
Abstract
In myelinated nerve fibers, action potential initiation and propagation requires that voltage-gated ion channels be clustered at high density in the axon initial segments and nodes of Ranvier. The molecular organization of these subdomains depends on specialized cytoskeletal and scaffolding proteins such as spectrins, ankyrins, and 4.1 proteins. These cytoskeletal proteins are considered to be important for 1) formation, localization, and maintenance of specific integral membrane protein complexes, 2) a barrier restricting the diffusion of both cytoplasmic and membrane proteins to distinct regions or compartments of the cell, and 3) stabilization of axonal membrane integrity. Increased insights into the role of the cytoskeleton could provide important clues about the pathophysiology of various neurological disorders.
Introduction
In nervous systems, action potentials (APs) must be conducted rapidly and efficiently over long distances. In vertebrates, this has been achieved through the development of myelin, a multilammelar structure that ensheathes axons typically along their entire length. Myelin increases membrane resistance and decreases membrane capacitance, thereby conserving ionic charge as the axolemma is depolarized during AP propagation. Schwann cells in the peripheral nervous system and oligodendrocytes in the central nervous system are the glial cells that produce myelin. Importantly, these glial cells also play active roles in regulating neuronal properties including axonal ion channel expression and localization. Myelinated axons are divided into several polarized domains, that include (but are not limited to) the axon initial segment (AIS), the node of Ranvier, and paranodal junctions where the myelin sheath attaches to the axon (Fig. 1). The AIS is a short axonal segment located adjacent to the cell body, whereas nodes are regularly spaced gaps between each myelin segment (approximately 1 μm long). Both nodes and the AIS are characterized by an “electron-dense” membrane undercoating that is thought to reflect the high density of voltage-gated Na+ and K+ (Nav and Kv, respectively) channels, cell adhesion molecules (CAMs), and cytoskeletal and scaffolding proteins found at these sites (reviewed in references (1, 2)). Functionally, Nav channels are the most important and significant component since they are responsible for generating the AP. Nav channels are selectively permeable to Na+ and open when the membrane is depolarized. The AIS is important for AP initiation (3, 4), whereas the node is required for saltatory AP propagation along the axons. Both AP initiation and propagation require the precise distribution of Nav channels accumulated at high density in these subdomains. For example, disruption of nodal Nav channel clusters by autoimmune processes causes profound limb weakness (5). How are Nav channels selectively localized and maintained in high densities at the AIS and the nodes? Recent data indicate that the submembranous cytoskeleton anchors Nav channels and other transmembrane proteins, and plays important roles for both formation and maintenance of the molecular organization at the AIS and nodes. This review describes the cytoskeletal protein complexes at polarized subdomains in myelinated axons and focuses on their possible roles.
Spectrin and Ankyrin-Based Cytoskeletons Associated with Nav Channel Clusters
Several classes of cytoskeletal proteins, including spectrins, ankyrins, and 4.1 proteins, are highly enriched at the AIS, nodes, or paranodes (1, 2, 6). Spectrins are a family of extended, flexible molecules comprising α and β subunits (reviewed in reference (7) in detail). These subunits are associated laterally to form anti-parallel heterodimers of α spectrin and β spectrin. Then, the heterodimers are assembled head-to-head to form heterotetramers. β spectrins contain NH2-terminal actin-binding domains, and the spectrin tetramers are bound to one another indirectly via short actin filaments to form a flexible meshwork. β Spectrins have triple helical repeating units, and the 15th spectrin repeat interacts with ankyrins. Ankyrins are large scaffolding proteins that link the spectrin-based membrane skeleton to various membrane proteins, including Nav channels, members of the L1 CAM family, and various ion exchangers and pumps (7). The ankyrin gene family of mammals includes three members: ankyrinR, ankyrinB, and ankyrinG. The protein 4.1 associates with β spectrin at a site in the NH2-terminal region (7). Protein 4.1 also interacts with F-actin, and stabilizes spectrin-actin complexes. Spectrins, ankyrins, and 4.1 proteins were first discovered as components of the erythrocyte cytoskeleton. A major function of the spectrin-based skeleton is to mechanically support the membrane bilayer permitting erythrocytes to survive as they are deformed during circulation through small capillaries. The spectrin-based membrane skeleton is found in most cells, including neurons. However, their organization, protein interactions, and functions are considerably more diverse than in erythrocytes. At the AIS, nodes of Ranvier, and paranodal junctions, the spectrin and ankyrin-based cytoskeletons are considered to be important for: 1) formation, localization, and maintenance of specific integral membrane protein complexes, 2) a barrier restricting the diffusion of both cytoplasmic and membrane proteins to distinct regions or compartments of the cell, and 3) stabilization of axonal membrane integrity.
Formation and Stabilization of Nav Channel Clusters at the AIS
The AIS is a unique subdomain of the neuron that contributes to AP initiation (3, 4). The molecular composition of the AIS includes Nav channels, Kv channels (specifically KCNQ2 and KCNQ3), NrCAM, the 186-kDa isoform of neurofascin (NF), and the scaffolding and cytoskeletal proteins, ankyrinG and βIV spectrin (1, 2) (Fig. 1A, D). Further, specific ankyrinG isoforms of 480 and 270 kD are found at the AIS and link Nav and KCNQ2/3 Kv channels (these have a common intracellular ankyrinG binding motif) to the underlying spectrin-based cytoskeleton (8–13). NF186 has an FIGQY ankyrinG-binding motif that is conserved among all members of the L1 CAMs (14). βIV Spectrin is one of five β spectrin family members in mammals. Two different splice variants of βIV spectrin have been shown to be present at the AIS: a full-length Σ1 variant, and a shorter Σ6 splice variant lacking the actin-binding domain and the first 10 spectrin repeats (Σ6 corresponds to approximately the last half of the Σ1 splice variant) (15–17).
Although the precise mechanism whereby the AIS is formed is not fully understood, it is clear that the AIS is intrinsically organized by neurons (18, 19). This is in stark contrast to other polarized domains of axons which depend on glial contact. Hippocampal neurons cultured without myelinating glia develop a well-defined AIS characterized by high densities of Nav channels. Cytoskeletal proteins have now been shown to play essential roles in the molecular assembly of the AIS. For example, ankyrinG, along with βIV spectrin, is present at developing Purkinje neuron initial segments before Nav1.6, NF186, or NrCAM can be detected, suggesting that the AIS cytoskeleton forms before these other membrane proteins are recruited to these sites (20). The clustering of AIS membrane proteins is severely impaired in ankyrinG or βIV spectrin mutant mice, demonstrating that ankyrinG and βIV spectrin both contribute to formation and maintenance of these integral membrane clusters (13, 16, 20–22). Compared to βIV spectrin, ankyrinG appears to be central to AIS formation. The interactions of the cytoplasmic domain of NF186 with ankyrinG are required for localization to the AIS (19). AIS localization of βIV spectrin depends on binding to ankyrinG through the βIV spectrin repeat 15 (23). βIV spectrin was not clustered at the AIS of ankyrinG deficient neurons (16, 20), and silencing of ankyrinG expression blocks clustering of Nav channels, NF186, or βIV spectrin both in vitro and in vivo (24). In contrast, in qv <it>3J </it>“quivering” mice whose βIV spectrins have a truncated proline-rich “specific” domain and lack the pleckstrin homology domain (25), ankyrinG is properly localized at the AIS (26). Furthermore, βIV spectrin by itself cannot recruit other interacting proteins to the AIS. However, loss of βIV spectrin (16, 27) or Nav channels (28) has also been reported to disrupt the AIS. One possible explanation for this difference is that βIV spectrin is required for AIS stability rather than formation (24), an idea that is consistent with βIV spectrin’s role at nodes of Ranvier (26). Taken together, the available data suggest that of the known AIS proteins, ankyrinG is the main organizer of the AIS. To determine how the AIS is specified, it will be necessary to determine how ankyrinG is recruited and localized specifically at the AIS rather than in proximal regions of dendrites.
Nodes of Ranvier
Nodes of Ranvier are short gaps in the myelin sheath and have nearly every protein component in common with the AIS (1, 2) (Fig. 1B–D). Similarly, central and peripheral nervous system nodes have a common cytoskeletal organization. In contrast to the AIS, the formation of nodes requires extrinsic interactions supplied by myelinating glia (18). During early developmental myelination, NF186 and NrCAM are the first axonal molecules clustered at nascent nodes (29, 30). In the peripheral nervous system, these two CAMs are clustered in the axonal membrane through interactions with glial derived extracellular matrix molecules like gliomedin, secreted from Schwann cells (31). These CAMs are thought to act as pioneer molecules, serving as nucleation sites for recruitment of ankyrinG (18–20, 29). Similar to the AIS, ankyrinG plays a central role in organizing a nodal protein complex including Nav channels, L1-family CAMs, and βIV spectrin. AnkyrinG is detected before clustering of Nav channels or βIV spectrin (32, 33). Knockdown of ankyrinG in myelinated dorsal root ganglion cells results in failure to cluster Nav channels at nodes (19). The ankyrinG-binding spectrin repeat 15 is required for localization of βIV spectrin to nodes (23). Thus, as at the AIS, ankyrinG appears to be a critical component for the molecular assembly of nodes of Ranvier.
Besides their role in the initial assembly of nodes, what might be the function of the specialized nodal cytoskeleton throughout life? The interactions between ankyrinG and membrane proteins may promote their stable and restricted localization at nodes. Consistent with this idea, neurons transfected with an NF186 construct lacking the ankyrin-binding domain show accelerated protein turnover of the introduced NF186 (19). Compared to the considerable numbers of developmental studies, very little is known about the mechanisms responsible for maintenance of polarized subdomains in myelinated axons.
Subdomains Near the Nodes of Ranvier
The paranodes flank both sides of the nodes of Ranvier (Fig. 1B–D). At these sites, myelin and axons form septate-like junctions which act as a diffusion barrier that contributes to formation and maintenance of nodal Nav channel complexes. The paranodal domain is characterized by a tripartite CAM complex, consisting of axonal contactin-associated protein (Caspr) and contactin, and the glial 155-kDa isoform of NF that mediates the attachment of the myelin sheath to the axon (1, 2). The ultrastructural studies of paranodal junctions suggested that paranodal multiprotein complexes are connected to the cytoskeleton (34). Consistent with this idea, a specialized paranodal cytoskeleton has been identified that consists of αII spectrin, βII spectrin, ankyrinB and protein 4.1B (35–40). In a mutant zebrafish without the expression of αII spectrin, nodal Nav channel clusters are disrupted (40). Examination of Caspr-null mice (41) demonstrated that in the absence of a paranodal junction, ankyrinB fails to accumulate at these sites, suggesting that neuronglia interactions are important for organizing the paranodal cytoskeleton (38). Consistent with this idea, mice lacking paranodal junctions undergo severe axon degeneration in the cerebellum, suggesting that neuronglia interactions are required for axon integrity (39). Protein 4.1B forms a ternary complex with actin and spectrin, and provides a link between the transmembrane Caspr and the axonal cytoskeleton (37). During development, protein 4.1B appears at paranodes after Caspr, and helps to stabilize the Caspr-contactin complex.
As another polarized membrane domain with a specialized cytoskeleton, the juxtaparanodes begin at the innermost axo-glial junction of the paranodes (Fig. 1D). This domain extends into the internodes for 5–15 μm, and is characterized by high densities of Kv1 channels (1, 2). Although the role of juxtaparanodal Kv1 channels is still unknown, they have been proposed to act as an active damper of re-entrant excitation and to help in maintaining the internodal resting potential (42, 43). These Kv1 channels colocalize with and form a macromolecular complex with the CAMs Caspr2 and Tag1 (44, 45). This protein complex is essential for Kv1 channel localization at the juxtaparanodes. Importantly, Caspr2 interacts with the scaffolding protein 4.1B (37). Similar to the paranodes, protein 4.1B immunoreactivity accumulates progressively at the juxtaparanodes during development, following the concentration of Caspr2. The protein 4.1B may anchor the juxtaparanodal Caspr2-Kv1 channel complex to the cytoskeleton.
The Polarized Cytoskeleton as a Diffusion Barrier
Neurons can be divided into two molecularly and functionally distinct compartments: an axonal domain and a somatodendritic domain with numerous tapered and highly branched dendrites. Certain specific molecules, vesicles, and organelles are only found in axons, and others are only found in dendrites. Such polarity may be maintained by a diffusion barrier that impedes mixing of components. Although not yet explicitly proven, the AIS is thought to act as a diffusion barrier that maintains the asymmetric distribution of membrane-associated molecules between axonal and somatodendritic compartments and its functional polarity (46, 47). Importantly, the restriction of proteins to distinct domains likely depends on the underlying cytoskeleton, since protein mobility was increased after pharmacological disruption of actin filaments (46). Surprisingly, the barrier function of the AIS extends from transmembrane proteins to lipids. Using single molecule tracking methods, Nakada et al. (48) demonstrated that the diffusion of individual lipids is restricted in the plane of the AIS membrane. Furthermore, the reduction in lateral mobility of phospholipids at the AIS correlated in time and space with expression of ankyrinG (48), supporting the idea that the cytoskeleton is essential for the diffusion barrier (46). How might lipids be restricted in their mobility? βIV spectrin has a pleckstrin homology lipid-binding domain, and this may contribute to the specific lipid composition of the AIS and nodes. Mutant mice lacking the βIV spectrin pleckstrin homology domain (25) have disrupted nodes of Ranvier (26). Furthermore, βIV spectrin was recently reported to be an essential component of the AIS barrier that selectively blocks mobility of the axonal molecule L1CAM both in vitro and in vivo, probably through direct interactions between ankyrinG and L1CAM (27). Thus, the spectrin and ankyrin-based cytoskeleton at the AIS is important for prevention of the intermixing of freely mobile molecules in the adjacent territories as well as for the immobilization of membrane proteins required for AP initiation.
Possible Role of Cytoskeleton for Axonal Integrity at the AIS and Nodes
One important role of cytoskeletal proteins is to physically stabilize the membrane organization and integrity. For example, mutations in the erythrocyte-specific αI-and βI-spectrin genes make the erythrocyte membrane fragile, resulting in elliptocytosis and spherocytosis, and consequently hemolytic anemia (49). Thus, cytoskeletal proteins are predicted to be important for the stabilization of axonal membrane integrity at polarized subdomains. Consistent with this idea, the nodal axonal membrane abnormally protrudes in mutant mice lacking βIVΣ1 spectrin (17, 22), and in qv <it>3J </it>“quivering” mice (26). Similarly, AIS components appeared to be disrupted in mutant mice lacking βIVΣ1 spectrin (17). These findings suggest that βIV spectrin is essential for axonal membrane stability at both the AIS and the nodes.
Spectrins may also have a role in maintaining the elasticity of axons. In the uncoordinated phenotype (unc-70) of Caenorhabditis elegans with dominantly inherited mutations in a β spectrin homologue, the axons spontaneously break by the acute strain generated during movement (50). Indeed, breakage can be prevented by paralyzing unc-70 mutant animals. In the vertebrate peripheral nervous system, axons are exposed to strains generated by length changes during movement. It seems likely that the nodes of Ranvier are vulnerable to movement-induced strain, because the nodes are not covered by a protective myelin sheath. The spectrin-based cytoskeleton may provide elasticity to the nodal axon to prevent mechanical injury. Further studies are required to elucidate how cytoskeletons contribute to prevent mechanical injury at polarized subdomains.
Cytoskeletal proteins may also have an important role for vesicular transport through the axons. Cytoplasmic vesicles are often accumulated around the nodal region in βIV spectrin mutant mice, although the nature of these vesicles is unknown (17, 26). Similar findings are reported in different mutant mice with disrupted paranodal junctions. In mice deficient in either the axonal CAM Caspr (51) or the glial ceramide galactosyltransferase (52), cerebellar Purkinje neurons showed large swellings accompanied by cytoskeletal disorganization at paranodal regions and axonal degeneration (39). Thus, disrupted paranodal junctions may cause damage to the paranodal cytoskeleton, and consequently result in axonal degeneration. These findings suggest that the link between paranodal axonglial junctions and the axonal cytoskeleton is important for axonal integrity and transport (53).
Finally, cytoskeletal proteins may regulate neuronal excitability by directly modifying gating of Nav channels. Persistent Na+ current was observed in tsA201 cells with Nav 1.6, which are predominantly expressed at AIS and nodes, whereas the current was reduced when ankyrinG was coexpressed (54). The membrane-binding domain of ankyrinG appeared to be critical for reduction of Na+ current. It is not known whether a similar phenomenon occurs in neurons.
Involvement of Cytoskeletal Proteins in Neurological Diseases
Some neurological diseases have been reported to be associated with disruption of the spectrin-based cytoskeleton. For example, mutations in human βIII spectrin, a 2,390-amino acid protein highly expressed in Purkinje cells, cause spinocerebellar ataxia type 5 in an 11-generation American kindred descended from President Abraham Lincoln’s grandparents and two additional families (55). Spinocerebellar ataxia type 5 is characterized by cerebellar cortical atrophy, Purkinje cell loss, and thinning of the molecular layer. Furthermore, cytoskeletal proteins can be target molecules of autoantibodies in paraneoplastic syndromes. In a case of breast cancer and paraneoplastic lower motor neuron syndrome, serum autoantibodies were found directed against AIS and nodes of Ranvier of myelinated axons (56). The major targets of the autoantibodies were βIVΣ1 spectrin and βIV spectrin 140, two isoforms of the βIV spectrin gene (57). In addition, disruption of the nodal cytoskeleton may partly contribute to loss of Nav channel clusters in a postinfectious autoimmune polyneuropathy. For example, autoantibodies to GM1 ganglioside specifically attack the nodes of Ranvier in peripheral motor nerve fibers, and cause the axonal form of Guillain-Barre′ syndrome (5). In the animal model of the disease, clustering of nodal molecules including βIV spectrin as well as Nav channels was disrupted via a complement-mediated immune pathway. The presence of these diseases related to disrupted spectrins underscores the importance of the cytoskeleton in the vertebrate nervous systems.
The role of the cytoskeleton at nodes or paranodes may provide important clues about the pathophysiology of demyelinating disorders. Multiple sclerosis (MS) is a chronic inflammatory demyelinating disease of the central nervous system. Despite the primary demyelinating nature of the disease, axon loss is now recognized as an important component of MS pathology that causes irreversible neurologic disability (58). The mechanisms of axonal loss in MS are still unknown. One of the possible explanations is that damaged paranodal axonglial interactions due to demyelination may result in disruption of the cytoskeleton and consequently axonal degeneration. Axonglia junctions are essential for the organization of the nodal and paranodal axonal cytoskeleton or axonal transport as well as the organization of molecular domains in the myelinated axons (53). In addition, a recent report suggested that autoantibodies to NF may cause axonal injury via a complement-mediated immune pathway and exacerbate the disease process of MS (59). Human pathology revealed disrupted clusters of nodal Nav channels or NF186, and paranodal Caspr or NF155 in MS lesions (60–63). Similar to the autoimmune nodal disruption in peripheral motor nerves (5), the anti-NF antibodies may disrupt the nodal cytoskeleton including βIV spectrin and cause axonal injury in MS. Efforts to better understand the roles of the spectrin and ankyrin-based cytoskeletons will increase insight into the mechanisms of neurological diseases.
Cytoskeleton in Glial Cells
As reviewed above, the interactions between neurons and glial cells are important for the formation and maintenance of polarized subdomains in myelinated nerve fibers. Furthermore, the integrity of the myelin influences the neuronal functions. For example, both myelin length and thickness determines nerve conduction velocity (64). Like neurons, myelinating glial cells are also highly polarized. However, very little is known about the molecular mechanisms regulating glial cell polarity, or the role of a submembranous cytoskeleton. F-actin and spectrin are both present beneath all Schwann cell surface membranes, but are excluded from compact myelin (65). Protein 4.1G has been reported to be expressed in Schwann cells and enriched at paranodes (66). Whether protein 4.1G contributes to formation or maintenance of paranodal structures is unknown. No ankyrin has been reported to be expressed in myelinated glial cells. Which cytoskeletal proteins are expressed in glial cells and how are they organized? What roles do glial cytoskeletons play in the initial formation or the maintenance of myelin? And finally, how do cytoskeletons contribute to the interactions between glial cells and neurons? As with neurons, we speculate that future studies will reveal an essential role played by the cytoskeleton in regulating the formation of polarized membrane domains in myelinating glial cells.
(A) βIV spectrin (red) is highly accumulated at the AIS in cultured rat hippocampal neurons. The somatodentritic domain is characterized by the staining of microtubule-associated protein 2 (MAP2; green). Scale bar = 20 μm. (B) In rat sciatic nerve, βIV spectrin (green) is highly accumulated at the node of Ranvier, whereas βII spectrin (red) is localized to paranodes flanking the node. The axon runs horizontally. Scale bar = 5 μm. (C) AnkyrinB (red) is also highly expressed at paranodal regions in rat sciatic nerve. Nav1.6 (green) is localized to the node. The axon runs horizontally. Scale bar = 5 μm. (D) Schematic presentation showing the molecular organization (ion channels, CAMs, and cytoskeletal proteins) at the specific subdomains of myelinated axons, AIS, node of Ranvier, paranode, and juxtaparanode. A color figure is available in the online version of the article.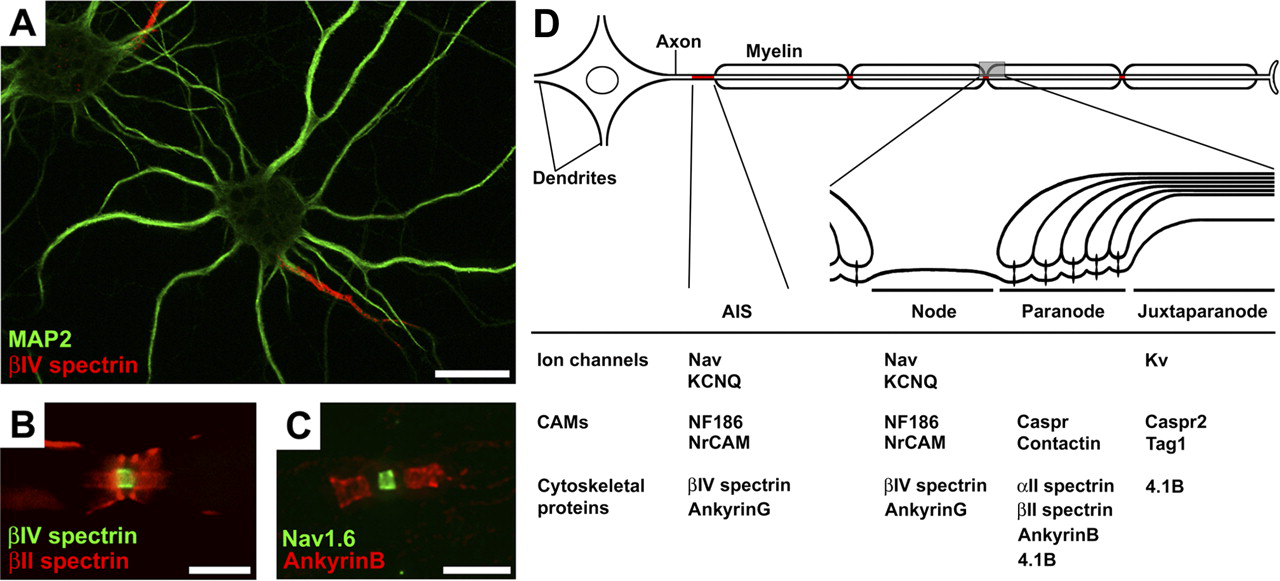
Footnotes
This work was supported by NIH grant NS044916 and the Dr. Miriam and Sheldon G. Adelson Medical Research Foundation.