
Abstract
In this study, we investigated the effect of the methanol extract of fruits of Vitis amurensis Rupr. (Vitaceae; MEVA) on the mast cell–mediated allergy model and studied the possible mechanism of action. Mast cell–mediated allergic disease is involved in many diseases, such as asthma and sinusitis. The discovery of drugs for the treatment of allergic disease is an important subject in human health. MEVA inhibited compound 48/80–induced systemic reactions and serum histamine release in a dose-dependent manner in mice. MEVA decreased immunoglobulin E (IgE)–mediated local allergic reactions, passive cutaneous anaphylaxis. MEVA dose-dependently reduced histamine release from mast cells activated by compound 48/80 or IgE. The inhibitory effect of MEVA on histamine release was mediated by the modulation of intracellular calcium. In addition, MEVA attenuated the phorbol 12-myristate 13-acetate and calcium ionophore A23187 (PMACI)–stimulated secretion of tumor necrosis factor-α, interleukin-6 (IL-6), and IL-8 in human mast cells. The inhibitory effect of MEVA on these proinflammatory cytokines was p38 mitogen-activated protein kinase and nuclear factor-κB (NF-κB) dependent. Our findings provide evidence that MEVA inhibits mast cell–derived, immediate-type allergic reactions and involvement of proinflammatory cytokines, p38 MAPK, and NF-κB in these effects.
Introduction
Mast cells are broadly distributed throughout mammalian tissues and play a critical role in a wide variety of biologic responses. Typically, mast cells have been considered not only in the association of immediate-type hypersensitivity, but also in late reactions, like inflammatory responses (1). Immediate-type hypersensitivity is mediated by histamine released in response to the antigen cross-linking of immunoglobulin E (IgE) bound to FcεRI on the mast cells (2). After activation via the FcεRI, the mast cells start the process of degranulation, which results in the releasing of mediators, such as products of arachidonic acid metabolism and an array of inflammatory cytokines (3). Among the inflammatory substances released from the mast cells, histamine is one of the best characterized and most potent vasoactive mediators implicated in the acute phase of immediate hypersensitivity (4).
The activation of mast cells leads to the phosphorylation of tyrosine kinase and the mobilization of internal calcium. This is followed by the activation of protein kinase C, an increase of mitogen-activated protein kinases (MAPKs), nuclear factor-κB (NF-κB), and the release of inflammatory cytokines. Activated mast cells can produce histamine and a wide variety of other inflammatory mediators, such as eicosanoids, proteoglycans, proteases, and several proinflammatory and chemotactic cytokines, such as tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), IL-8, IL-4, IL-13, and transforming growth factor-β (3, 5). MAPKs and transcription factor NF-κB have important activities as mediators of cellular responses to extracellular signals. Some of the MAPKs important to mammalian cells include extracellular signal-regulated kinase (ERK), c-jun N-terminal kinase (JNK), and p38. NF-κB and p38 MAPK are thought to play an important role in the regulation of proinflammatory molecules on cellular responses, especially TNF-α, IL-6, and IL-8 (6–8).
Vitis amurensis Rupr. (Vitaceae) has been used for centuries as traditional oriental medicine (9). This crude drug contains polyphenols, mainly, catechin, procyanidin, and amurensin (10, 11). It is native to Korea, China, and Japan. It has been reported that the roots of V. amurensis have anti-inflammatory and antitumor activities (12–14). The aim of this study is to evaluate the antiallergic effect of the methanol extract of fruits of V. amurensis (MEVA) and to understand the mechanism of effect.
Materials and Methods
Animals.
The original stock of male ICR mice (20–30 g) and male Sprague-Dawley rats (200–300 g) were purchased from the Dae-Han Biolink Co. Ltd. (Daejeon, Korea). The animals were maintained in the College of Pharmacy of Woosuk University. The animals were housed 5–10 per cage in a laminar air flow room (conventional condition) maintained at a temperature of 22°C ± 2°C and with a relative humidity of 55% ± 5% throughout the study. The care and treatment of the mice were in accordance with the guidelines established by the Public Health Service Policy on the Humane Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee.
Reagents and Cell Culture.
Compound 48/80, antidinitrophenyl (anti-DNP) IgE, DNP–human serum albumin (HSA), α-minimal essential medium (α-MEM), o-phthaldialdehyde, and phorbol 12-myristate 13-acetate and calcium ionophore A23187 (PMACI) were purchased from Sigma Chemical Co. (St. Louis, MO). The rTNF-α, rIL-8, and rIL-6, and anti–TNF-α, anti–IL-8, and anti–IL-6 antibodies were purchased from R&D Systems Inc. (Minneapolis, MN). The human mast cell line (HMC-1) was grown in Iscove’s media (Life Technologies, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS) and 2 mM glutamine at 37°C in 5% CO2. Passages 4–8 of cultures were used in all experiments.
Preparation of Methanol Extract of V. amurensis (MEVA).
The fruit of V. amurensis Rupr. was collected in Imsil-Gun, South Korea, on September 25, 2006. A voucher specimen (number WSP-06–02) was deposited at the Herbarium of the College of Pharmacy of Woosuk University. V. amurensis was ground (400 g, 30 secs) at room temperature using Micro Hammer-Cutter Mill (Culatti Co., Zurich, Switzerland). The particle size was 0.5–2 mm after grinding. The plant sample (200 g) was extracted twice with 70% methanol (500 ml) at 70°C for 5 hrs in water bath. The extract was filtered through Whatman No. 1 filter paper, and the filtrate was lyophilized using a 0.45-μm syringe filter. The yield of dried extract from starting crude materials was about 20.1%. The dried extract was dissolved in saline or Tyrode buffer A (10 mM HEPES, 130 mM NaCl, 5 mM KCl, 1.4 mM CaCl2, 1 mM MgCl2, 5.6 mM glucose, and 0.1% bovine serum albumin) before use.
Compound 48/80–Induced Systemic Reaction.
Compound 48/80–induced systemic reaction was carried out as previously described (15). Briefly, the mice (n = 10 per group) were given an intraperitoneal injection of 8 mg/kg body wt of the mast cell degranulator, compound 48/80. MEVA was dissolved in saline and administered intraperitoneally at doses of 1–1000 mg/kg body wt 1 hr before the compound 48/80 injection. In the time-dependent experiment, MEVA (1000 mg/kg) was administered 5, 10, and 15 mins after compound 48/80 injection (n = 10 per group). Mortality was monitored for 1 hr after induction of anaphylactic shock.
Passive Cutaneous Anaphylaxis (PCA).
The mice were injected intradermally with 0.5 μg anti-DNP IgE. After 48 hrs, each mouse (n = 10 per group) received an injection of 1 μg DNP-HSA in PBS containing 4% Evans blue (1:4) via tail vein. MEVA (1–1000 mg/kg BW) was intraperitoneally administered 1 hr before the challenge. Thirty minutes after the challenge, the mice were killed and the dorsal skin (diameter 1 cm) was removed in order to measure the pigment area. The amount of dye was determined colorimetrically after extraction with 1 ml of 1 M KOH and 9 ml of a mixture of acetone and phosphoric acid (5:13). The intensity of the absorbent was measured at 620 nm in a spectrophotometer (UV-1201; Shimadzu, Kyoto, Japan).
Preparation of Rat Peritoneal Mast Cells (RPMCs).
Mast cells were separated from the rat peritoneal cavity cells as previously described (16). In brief, the peritoneal cells were suspended in Tyrode buffer, layered on 2 ml metrizamide (22.5 w/v), and centrifuged at 400 g for 15 mins at 4°C. The cells that remained at the buffer-metrizamide interface were aspirated and discarded; the cells in the pellet were washed and resuspended in 1 ml Tyrode buffer. Mast cell preparations were about 95% pure, as assessed by toluidine blue staining. More than 95% of the cells were viable as judged by Trypan blue exclusion.
Histamine Assay.
The histamine content was measured by the o-phthaldialdehyde spectrofluorometric procedure, as previously described (7). RPMCs were incubated with MEVA (0.001–1 mg/ml) or media alone for 10 mins at 37°C before the addition of compound 48/80 (5 μg/ml) and incubation for an additional 10 mins. The cells were separated from the released histamine by centrifugation at 400 g for 5 mins at 4°C. The fluorescent intensity was measured at emission 438 nm and excitation 353 nm using a spectrofluorometer (RF-5301 PC; Shimadzu).
Intracellular Calcium.
Fura-2/AM (2μM; Molecular Probes, Eugene, OR) was used to determine the intracellular calcium following the manufacturer’s protocol briefly described as following. RPMCs and HMC-1 cells were preincubated with Fura-2/AM for 30 mins at 37°C. After washing the dye from the cell surface, the cells were treated with MEVA for 10 mins before addition of compound 48/80 (2 μg/ml). The fluorescent intensity was recorded using a fluorescent plate reader (Molecular Devices, Sunnyvale, CA) at an excitation of 340 nm and an emission of 500 nm.
Assay of TNF-α, IL-6, and IL-8 Secretion.
TNF-α, IL-6, and IL-8 secretion was measured by modification of an enzyme-linked immunoabsorbent assay (ELISA). HMC-1 cells were cultured with α-MEM plus 10% FBS and resuspended in Tyrode buffer A. The cells were sensitized with PMA (20 nM) plus A23187 (1 μM) for 16 hrs in the absence or presence of MEVA. The ELISA was performed by coating 96-well plates with 6.25 ng/well monoclonal antibody with specificity for TNF-α, IL-6, and IL-8, respectively. Before use and between subsequent steps in the assay, the coated plates were washed twice with PBS containing 0.05% Tween-20 and twice with PBS alone. For the standard curve, rTNF-α, rIL-6, and rIL-8 were added to the serum, which was previously determined to be negative to endogenous TNF-α, IL-6, and IL-8. After exposure to the medium, the assay plates were exposed sequentially to biotinylated anti-human TNF-α, IL-6, or IL-8 and 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) tablet substrates. Optical density readings were made within 10 mins of the addition of the substrate with a 405-nm filter.
Nuclear Protein Extraction.
Preparation of nuclear extract was basically as described elsewhere (17). Briefly, after cell activation for the times indicated, cells were washed in 1 ml ice-cold PBS, centrifuged at 400 g for 5 mins, resuspended in 400 μl ice-cold hypotonic buffer (10 mM HEPES/KOH, 2 mM MgCl2, 0.1 mM EDTA, 10 mM KCl, 1 mM dithiothreitol [DTT], 0.5 mM phenylmethylsulfonyl fluoride [PMSF], pH 7.9), left on ice for 10 mins, vortexed, and centrifuged at 15,000 g for 30 secs. Pelleted nuclei were gently resuspended in 50 μl ice-cold saline buffer (50 mM HEPES/KOH, 50 mM KCl, 300 mM NaCl, 0.1 mM EDTA, 10% glycerol, 1 mM DTT, 0.5 mM PMSF, pH 7.9), left on ice for 20 mins, vortexed, and centrifuged at 15,000 g for 5 mins at 4°C. Aliquots of the supernatant that contained nuclear proteins were frozen in liquid nitrogen and stored at −70°C.
Western Blot Analyses.
HMC-1 cells were washed with PBS and resuspended in lysis buffer. Samples were electrophoresed using 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, as described elsewhere (18), and then transferred to a nitrocellulose membrane. The phosphorylation of p38 MAPK was assayed using anti–phospho-p38 MAPK antibody (Santa Cruz Biotechnology, Santa Cruz, CA). Immunodetection was done using an enhanced chemiluminescence detection kit (Amersham Pharmacia, Piscataway, NJ).
Electrophoretic Mobility Shift Assays (EM-SA).
Nuclear protein (10 μg) was incubated for 20 mins at room temperature with 20 μg bovine serum albumin, 2 μg poly(dI–dC) from Pharmacia (Uppsala, Sweden), 2 μl buffer C (20 mM HEPES/KOH, 20% glycerol, 100 mM KCl, 0.5 mM PMSF, pH 7.9), 4 μl buffer F (20% ficoll-400, 100 mM HEPES/KOH, 300 mM KCl, 10 mM DTT, 0.5 mM PMSF, pH 7.9), and 20,000 cpm of a 32P-labeled probe that encoded the κB consensus sequence (5′-CAG AGG GGA CTT TCC GAG AG-3′) in a final volume of 20 μl. DNA-protein complexes were resolved at 180 V for 4 hrs in a native 4% polyacrylamide gel, dried, and visualized (with autography using a Fuji x-ray film).
Transient Transfection and Luciferase Activity Assay.
For transient transfections, HMC-1 cells were seeded at 2 × 106 in a six-well plate 1 day before transient transfection. The expression vectors containing the NF-κB luciferase reporter construct (pNF-κB–LUC, plasmid containing NF-κB binding site; Stantagen, Grand Island, NY) were transfected with serum- and antibiotic-free Iscove’s medium containing 8 μl Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA). After 5 hrs of incubation, medium was replaced with Iscove’s medium containing 10% FBS and antibiotics. Cells were allowed to recover at 37°C for 20 hrs and subsequently were stimulated as indicated. Cell lysates were prepared and assayed for luciferase activity using Luciferase Assay System (Promega, Madison, WI), according to the manufacturer’s instructions.
Statistical Analysis.
Statistical analyses were performed using SAS statistical software (SAS Institute, Cary, NC). Treatment effects were analyzed using oneway ANOVA, followed by Duncan’s multiple range tests. P < 0.05 was used to indicate significance.
Results
Effect of MEVA on Compound 48/80–Induced Systemic Reaction.
To determine the effect of MEVA on allergic reaction, an in vivo model of a systemic reaction was used. Compound 48/80 (8 mg/kg) was used as a model of induction for a systemic fatal allergic reaction. After the intraperitoneal injection of compound 48/80, the mice were monitored for 1 hr, after which the mortality rate was determined. As shown in Table 1, injection of compound 48/80 into mice induced fatal shock in 100% of animals. When MEVA was intraperitoneally administered at a concentrations ranging from 1 to 1000 mg/kg body wt for 1 hr, the mortality with compound 48/80 was dose-dependently reduced. In addition, the mortality of mice administered with MEVA (1000 mg/kg) 5, 10, and 15 mins after compound 48/80 injection increased time dependently (Table 2). We evaluated the effect of MEVA treatment on compound 48/80–induced serum histamine release. Our results indicated that MEVA dose-dependently decreased the compound 48/80–induced serum histamine release (Fig. 1A).
Effect of MEVA on IgE-Mediated PCA.
Another way to test the anaphylactic reactions is to induce PCA. A local extravasation was induced by a local injection of IgE followed by an antigenic challenge. Intraperitoneal injection of MEVA dose-dependently inhibited PCA reaction (Fig. 1B).
Effect of MEVA on Histamine Release from RPMCs.
We next evaluated the ability of MEVA to inhibit compound 48/80–induced and IgE-mediated histamine release from RPMCs. Treatment with MEVA dose-dependently inhibited compound 48/80–induced and IgE-mediated histamine release at concentrations of 0.1–1 mg/ml (Fig. 2). The concentration of MEVA treatment used in these studies had no significant effect on the viability of RPMCs (data not shown).
Effect of MEVA on Intracellular Calcium in RPMCs and HMC–1 Cells.
Calcium movements across membranes of mast cells are critical to histamine release (19). To investigate the mechanism of MEVA on the reduction of histamine release, we assayed intracellular calcium levels. Figure 3 shows the stimulation of intra-cellular calcium when mast cells are treated with compound 48/80 (2 μg/ml). Preincubation of MEVA with cells decreased the intracellular calcium level induced by compound 48/80.
Effect of MEVA on the Secretion of Proinflammatory Cytokines from HMC-1 Cells.
TNF-α, IL-6, and IL-8 are the most important proinflammatory cytokines. Therefore, we tested the effect of MEVA on TNF-α, IL-6, and IL-8 secretion from HMC-1 cells. The HMC-1 cell line is a useful cell for studying the cytokine activation pathway (7, 20). Culture supernatants were assayed for each cytokine level by ELISA methods. Stimulation of HMC-1 cells with PMA (20 nM) and A23187 (1 μM) during 16 hrs induced the secretion of all three types of cytokines. MEVA dose-dependently inhibited the secretion of TNF-α, IL-6, and IL-8 in PMACI-stimulated HMC-1 cells (Table 3).
Effect of MEVA on Activation of p38 MAPK and NF-κB.
MAPK pathways play a crucial role in the regulation of proinflammatory molecules on cellular responses (21, 22). Previously, we documented that PMACI activates all three types of MAPKs, such as p38, JNK, and ERK, at 15–30 mins in HMC-1 cells (7, 15). To evaluate the mechanisms of effect of MEVA on the secretion of proinflammatory cytokines, we examined the effect of MEVA on the activation of MAPKs. MEVA attenuated the PMACI-induced phosphorylation of p38 MAPK (Fig. 4) but did not affect the phosphorylation of JNK and ERK (data not shown).
NF-κB is an important transcriptional regulator of inflammatory cytokines and plays a crucial role in immune and inflammatory responses. To investigate the intracellular mechanism responsible for the inhibitory effect of MEVA on the secretion of proinflammatory cytokines, we examined the effect of MEVA on NF-κB activity using EMSA and luciferase reporter assay. Cells pretreated with MEVA were subsequently stimulated by PMACI, and the effect of MEVA on binding activity of transcription factors was examined. In the presence of MEVA, PMACI-induced NF-κB/DNA binding was markedly suppressed (Fig. 4C). To confirm the inhibitory effect of MEVA on NF-κB activation, we examined the effect of MEVA on the NF-κB–dependent gene reporter assay. HMC-1 cells were transiently transfected with a NF-κB luciferase reporter construct or an empty vector. Exposure of cells to PMACI increased the luciferase activity in the cells transfected with the NF-κB luciferase reporter construct (Fig. 4D). MEVA significantly reduced the PMACI-induced luciferase activity.
Discussion
The results of this study demonstrated that MEVA has antiallergic properties. MEVA inhibited compound 48/80–induced systemic allergic reaction and histamine release from mast cells. These results indicate that mast cell–mediated, immediate-type allergic reactions are inhibited by MEVA. In addition, the MEVA-administered mice are protected from IgE-mediated PCA, which is one of the most important in vivo models of anaphylaxis in a local allergic reaction. This finding suggests that MEVA might be useful in the treatment of allergic diseases, especially skin reactions.
The intracellular calcium is the critical functional compartments in the degranulation of mast cells. Calcium movements across the membranes of mast cells represent a major target for effective antiallergic drugs, as these are essential events that link stimulation to secretion (3, 23). The transduction pathways modulating intracellular calcium are modified by ADP-rybosylates G protein-binding protein (24). Our results, which show an attenuation of intracellular calcium in mast cells with MEVA treatment, are consistent with other reports. According to these observations, we strongly suggest that decreased intracellular calcium might be involved in the inhibitory effect of MEVA on histamine release, and MEVA might have membrane-stabilizing activity through G protein.
Numerous reports established that stimulation of mast cells with compound 48/80 or IgE initiates the activation of signal transduction pathway, which leads to histamine release. Several recent studies have shown that compound 48/80 and other polybasic compounds are able—apparently directly—to activate G proteins (25). Compound 48/80 increases the permeability of the lipid bilayer membrane by causing a perturbation in the membrane. This result indicates that the increase in membrane permeability may be an essential trigger for the release of the mediator from mast cells. In this sense, antiallergic agents having a membrane-stabilizing action may be desirable. MEVA might stabilize the lipid bilayer membrane, thus preventing the perturbation being induced by compound 48/80.
The HMC-1 cell line is a useful cell for studying cytokine activation pathways (26). The spectrum of cytokines produced by HMC-1 cells with PMACI stimulation supports the well-recognized role of mast cells in immediate-type hypersensitivity. Proinflammatory cytokines, including TNF-α, IL-6, and IL-8, play a major role in triggering and sustaining the allergic inflammatory response in mast cells (27, 28). Mast cells are a principal source of TNF-α in human dermis. TNF-α has an important amplifying effect in asthmatic inflammation and potently stimulates airway epithelial cells to produce cytokines (5). It is also a potent inducer of other inflammatory cytokines, including IL-1, IL-6, IL-8, and granulocyte-macrophage colony-stimulating factor. IL-6 is produced from mast cells, and its local accumulation is associated with PCA reaction (28). IL-8 from mast cells acts on surrounding cells, such as neutrophils, T lymphocytes, and eosinophils, and plays a role in activation of inflammatory effector cells (29). These reports indicate that the reduction of proinflammatory cytokines from mast cells is one of the key indicators of reduced allergic symptom. In our present study, MEVA inhibited the secretion of TNF-α, IL-6, and IL-8 in PMACI-stimulated HMC-1 cells. This result suggests that the antiallergic effect of MEVA results from its reduction of TNF-α, IL-6, and IL-8 from mast cells.
Intracellular calcium plays an important role in the expression of inflammatory cytokines. Depletion of intra-cellular calcium blocked the IgE-induced TNF-α and IL-6 expression through the NF-κB signaling pathway in RBL-2H3 mast cells (30). Because of the reducing effect of MEVA on the intracellular calcium, we suggest that one possible pathway of the inhibitory effect of MEVA on the inflammatory cytokines is mediated by the reduction of intracellular calcium in mast cells.
The MAPK cascade is one of the important signaling pathways in immune responses (21). The expression of inflammatory cytokines is regulated by MAPKs. The exact signaling pathways among three types of MAPKs, such as p38, ERK, and JNK, are still unclear; however, p38 MAPK is thought to play an important role in regulation of inflammatory responses. Activation of p38 MAPK is essential for the expression of the proinflammatory cytokines (31, 32). Our previous report showed that PMACI simultaneously activated all three MAPKs in HMC-1 cells. Transfection of dominant-negative p38 and specific p38 MAPK inhibitor, SB 203580, decreased expression of TNF-α and IL-6 (7). In the present study, MEVA specifically inhibited the activation of p38 MAPK but not ERK or JNK. These data suggest that MEVA has the inhibitory activity on p38 MAPK activation and downstream TNF-α, IL-6, and IL-8 production.
Expression of TNF-α, IL-6, and IL-8 gene is dependent on the activation of transcription factor NF-κB (33). Activation of NF-κB requires phosphorylation and proteolytic degradation of the inhibitory protein IκBα, an endogenous inhibitor that binds to NF-κB in the cytoplasm (6). In PMACI-stimulated mast cells, MEVA inhibited DNA binding of NF-κB and NF-κB–dependent gene transcription. We also previously reported that PDTC, the potent inhibitor of NF-κB, reduced PMACI-induced production of TNF-α, IL-6, and IL-8 in HMC-1 cells (7, 17). These data demonstrate that MEVA attenuates activation of NF-κB and downstream TNF-α, IL-6, and IL-8 production.
Because we used a whole extract of V. amurensis, not a purified component, the active components that are responsible for the biologic effect are not clear at this time. The effort to identify active components from the fruits of V. amurensis in the mast cell–mediated allergic reaction is ongoing in our laboratory. In the present report we provided evidence that MEVA inhibits a model of mast cell–mediated allergic reactions and their possible mechanisms, such as p38 MAPK and NF-κB, and downstream proinflammatory cytokines. The results obtained in the present study shows that MEVA contributes to the prevention or treatment of mast cell–mediated allergic diseases.
Effect of MEVA on Compound 48/80–Induced Systemic Anaphylaxis a
Time-Dependent Effect of MEVA on Compound 48/80–Induced Systemic Anaphylaxis a
Effect of MEVA on PMACI-Stimulated Cytokine Secretion from HMC-1 Cells a
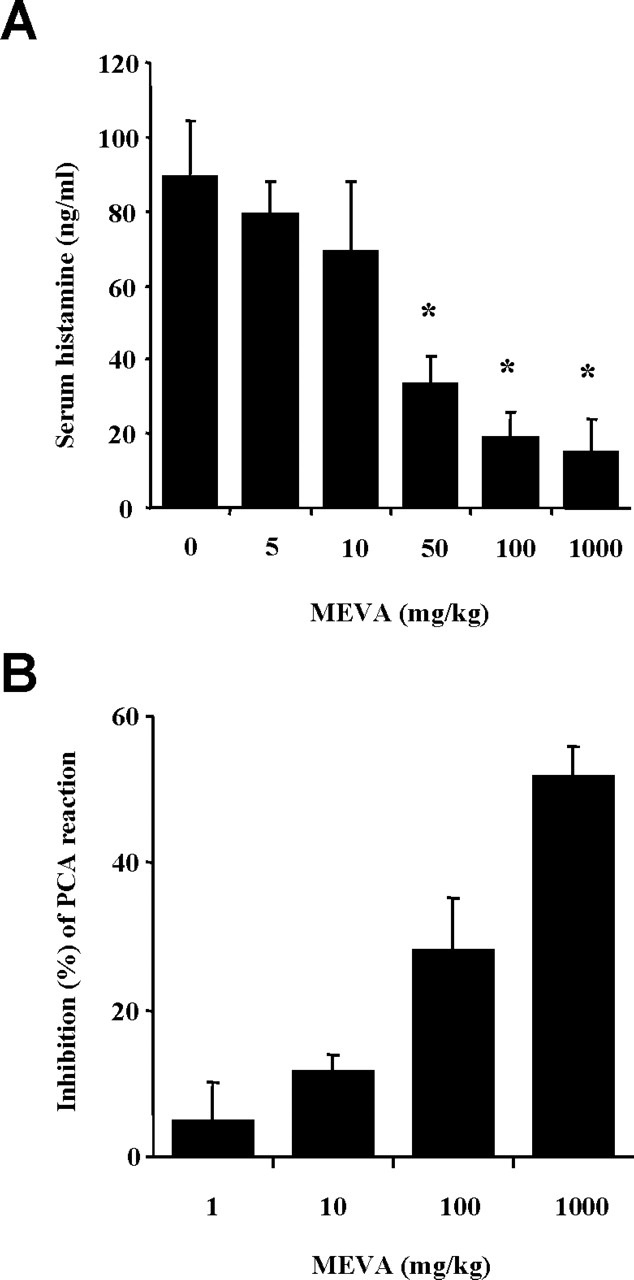
Effect of MEVA on serum histamine release and PCA reactions. (A) Groups of mice (n = 10 per group) were intra-peritoneally treated with 200 μl of saline or MEVA at various doses 1 hr before the intraperitoneal injection of the compound 48/80 (8 mg/kg). The blood was obtained from the heart of each mouse, and histamine content was measured by spectrofluorometry. (B) MEVA was administered 1 hr prior to the challenge with antigen. Each amount of dye was extracted as described in “Materials and Methods” and measured by spectrophotometry. *P < 0.05, statistically significant.
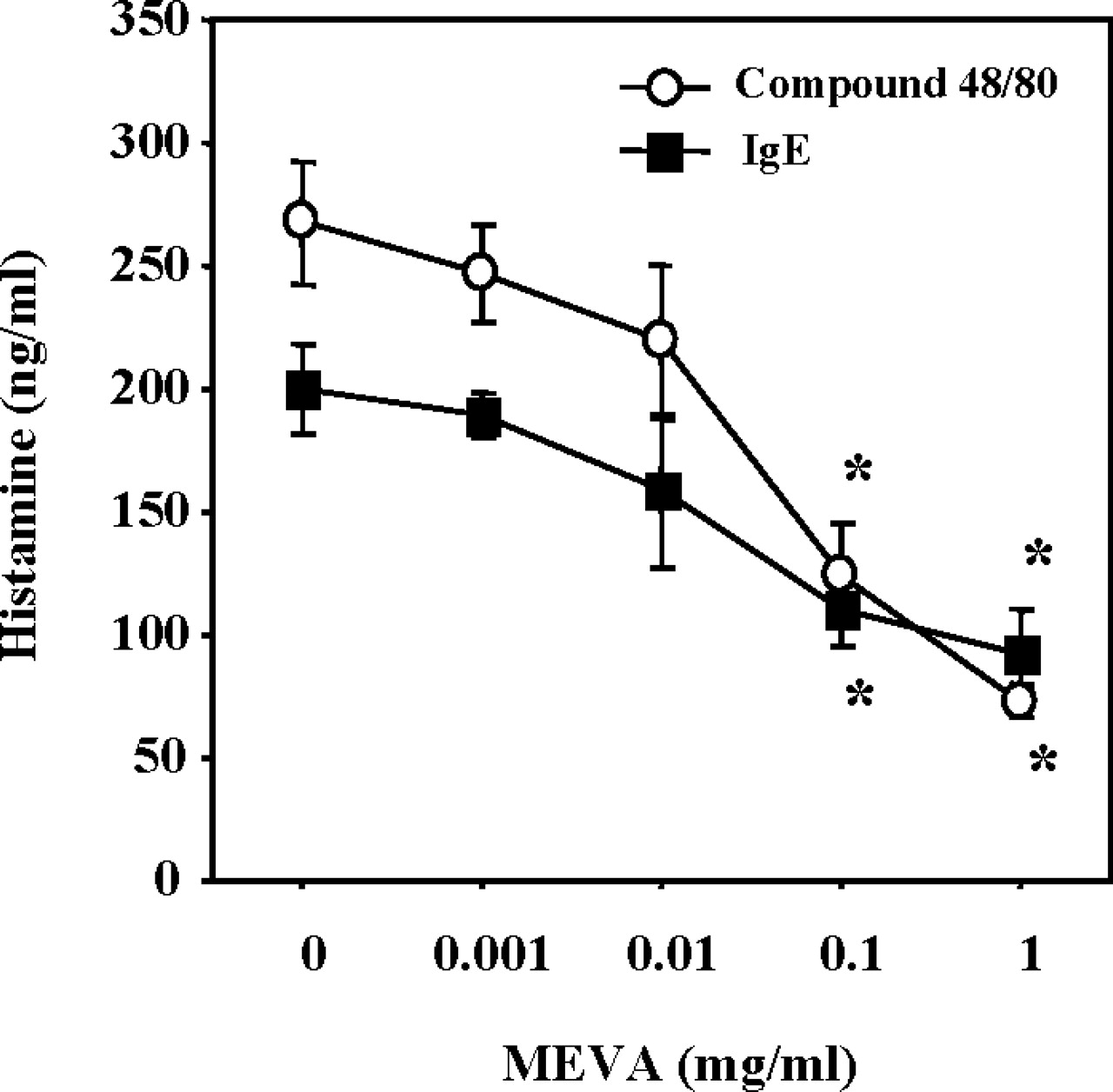
Effect of MEVA on compound 48/80–induced or IgE-mediated histamine release from RPMCs. The cells (2 × 105 cells/ml) were preincubated with MEVA at 37°C for 10 mins prior to incubation with either compound 48/80 (naive RPMCs) or DNP-HSA (anti-DNP IgE-sensitized RPMCs). Each value represents the mean ± SEM of three independent experiments. *P < 0.05 (significantly different from the compound 48/80 or IgE value).

Effect of MEVA on intracellular calcium in mast cells. RPMCs and HMC-1 cells were stained with Fura-2

Effect of MEVA on activation of p38 MAPKs and NF-κB in HMC-1 cells. After pretreatment of MEVA for 30 mins, HMC-1 cells were stimulated by PMA (20 nM) and A23187 (1 μM) for 15 mins for p38 MAPK activation or 2 hrs for NF-κB activation. (A) Phosphorylation of p38 MAPKs was analyzed by Western blot. (B) Densitometric analysis of p38 MAPK. (C) Inhibition of NF-κB binding activity by MEVA was analyzed by EMSA. (D) For NF-κB luciferase assay, cells were transiently transfected with the NF-κB luciferase reporter construct or empty vector. NF-κB–dependent transcriptional activity was determined by luciferase activity assay. Each value represents the mean ± SEM of three independent experiments. *P < 0.05, statistically significant. PMACI: PMA and A23187.
Footnotes
This work was supported by the Korea Science and Engineering Foundation (KOSEF) grant funded by the Korean government (MOST; M10646020001-06N4602–00110), and by the Korea Research Foundation Grant funded by the Korean government (MOEHRD, Basic Research Promotion Fund; KRF-2006–003-E00427).