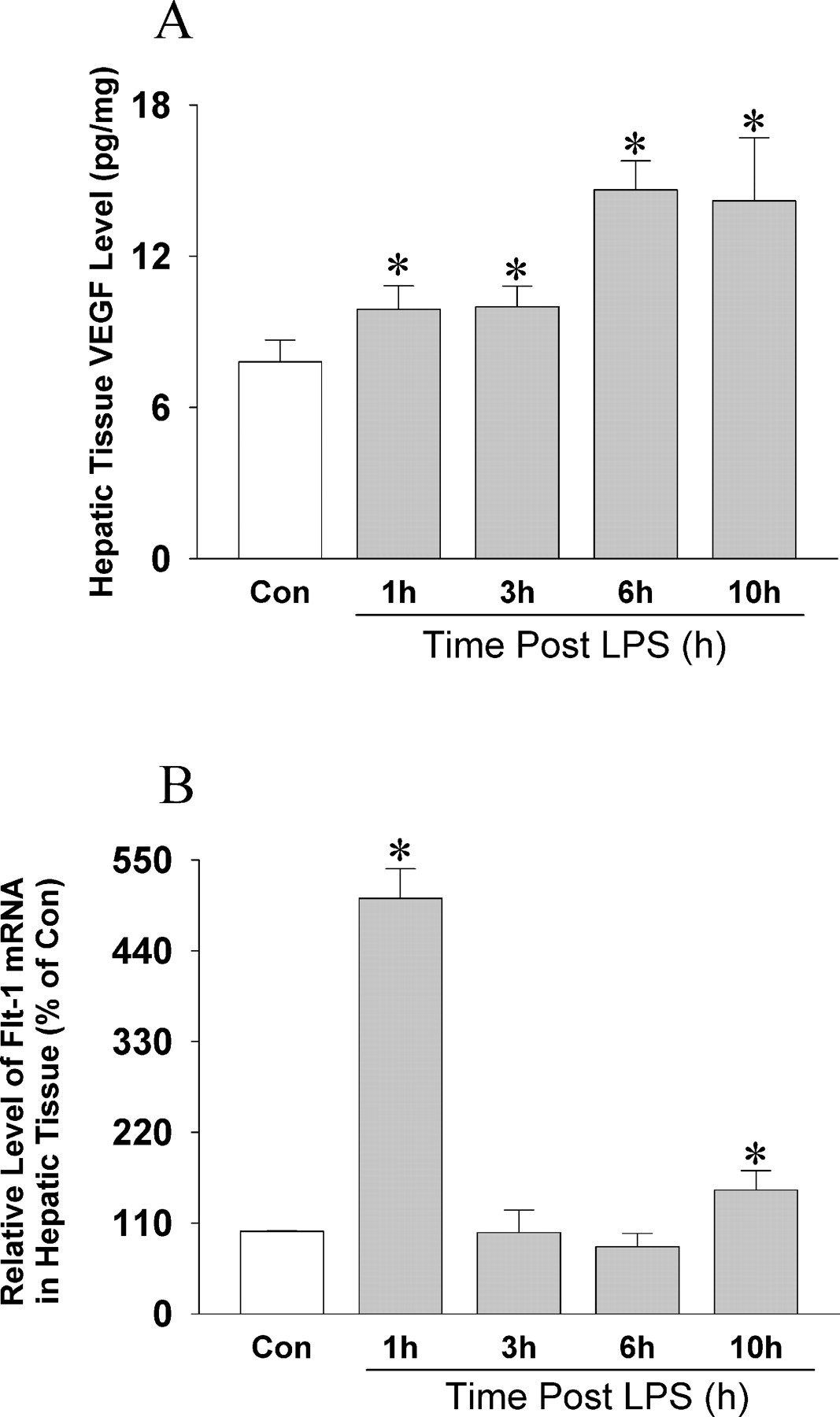
The VEGF protein level was also increased in hepatic tissue in a time-dependent manner (Fig. 2A). The plasma VEGF level also increased in rats that received LPS as the severity of sepsis increased (47 ± 13 pg/ml in the control group, 64 ± 20 pg/ml 3 hrs after receipt of LPS, and 73 ± 30 pg/ml after 10 hrs). The expression of the gene encoding Flt-1, the vascular permeability receptor of VEGF, also significantly increased in liver tissue after LPS administration.
Discussion
The present study demonstrates that the expression of ET-1 in liver tissue increases in a time-dependent manner in rats that received LPS. Moreover, the expression of another vasoregulatory molecule, VEGF, in liver tissue also increases with the progression of sepsis. It should be noted that the plasma levels of both ET-1 and VEGF are increased in rats with LPS-induced sepsis. Moreover, the features of acute liver injury are evident in septic liver tissues. Both the systolic and diastolic blood pressure were significantly decreased in LPS-treated rats.
Acute hepatic failure after severe trauma and sepsis is still associated with a high mortality rate (8). LPS is a component of the cell walls of gram-negative bacteria (9, 10). Proinflammatory mediators, such as LPS and tumor necrosis factor-α (TNF-α), can directly lead to hepatocellular damage (10). A large number of inflammatory mediators are involved in the complex pathophysiology of sepsis and endotoxemia. Several of the manifestations, such as pulmonary hypertension, regional hypoperfusion, and extravasation of leukocytes, can be attributed to endothelin (11, 12). ET-1 is a powerful vasoactive peptide and is secreted by various cells in the liver, including vascular and sinusoidal endothelial cells and hepatic stellate cells (13–15). ET-1 causes strong vasoconstrictive action after binding to its receptor (16). ET receptors have been classified into two receptor subtypes, ETA and ETB, and both are present in various liver cells. These receptors are involved in many biological and pathophysiological responses, including hepatic microcirculation (17, 18). Release of ET-1 from sinusoidal endothelial cells acting on Ito cells mediating contraction is one mechanism suggested for flow regulation (19). In the current study, the ET-1 peptide level increased in hepatic tissue in a time-dependent manner. Oldner et al. (20) reported that endotoxemia induced a 6-fold increase in portal-hepatic vascular resistance, and this resistance was effectively counteracted by nonselective ET-receptor antagonism. Thus, the time-dependent upregulation of ET-1 in the septic liver may be highly important in the regulation of hepatic blood flow during sepsis. Controversial reports exist concerning the role of ET receptors in the regulation of hepatic blood flow. Although both ETA and ETB receptors have been reported to mediate the vasoconstrictor response in the liver (21), Ruetten et al. (22, 23) observed reduced severity of hepatocellular injury in endotoxemic rats treated with ETB receptor agonists but not in those treated with ETA receptor antagonists. Thus, the effects of ET on the pathogenesis of liver injury are complex and heterogeneous and may vary between experimental models and species. The liver is, however, a highly important organ under septic conditions, and more-detailed studies should focus on the gene and protein expressions and regulations of different components of the ET system in a time-dependent manner after LPS administration.
VEGF has been extensively investigated recently in various hepatic diseases, such as primary and secondary hepatocellular carcinoma, liver cirrhosis, hepatitis, and even benign tumors in the liver (24). VEGF has been verified to be closely involved in the development and metastases of hepatocellular carcinoma and correlated with a high risk of hepatic metastases and a poor prognosis in gastrointestinal cancer (24). Antibodies against VEGF or other drugs to suppress VEGF expression have also been successful at controlling the division of hepatocellular carcinoma cells and spread of metastases in vitro and in animal models (24). Tissue expression and circulating levels of VEGF protein tend to increase during acute and chronic hepatitis and to decrease during cirrhosis (24).
In the present study, VEGF protein and the gene expression of its vascular permeability receptor, Flt-1, were upregulated in the liver after LPS administration. LPS-induced liver sepsis exhibits the features of acute liver injury. The role of VEGF in the pathogenesis of sepsis has only recently begun to be appreciated. In the septic liver, VEGF may influence the increased permeability of hepatic microvessels. TNF-α and LPS markedly increase VEGF protein secretion and mRNA expression in macrophages of cirrhotic patients (25). Moreover, ET-1 is also a potent inducer of VEGF expression (26). In a recent study, Flt-1 agonists were shown to have therapeutic potential for preservation of organ function in certain liver disorders (27). Thus, the upregulation of Flt-1 in liver after LPS administration may be a compensatory change to preserve the hepatic function, but this is only speculatioň.
VEGF in the septic liver may cause the induction of nitric oxide (NO) production. By binding to its receptors, VEGF initiates the signaling cascades that lead to NO production (28). Administration of LPS results in the increased expression and activity of iNOS/NOS2, with a concomitant increase in NO generation (29, 30) that, if unchecked, can exacerbate septic shock and lead to cellular and tissue damage, extreme vasodilatation, loss of systemic vascular resistance, hypotension (31), and, finally, death.
In the present study, ET-1 was upregulated in the liver tissue of rats in a time-dependent manner after LPS administration. In addition, the expression of VEGF and Flt-1 in liver tissue also increased in association with the progression of sepsis. Again, LPS caused the decrease in both systolic and diastolic blood pressure in the present study. Thus, there might be a loss of balance in the complex and tight interaction and regulation of different vasoregulatory molecules (i.e., ET-1 and VEGF) in liver tissue, as well as in systemic circulation in rats, after LPS administration.
Changes in the ET-1 level in hepatic tissue in rats after LPS administration and in
control rats. The ET-1 level in hepatic tissue was determined by ELISA
(n = 10). Results are mean ± SD. *P < 0.01,
with respect to control. (A) Changes in the VEGF level in hepatic tissue in rats after LPS administration and
in control rats. The VEGF level in hepatic tissue was determined by ELISA
(n = 10). Results are mean ± SD. *P < 0.01,
with respect to control. (B) Changes in the Flt-1 level in hepatic tissue in rats
after LPS administration and in control rats. The level of expression of the gene
encoding Flt-1 was determined by RT-PCR (n = 10). Results are mean ±
SD. *P < 0.01, with respect to control.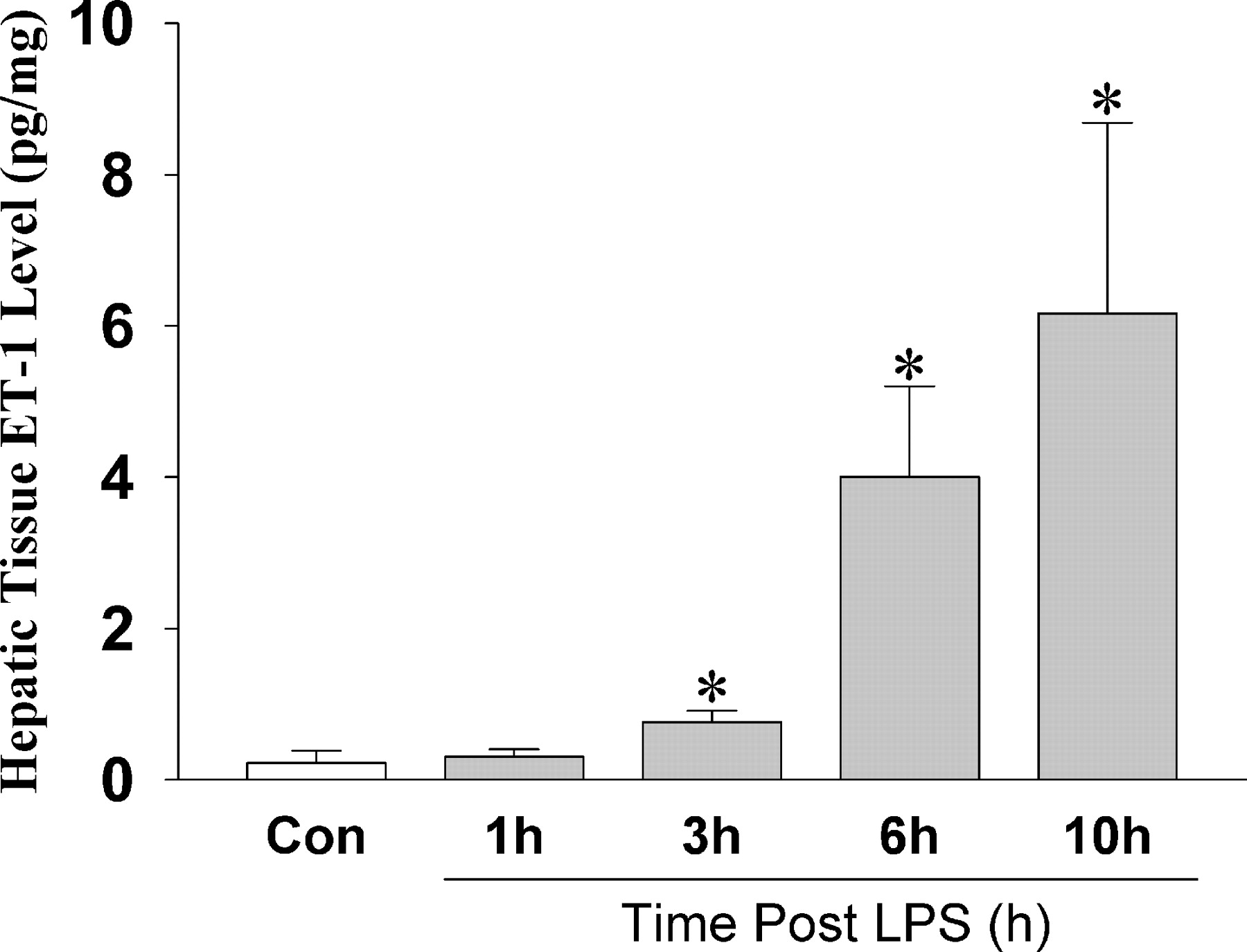
Footnotes
This work was supported by grants-in-aid for scientific research from the Ministry of Education, Science, Sports and Culture of Japan (15390077 and 15650130), and a grant from the Miyauchi project of Tsukuba Advanced Research Alliance at University of Tsukuba.