
Abstract
Oxygen tension can provide an important determinant for differentiation and development of many cells and tissues. Genetic regulation of hemato-endothelial commitment is known to respond to oxygen deprivation via stimulation of hypoxia inducible factors (HIFs). Here, we use a closed bioreactor system to monitor and control the dissolved oxygen during differentiation of human embryonic stem cells (hESCs) via formation of embryoid bodies (hEBs). Exposing hESC-derived EBs to ambient oxygen at or below 5% results in stabilization of HIF-1α and increased transcription of hypoxic responsive genes. Interestingly, we find that rather than HIF-1α expression being stable over prolonged (7–16 days) culture in hypoxic conditions, HIF-1α expression peaks after approximately 48 hours of hypoxic exposure, and then declines to near undetectable levels, despite constant hypoxic exposure. This transient stabilization of HIF-1α during hESC-derived EB culture is demonstrated for four distinct stages of differentiation. Furthermore, we demonstrate hEB cell expansion is slowed by hypoxic exposure, with increased apoptosis. However, hEB cell proliferation returns to normal rates upon return to normoxic conditions. Therefore, although hypoxia effectively stimulates hypoxic responsive genes, this single variable was not sufficient to improve development of hemato-endothelial cells from hESCs.
Introduction
The formation of blood and endothelial cells is intimately intertwined during mammalian embryogenesis. Hematopoietic development first arises in yolk sac blood islands consisting of clusters of primitive erythroblasts surrounded by maturing endothelial cells (1–3). A second wave of hematopoietic cell production takes place within the aorta-gonads-mesonephros (AGM) region of the embryo proper. Here hematopoietic cells arise either very near or directly from vascular endothelial cells of the dorsal aorta (4–6). The onset of these hematopoietic and angiogenic events occurs during embryogenesis as the diffusion of oxygen and nutrients becomes limiting due to the growing size of the embryo and the formation of dense organ structures (7). The formation of the cardiovascular system allows for sufficient oxygen and nutrient delivery throughout the developing embryo to ensure proper development. The oxygen gradient formed across the growing embryo regulates various genes required for the patterning and differentiation of the cardiovascular system including flt-1, flk-1, vegf, and epo (8). In vitro, a decrease in oxygen in the culture environment amplifies the expression of these genes in murine embryos (9).
Hypoxia inducible factors (HIFs) are global regulators of this oxygen sensing response and are activated in regions of decreased oxygen concentrations (10). HIF proteins are members of the basic helix-loop-helix/Per-ARNT-Sim (bHLH-PAS) family of proteins and are known to regulate many biological processes, including angiogenesis, erythropoiesis, oxygen homeostasis, circadian rhythms, and glycolysis. HIF is a heterodimer consisting of two subunits: the inducibly expressed HIFα and the constitutively expressed HIFβ (also termed aryl hydrocarbon receptor nuclear translocator, ARNT). Although HIF-1α is expressed in most mammalian cells, HIF-2α (EPAS) and HIF-3α (11) are more restricted in their pattern of expression. Interestingly, of these HIFα subunits, HIF-2α has been shown to uniquely regulate the expression of pluripotent embryonic stem cell marker, oct-4 (12), and cultivation in reduced oxygen has been implicated in maintaining the pluripotency of hESCs in maintenance conditions (13).
HIF-1α expression and activity is post-translationally regulated. Under normoxic conditions, HIF-1α is subject to rapid ubiquitination and proteasomal degradation with a half-life of less than 5 minutes (7, 14, 15). Under hypoxic conditions, however, HIF-1α subunits are stabilized, accumulated in the cell cytoplasm, and translocated to the nucleus, where they dimerize with HIFβ subunits (10). HIF heterodimers form transcriptional complexes that induce gene expression by binding to a 50 base pair hypoxic responsive element (HRE) (16). Over 70 genes have been identified to be responsive to hypoxic conditions, including glycolytic genes, glut-1 and aldolase, and genes implicated in hematopoietic and endothelial cell development, flt-1, flk-1, epo, vegf, and others (10).
Knockout mutations of hif-1α (17–19), hif-2α (20, 21), and arnt (22–24) lead to gestational lethality of murine embryos. Phenotypic manifestations of these knockouts include defects in placental and yolk sac formation as well as reduction in vasculature, blood cell, and heart formation. These findings suggest that proper responses to oxygen sensing are vital for proper cardiovascular formation and suggest that oxygen tension is likely an important determinant of hematopoietic and endothelial cell production.
In vitro cultivation of human cord blood and bone marrow cells in reduced oxygen lead to increased frequency and number of hematopoietic progenitor cells (25) and enhanced multilineage repopulating capabilities in severe combined immunodeficient mice, a measure of putative hematopoietic stem cells (26, 27). Similarly, studies utilizing mouse embryonic stem cells (mESCs) cultivated under hypoxic conditions have been shown to promote the commitment to mesoderm and generation of hemato-endothelial progenitor cells (8, 23, 28, 29).
Although these reports have improved our understanding of the role of oxygen in mammalian hematopoietic development, human embryonic stem cells (hESCs) are now commonly used for analysis of the earliest stages of human-specific blood and endothelial cell development (30–32). Previous studies by our group and others demonstrate two waves of hematopoietic development from hESCs (33–38). A primary wave of CD34+ cells co-expressing CD31 likely represents early hematopoietic precursor cells with a common mesodermal/hemato-endothelial potential. A second wave of CD34+ cells co-expressing CD45 cells likely represents more mature hematopoietic progenitors (36). The use of reduced oxygen environments to support differentiation of hESCs toward mature hematopoietic and endothelial cell lineages has not been clearly investigated.
Here we more closely examine the effect of hypoxia on the culture, transcriptional response, and differentiation of hESCs-derived cell aggregates termed embryoid bodies (hEBs). This method of differentiation has been proposed as a means to produce clinically relevant numbers of therapeutic cells, citing ease in scalability and amenity to stirred and environmentally controlled conditions (39, 40). Better characterization of hESC differentiation remains vital to realize future clinical applications of these novel stem cell–based technologies. A bioreactor culture system is utilized to precisely monitor and control the dissolved oxygen concentration during hEB differentiation. We define an oxygen concentration under which a hypoxic response can be elicited from hEBs. Interestingly, the accumulation of HIF-1α and increased transcription of downstream targets are transient events, despite constant exposure to hypoxic conditions. Additionally, this transient behavior of HIF-1α and stimulation of target genes can be induced by hypoxic exposure during any stage of hEB differentiation.
Materials and Methods
Human Embryonic Stem Cell Culture.
hESCs, line H9, (obtained from Wicell, Madison, WI) were maintained as undifferentiated cells as previously described (33, 39, 41). Colonies of hESCs were passed every 5 to 7 days as needed. Whole or partial colonies were treated with 1 mg/mL collagenase IV (Invitrogen) in basal media for 5 minutes to remove from MEF feeder layers. hESC were routinely characterized and found to have a normal karyotype.
Human Embryoid Body Formation, Culture, and Hematopoietic Differentiation.
To promote hematopoietic differentiation hEBs were formed as previously described (33, 39, 41). A complete media change occurred after the first 24 hours in order to remove debris. Otherwise, half-media changes occurred every 3 to 4 days as needed. hEBs were cultured under normoxic conditions in a humidified Forma incubator maintained at 37° C, 5% carbon dioxide. Cell counting was performed following 10 minute digestion in 0.05% trypsin supplemented with 2% (v/v) chicken serum (Sigma), and viable cells were identified by 0.4% trypan blue exclusion.
Bioreactor Culture.
A 1-L glass reactor was equipped with silicon tubing to increase oxygen mass transfer and probes for pH, temperature, and dissolved oxygen (all from Cole Parmer). The bioreactor was situated inside a cell culture hood on top of a magnetic stirrer (Bellco) set at 80 rpm. The gas inlets consisted of separate lines for nitrogen, oxygen, and carbon dioxide, mixed together in various proportions through mass flow controllers (Brooks Instruments), and fed into the reactor at a rate of 50 mL/min. Set points of 7.2 for pH and 5% and 25% saturation for dissolved oxygen were maintained by a PID controller as part of LabVIEW software (National Instruments). The oxygen probe was calibrated by flushing the reactor with 100% air (100% saturation) and 100% nitrogen (0% saturation) and was set after the dissolved oxygen measurement had stabilized for > 10 min. Oxygen uptake rate was calculated by a stationary liquid phase balance (42). The mass transfer coefficient, kLa, for this calculation (4.7/hr) was determined by the down slope method at the start of the culture, in the absence of cells, and with the head space devoid of oxygen. kLa was assumed to remain constant throughout the culture.
Modular Hypoxic Cultures.
Differentiating EBs were exposed to hypoxic conditions by culturing in a sealed 6.2-L modular incubator (Billups Rothenberg Inc., Del Mar, CA, MIC-101) placed inside a Forma incubator maintained at 37° C. Modular incubators were purged every 24 hours with an atmosphere of 5% carbon dioxide, 5%, or 1% oxygen, and balance nitrogen. Hypoxic cultures were maintained for 7 days, and RNA, protein, and cell samples were taken after 12 hours and 1, 2, 5, and 7 days of exposure. Half-media changes occurred every 3 to 4 days as needed, as with normoxic controls. For hypoxic exposure period assessment all cultures were performed in 1% oxygen environments. hESCs were cultured in maintenance conditions, and media were changed daily. Apoptosis studies were performed by exposing hEBs to 1% oxygen between days 3 and 10 of differentiation.
Pimonidazole Detection of Hypoxic Exposure, Fluorescent Microscopy.
The hypoxia marker pimonidazole hydrochloride (Hypoxyprobe-1, Chemicon) was used to demonstrate exposure to hypoxic conditions in plate-based cultures. Pimonidazole is a chemically stable, water-soluble, bioreductive probe with an immunoreactive side chain that irreversibly binds to cells exposed to oxygen concentrations at or below 14 μ M (10 mmHg). Thirty mM pimonidazole stock solution was added to hEB culture medium 1 hour prior to harvest at a final concentration of 100 μ M. Upon harvesting hEBs were frozen in Tissue-Tek O.C.T. Compound (Sakura Finetek, Torrance, CA) at − 80° C. Tissues were sectioned and fixed as previously described (39). Sections were incubated overnight at 4° C with mouse anti-pimonidazole MAb (1:50 in PBS, Chemicon, 90204), followed by 4 hours in donkey-derived ALEXA 555 (Eugene, OR). Slides were mounted using Prolong® Gold antifade reagent with DAPI (Molecular Probes).
HIF-1α Protein Detection by Western Blot.
Total cellular protein was extracted from hEBs using RIPA Buffer (Sigma) supplemented with Complete Mini protease inhibitors (Roche Diagnostics, Mannheim, Germany) per the manufacturer’s instructions at a concentration of 107 cells/1 mL buffer. Lysates were frozen at − 80° C for 24 hours to ensure proper disruption of the cellular membrane. Protein lysates were dissolved in Laemmli buffer supplemented with β-mercaptoethanol and boiled for 5 minutes. Twenty-two microliters of sample/lane were subjected to SDS-PAGE in 7.5% gels and in Tris/glycine buffer. Proteins were transferred to nitrocellulose membranes. HIF-1α was detected using mouse anti-HIF-1α MAb (1:500, 610958, BD Transduction Laboratories) and HRP-conjugated goat anti-mouse IgG (1:1000, sc-2005, Santa Cruz Biotechnology). Actin was detected using goat anti-ACTIN polyclonal Ab (1:1000, sc-1616, Santa Cruz) and HRP-conjugated donkey anti-goat IgG (1:5000, sc-2020, Santa Cruz). Protein-antibody complexes were detected using Super-Signal® West Pico or West Dura Chemiluminescent Substrates (34077 or 34075, Pierce) and developed on Kodak X-OMAT LS imaging film for 1–5 minutes.
Reverse Transcription and Quantitative PCR Analysis.
Total RNA was extracted from EBs using an RNeasy mini kit (Qiagen, Valencia, CA), and homogenized with a Qiashredder (Qiagen), according to the manufacturer’s instructions. Total RNA was quantified by UV spectrophotometry, and 0.2 μ g RNA was used for each RT reaction as previously described (39). QPCR samples were prepared in triplicate 25 μ L reactions including 12.5 μ L 2× SYBR® Green Master Mix (Applied Biosystems), 10 ng of reverse transcribed cDNA, and forward and reverse primers (Table 1) at a final concentration of 200 nM. QPCR was preformed on an ABI Prism 7000 SDS machine (Applied Biosystems) with a program of 95° C for 15 min, and 40 cycles of 95° C for 30 s, 55° C for 1 min, and 72° C for 30 s. Fluorescence acquisition occurred at the end of the 72° C extension stage. Gene expression at each time point is reported as a relative fold expression over hESCs (day 0 and in maintenance conditions) and is normalized to actin expression in each sample (43). Error bars demonstrate the standard error of triplicate samples and account for variance in actin and the gene of interest for both the sample and the hESC control (43). Statistical significance was determined by Student’s t test with P value less than 0.05. The specificity of the reaction was checked postamplification by melt curve analysis.
Flow Cytometry.
Single cells resulting from trypsin disassociation of hEBs (above) were processed for flow cytometry as previously described (Cameron, 2006). Apoptotic cells were identified by Annexin V-PE staining using Apoptosis Detectin Kit (559763, BD Pharmingen). Viable cells were identified by 7-amino-actinomycin D exclusion. Jurkat cells treated with camptothecin (5 μ M) were used as a positive control. All analyses were performed with a FACSCalibur (Becton-Dickinson, San Jose, CA) and Cell Quest Pro and FloJo analysis software.
Results
Bioreactor Cultivation of hESC-Derived EBs in Reduced Oxygen Environments.
To monitor and tightly control the oxygen environment in which hEBs were cultured, a closed 1-L glass reactor outfitted with a Clark-type oxygen probe was used to measure the dissolved oxygen concentration within the differentiation media. This stirred culture system provides a more homogeneous culture environment, simplifying process monitoring and control. Culturing hEBs in spun conditions produces hEBs with greater uniformity, improved cell numbers, and with normal hematopoietic differentiation (39). Here, hEBs were cultured for 16 days in a stirred bioreactor with dissolved oxygen concentration of 5% and 25% of saturation. These dissolved oxygen concentrations correspond to approximately 1% and 5% ambient oxygen. Throughout the culture, probe measurements were recorded every 5 minutes. The 5% and 25% of saturation set points were maintained by online manipulation of oxygen and nitrogen flow controllers run with LabVIEW software. An increased oxygen flow rate was necessary to maintain the dissolved oxygen concentration in response the increased oxygen consumption of the expanding hEBs exposed to 25% oxygen saturation (Fig. 1A). A balance equation on oxygen transfer and consumption was used to calculate oxygen uptake rate of live cells (42). The specific oxygen uptake rate of cells was determined to range from 1–3 mmol/1010 cells/hour throughout the culture exposed to 25% of saturation. Due to decreased cell proliferation and low oxygen consumption, the oxygen uptake rate of hEBs cultivated under 5% oxygen saturation could not be accurately determined (data not shown).
hEBs were seeded at an initial cell concentration of 1 × 105 cells/mL, and reached a final concentration of 1.7 × 106 cells/mL when cultivated in 25% oxygen saturation (Fig. 1B). This represents approximately a 17-fold expansion over the 16-day culture and is consistent with growth rates and expansion of hEBs cultivated under spun conditions previously reported (39). The number of viable cells from hEBs cultivated in 5% oxygen saturation decreased by the fourth day of cultivation to 4 × 104 cells/mL, though recovered to reach 1.4 × 105 cells/mL by the end of the 16-day culture period (Fig. 1B).
We hypothesized that exposure of hEBs to reduced oxygen environments would stabilize HIF-1α protein, allowing it to accumulate in the cell cytoplasm, resulting in increased transcription of downstream targets. Surprisingly, immunoblots were unable to detect HIF-1α expression from hEBs at the 16-day endpoint of either 5% or 25% of oxygen saturation culture (Fig. 1C), and only modest increases of HIF-1α targets, VEGF, and GLUT-1 were detected (data not shown).
HIF-1α Accumulates in EBs Cultured in Reduced Oxygen.
To address the decreased cell proliferation and inability to detect HIF-1α expression after 16 days of hEB development, we next focused on reduced durations of hypoxic exposure. As previous studies by our group and others have demonstrated the development of progenitor cells with hematopoietic and endothelial potential arise from hEBs after approximately 7 to 10 days of culture (33, 35, 44), we chose to expose hEBs to hypoxic conditions for a 7-day period, starting at day 11 of differentiation.
hEBs were cultured under normoxic (21% oxygen) or hypoxic conditions with oxygen atmospheres of 1% and 5% for 7 days. Samples were monitored for protein and transcript expression after the first 12, 24, and 48 hours, and again at the fifth and seventh days of culture. Initial studies verified that hEBs were sufficiently exposed to constant reduced oxygen environments by staining for formation of reductively activated pimonidazole hydrochloride (PH) adducts. PH binds irreversibly to cells at dissolved oxygen concentrations at or below 14 μ M (45). hEBs cultured in both 5% and 1% oxygen atmospheres were intensely stained for presence of PH adducts after just 12 hours of hypoxic exposure, and equally so after 7 days of exposure (Fig. 2).
Immunoblots for HIF-1α protein demonstrate accumulation of HIF-1α could be detected as early as 12 hours in both 1% and 5% oxygen cultures. The level of HIF-1α reaches a maximum between days 1 and 2 (Fig. 3A). Interestingly, HIF-1α expression was significantly reduced by the fifth and seventh day of exposure, despite the constant reduced oxygen conditions (Fig. 3A). This is consistent with our findings from the initial 16-day bioreactor experiments (Fig. 1C). As expected, HIF-1α was not detectable above basal levels in hEBs exposed to normoxic conditions at any time point (Fig. 3A, top).
Hypoxia Induces Transcription of HIF-1α Targets in hEBs.
After 12 hours, targets of HIF-1α , VEGF and GLUT-1 were also more highly transcribed in samples cultured in both 1% and 5% oxygen atmospheres, as compared with 21% controls (Fig. 3B and C). Quantitative PCR analysis demonstrates expression of VEGF to be 7.0-, 6.0-, and 2.6-fold more highly expressed in 1%, 5%, and 21% oxygen, respectively, compared with hESCs. Similarly, GLUT-1 expression is 4.9-, 5.5-, and 1.5-fold more highly expressed in 1%, 5%, and 21% oxygen than in hESCs. This increased transcription continued through 5 days of exposure. Parallel to HIF-1α protein expression, these HIF-1α responsive genes also demonstrated reduced expression at latter time points of hypoxic exposure. Notably, increased VEGF and GLUT-1 expression in hEBs represent statistically significant differences between 1% or 5% cultures and 21% controls at each time point (P value < .05). As expected, since HIF-1α stabilization under hypoxic conditions is regulated post-translationally, we show that HIF-1α transcripts were expressed at similar levels in samples cultured in 1% and 5% oxygen conditions and in hEBs cultivated in 21% oxygen as compared with undifferentiated hESCs (Fig. 3D).
Hypoxia Inhibits Growth and Induces Apoptosis.
We monitored hEB growth kinetics throughout the duration of hypoxic exposure under 1% and 5% oxygen conditions. These studies found markedly reduced cell expansion under hypoxic conditions. hEBs were initially cultured at a concentration of approximately 4 × 105 cells/mL (Fig. 4A). Cell numbers from 1% oxygen cultures did not increase during the 7-day period, though no noticeable reduction in viable cells was seen by trypan blue exclusion (data not shown). Total cell numbers from hEBs exposed to 5% oxygen had a modest increase to 6 × 105 cells /mL by the seventh day of hypoxic exposure representing a 1.5-fold increase. In contrast, hEBs cultivated in normoxic conditions reached 11.5 × 105 cells/mL, representing a 2.8-fold increase over the same 7-day window, consistent with our previous findings for hEB expansion (Fig. 3A) (39).
To assess better if the reduced cell proliferation under hypoxia was due to apoptosis in hEBs exposed to low oxygen, we quantified the number of apoptotic cells using Annexin V-PE and 7-amino-actinomycin (7AAD) (Fig. 4B). hEBs were subjected to 1% oxygen exposure for up to 7 days. hEB-derived cells from each culture were analyzed on day 10 of differentiation following the indicated period of hypoxic exposure. Percentages of viable cells (7AAD− Annexin V−) quantified by this method remained at 88% following 12 hours of hypoxic exposure and fell to 67% in cultures that had been exposed to hypoxia for 7 days. Early apoptotic cells (7AAD− Annexin V+) made up 3.1% of total cells after 12 hours of hypoxic exposure yet reached 11% in cultures exposed to hypoxia for 7 days. Cells in late apoptosis or having already succumbed to apoptotic cell death (7AAD+ Annexin V+) comprised 3.4% of the population after 12 hours of exposure and 16% of cells cultured for 7 days in hypoxic conditions. These results suggest that prolonged exposure to hypoxic conditions results in reduced cell proliferation due at least in part by stimulation of apoptotic pathways.
HIF-1α Accumulation Is Transient Independent of Differentiation Stage.
To determine if similar responses to hypoxic exposure, including transient HIF-1α accumulation and stimulation of downstream targets, could be elicited from hEBs at different stages of differentiation, hEBs were exposed to 1% oxygen for 7 days beginning at days 1, 4, 7, and 11 of differentiation. Before and after hypoxic exposure, hEBs were maintained in 21% oxygen conditions, and a control population of hEBs was maintained in 21% oxygen for the duration of differentiation.
As before, HIF-1α was detected after 12 hours of exposure in each hypoxic exposure period and reached maximum expression between 1 and 2 days (Fig. 5). Consistent with exposing hEBs to different reduced ambient oxygen conditions (Fig. 3A), in each hypoxic exposure period examined here, HIF-1α expression was reduced to near undetectable levels by the fifth and seventh day of hypoxic culture (Fig. 5). Interestingly, this same transient accumulation of HIF-1α was detected in undifferentiated hESCs cultured at 1% oxygen and otherwise standard maintenance conditions (Fig. 5, top). As expected, HIF-1α expression was not detectable in cells exposed to normoxic conditions at the beginning or end of any exposure period. Therefore, these studies demonstrate a transient pattern of stabilized HIF-1α expression in hESCs either being maintained in an undifferentiated state or induced to differentiate and as independent of the stage of differentiation.
Hypoxia Stimulates Targets Independent of Differentiation Stage but Does Not Modulate Differentiation.
Exposure of hEBs to hypoxia at different stages of differentiation again revealed VEGF (Fig. 6A) and GLUT-1 (Fig. 6B) to be upregulated after 12 hours of exposure, consistent with HIF-1α accumulation (Fig. 5). This rapid upregulation was detected during all hypoxic exposure periods, independent of the stage of hEB differentiation. Here, hypoxic hEBs demonstrated on average a 3-fold higher VEGF expression compared with hEBs maintained in normoxic conditions in each exposure period examined. Hypoxic hEBs also demonstrated approximately 4-fold higher GLUT-1 expression.
As hEBs were being exposed to hypoxic conditions at different stages of differentiation, we monitored the expression of genes marking both pluripotency and differentiation. As expected upon differentiation, OCT-4, a marker for maintenance of undifferentiated hESCs, was significantly downregulated but was not differentially expressed during any hypoxic exposure period as compared with 21% controls (Fig. 6C). Similarly, OCT-4 was not differentially expressed by exposing hESCs in maintenance conditions to hypoxia (data not shown).
Expression of genes marking differentiation toward each embryonic germ layer: mesoderm (BRACHURY, Fig. 6D), endoderm (ALPHA-FETOPROTEIN, Fig. 6E), and ectoderm (NEUROFILAMENT 200, Fig. 6F) were monitored. These genes were expressed with expected kinetics, though they did not appear specifically modulated toward any one germ layer, as they were not differentially expressed in response to any 7-day hypoxic exposure window as compared with 21% controls. These results suggest hypoxic exposure alone is not sufficient to induce differentiation of hESCs cells toward one specific germ layer. Genes marking later stages of differentiation, including ε -GLOBIN, C-MET, and GATA-2, were also evaluated but found not to be significantly induced in these heterogeneous populations of hEB cells at these early stages of differentiation (Fig. 6G–I).
Hypoxia Transiently Inhibits Growth Only During Hypoxic Exposure.
Evaluation of cell expansion rates of hESC-derived cells when cultured under hypoxic conditions during different stages of hEB differentiation again demonstrated reduced cell proliferation. However, this reduction of cell expansion occurred only during the periods of time in which the hEBs were exposed to reduced oxygen. Importantly, when the cultures were then returned to normoxic conditions after 7 days of hypoxia, in all cases cells returned to near normal growth rates (Fig. 6J). hEBs were initially plated at a concentration of approximately 1 × 105 cells/mL. Although there was modest reduction in total cell growth and proliferation in cells exposed to hypoxic conditions at earlier stages of differentiation (exposure starting at day 1 and 4 of differentiation), overall the hEB cultures were all able to attain near normal cell numbers by the end of the 17-day differentiation period, reaching between 0.9 and 1.2 × 106 cells/mL (Fig. 6J).
Hypoxic Exposure and Hemato-Endothelial Development.
Finally, we evaluated the hematopoietic and endothelial differentiation potential of hEBs exposed to both early and late hypoxic exposure periods, namely between days 4 and 11 and between days 10 and 17 of differentiation. The hematopoietic and endothelial potential of hEBs cultured under hypoxic conditions was evaluated by flow cytometric analysis for CD34, CD31, and CD45. hEBs cultured in reduced oxygen between days 4 and 11 (Fig. 7A–C) produced similar percentages of CD34+cells as compared with the 21% control (Fig. 7A). Here, peak development of CD34+ cells occurred earlier in hEBs exposed to 1% oxygen (day 7), than in those exposed to 5% (day 11). In contrast, peak development of CD34+CD45+ cells (more specific population of hematopoietic progenitor cells) was seen at day 17 of culture, and was not found to be markedly different between the hypoxic and normoxic conditions (Fig. 7C).
hEBs cultured in reduced oxygen between days 10 and 17 of differentiation (Fig. 7D–F) demonstrated an early increase in CD34+ cells in hEBs exposed to 1% oxygen as compared with cells cultured in 5% and 21% oxygen during this hypoxic exposure period (Fig. 7D). However, this increase in proportion of cells expressing CD34 was not maintained after returning cells to normoxic conditions. Additionally, the development of CD34+ cells declined at later times through this period of differentiation, unlike other normoxic controls (Fig. 7A). Development of CD34+CD45+ cells were slightly increased at day 14 of culture, though this population diminished at later time points compared with the normoxic (21%) cultured cells (Fig. 7F).
Discussion
In these studies, we investigate the cultivation of hESC-derived EBs in environments of reduced oxygen concentrations in an effort to define more optimal conditions to promote the differentiation of hESCs. Furthermore, these studies provide insight into the molecular responses of hEBs under short- and long-term exposure to hypoxic conditions.
Multiple studies using isolated hematopoietic cells have demonstrated the role of oxygen to be important for hematopoietic stem cell development. Mature blood cells and rapidly proliferating progenitors are thought to reside in areas of the bone marrow with higher oxygen tensions, whereas immature progenitors reside in regions of relatively low oxygen, between 2–7% (25, 46). In vitro expansion of hematopoietic progenitor cells from mouse bone marrow (47–49), human bone marrow (25, 27), or human cord blood (25, 26, 50) have demonstrated increased cell expansion, cell survival, production of progenitor cells, or marrow repopulating ability in ambient oxygen concentrations reduced to between 0.9–5%. These findings could be partially attributed to reduction in oxidative stresses (51, 52). However, work in recent years with transgenic mice and mouse ESCs has focused on the role of hypoxic inducible proteins to regulate responses to oxygen deprivation as a means to potentially stimulate the development of hematopoietic and endothelial progenitor cells (8, 24).
Many hypoxic-based studies are performed by manipulation of atmospheric oxygen conditions without monitoring the dissolved oxygen environment to which cells are exposed. Dissolved oxygen concentrations are heavily dependant upon cell densities, cellular metabolism, and oxygen diffusion through the culture medium. Here, we utilized a stirred culture system in which the dissolved oxygen concentration could be monitored and tightly controlled. Additionally, stirred culture systems have been well characterized and provide a homogeneous culture environment amenable to process control. Previous work by our group demonstrated increased cell numbers, efficient hematopoietic differentiation, and diminished aggregation of hEBs cultivated in stirred vessels (39).
With this reactor system, we were able to measure and control the dissolved oxygen concentration at 25% and 5% saturation during repeated 16-day cultures of hEBs. Cultivation in 25% of saturation allowed for measuring the specific oxygen uptake rate of hEB cells and was found to be 1–3 mmol/1010 cells/hour. However, markedly attenuated cell expansion under the lowest oxygen concentration was found, and subsequent studies suggest that this is likely due to increased apoptosis in hEBs cultured under hypoxic conditions (Fig. 4). Additionally, HIF-1α stabilization was not detected after 16 days of culture. These results subsequently lead us to utilize a screening method in which multiple oxygen concentrations could be tested in parallel. Although the dissolved oxygen concentration in these cultures was not directly measured, the hEBs were maintained below a cell concentration where the oxygen uptake would be greater than the rate of oxygen diffusion into the media. Similarly, under no circumstances would the dissolved oxygen concentration exceed that which is saturated by the 1%, 5%, or 21% ambient oxygen. Additionally, the duration of hypoxic exposure was decreased to 7 days, and the effect of imposing these 7 day hypoxic exposures was explored during different stages of hEB differentiation.
Under these conditions, we demonstrate exposure of hEBs to 5% ambient oxygen was sufficiently low to detect accumulation of HIF-1α as early as 12 hours after exposure (Fig. 3). More interestingly, HIF-1α protein expression was transient under constant hypoxic conditions, and was independent of the stage of differentiation at which hypoxic exposure was begun (Fig. 4). This likely explains why HIF-1α was not detected at the end of the initial 16-day reactor cultures (Fig. 1). Moreover, this transient expression was not restricted to differentiating cells, as undifferentiated hESCs cultured in maintenance conditions and in low oxygen demonstrated transient HIF-1α accumulation (Fig. 5). Staining for protein adducts of reduced pimonidazole verified that hEBs experienced constant hypoxic conditions for the duration of the 7-day hypoxic exposure in both 1% and 5% oxygen atmospheres (Fig. 2). To our knowledge, this transient HIF-1α stabilization has not been previously reported.
Furthermore, hEBs maintained in 21% oxygen and proportionally sized to those in hypoxic conditions show no signs of oxygen deprivation, even at their core (Fig. 2), demonstrating that oxygen diffusion is not a limiting factor for these large cell aggregates (approximately 500 μ m in diameter) when cultured in normoxic conditions. A study measuring pericellular oxygen pressure within mouse EBs with diameters ranging from 510–890 μ m, found only moderate reductions in oxygen tension, which fell to 100 mmHg (13.2%) at the EBs core when cultured in 21% oxygen (53), suggesting that, like hEBs, their density does not prevent oxygen diffusion to the core of the aggregate.
As HIF-1α expression is regulated exclusively at the protein level, transcripts of HIF-1α were not more highly expressed in samples cultured in 1% or 5% oxygen atmospheres at any time point examined, nor were they more highly expressed in hEBs cultivated in 21% oxygen. However, increased expression of downstream HIF-1α targets, VEGF and GLUT-1, was demonstrated. Although stimulation of vegf and other hypoxic responsive genes alone was not sufficient to modulate cell differentiation toward hematopoietic fates in these studies, its stimulation via hypoxic cultivation may alleviate the need for its exogenous addition in combination with other cytokines to other culture systems (33, 35, 37, 54).
Finally, these studies highlight what may be another key difference between mouse and human ESCs, and ESC-derived EBs. Several differences, including LIF requirements (55), surface antigen expression patterns (56, 57), and growth kinetics (58, 59) have been well documented. Additionally, several studies utilizing mouse embryonic stem cells (mESCs) cultivated under hypoxic conditions have been shown to promote the commitment to mesoderm and generation of hemato-endothelial progenitor cells (8, 23, 28, 29). However, the culture methods of mouse and human ES cells are quite different and may explain the differences observed in their expansion and differentiation under hypoxic conditions. For example, the formation of mEBs is achieved from a single cell suspension of rapidly expanding mESCs, whereas hEBs are formed from intact colonies of hESCs by collagenase or dispase digestion to form small cell aggregates that expand more slowly (60–62). This may allow the mEBs to proliferate and differentiate under hypoxic conditions more readily, while hEBs may not be able to overcome the impact of apoptotic induction imposed by hypoxic culture.
Prolonged hypoxic exposure has been shown to induce apoptosis in mEBs and HUVEC cells exposed to > 5 days of hypoxia (63, 64). Importantly, we found returning hESC-derived EBs to normoxic conditions following 7 days of hypoxic exposure restored them to near normal growth rates. However, a further reduction of hypoxic exposure time, to less than the 7 days investigated here, may be essential for providing an environment in which differentiating hematopoietic and endothelial cells can flourish. This exposure period must also coincide with a stage of hEB differentiation where induction of hypoxic responsive genes will impact differentiation of progenitor cells present at that particular time.
In conclusion, we describe a transient accumulation of HIF-1α protein and downstream targets in both undifferentiated hESCs and hEBs. These results are an important step in developing a better understanding of the role of oxygen in human hematopoietic and endothelial cell development and hESC-derived cell differentiation, and are an important step toward defining conditions for novel human stem cell-based technologies.
qPCR Primer Pairs

Bioreactor cultivation of hESC-derived EBs.

Immunohistochemical staining of hEBs under hypoxic or normoxic conditions. Fluorescent microscopy revels positive immunoreactivity (red) with pimonidazole hydrochloride adducts formed within hEBs exposed to both 5% and 1% oxygen atmospheres. Staining demonstrates constant exposure to reduced oxygen for 7 days. Cell nuclei stained with DAPI (blue). Scale bar is 100 μ m.

Transient accumulation of HIF-1α and transcription of hypoxic responsive genes in hEBs. (A) Western blot analysis of HIF-1α protein following 0.5, 1, 2, 5, and 7 day(s) of exposure to 21%, 5%, and 1% oxygen atmospheres. HIF-1α from HeLa cells exposed to anoxia for 12 hours is shown as a control (lane C). ACTIN expression is shown as loading control. HIF-1α accumulation in hEBs exposed to both 5% and 1% oxygen begins as early as 12 hours (0.5 days), and peaks between 1 and 2 days of exposure. HIF-1α protein levels return to basal levels by the fifth and seventh days of exposure. Hypoxic responsive genes (B) VEGF and (C) GLUT-1 were upregulated in 5% and 1% environments. VEGF and GLUT-1 expression levels represent statistically significant differences in 1% or %5 oxygen cultures as compared with 21% controls at each time point (P value < .05). (D) HIF-1α mRNA levels did not change in response to hypoxic exposure. Error bars represent standard error of triplicate qPCR reactions.
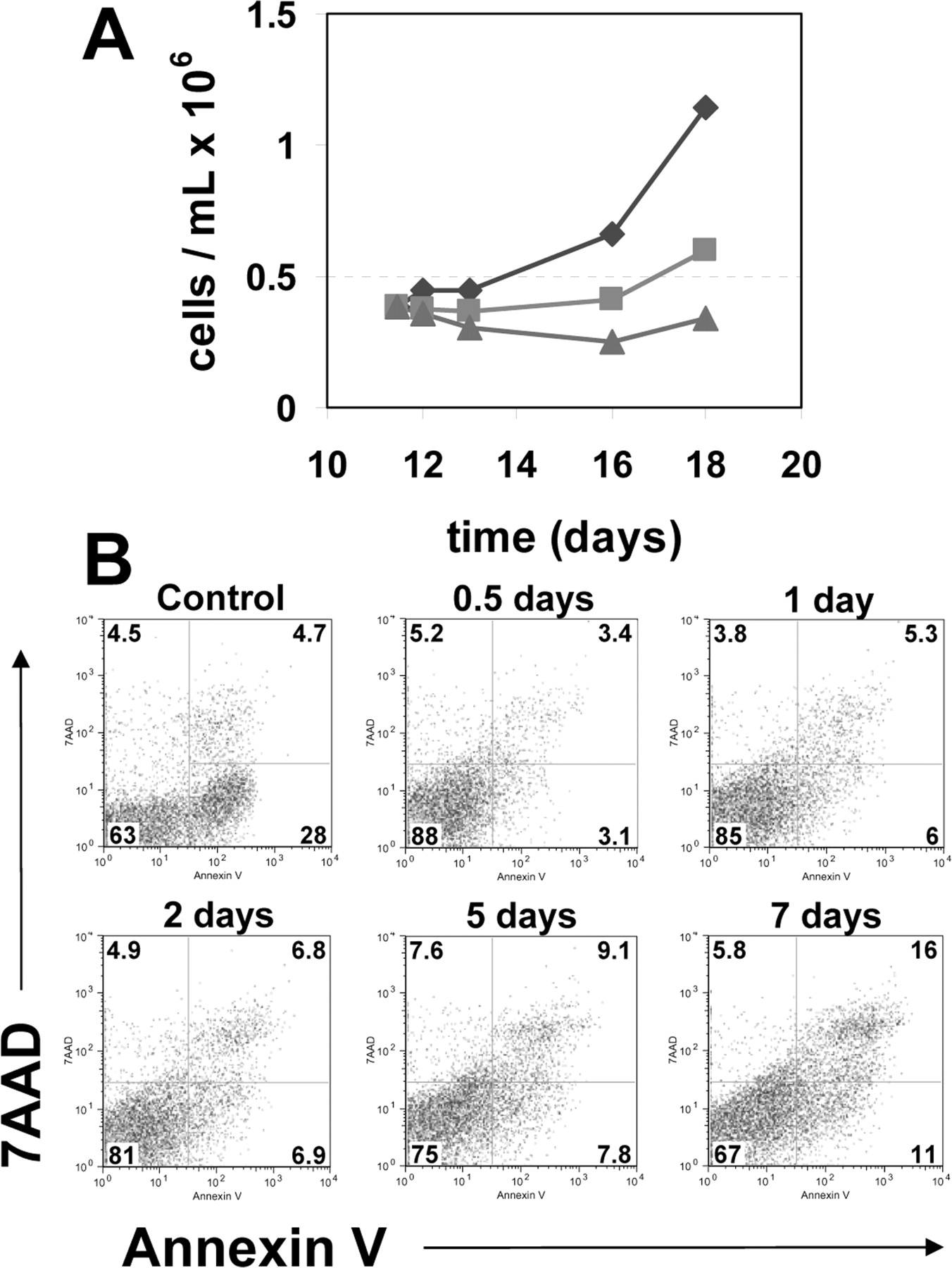
Growth kinetics and apoptosis in hEBs cultivated under hypoxic conditions. (A) hEBs placed in 5% (▪) and 1% (▴) oxygen atmospheres at a cell concentration of approximately 4 × 105 cells/mL on day 11 of differentiation demonstrate attenuated growth kinetics in an oxygen dependant manner. Twenty-one percent (♦) shown as a control. (B) Time course of apoptosis in response to hypoxic exposure. Flow cytometric analysis of cell viability and apoptosis induction in hEBs on day 10 of differentiation following 0.5, 1, 2, 5, or 7 days of 1% oxygen exposure. Viable cells exclude 7-amino-actinomycin uptake (7AAD) (y-axis) and apoptotic cells that bind Annexin V-PE (x-axis). Jurkat cells induced to undergo apoptosis following exposure to camptothecin are shown as a positive control.
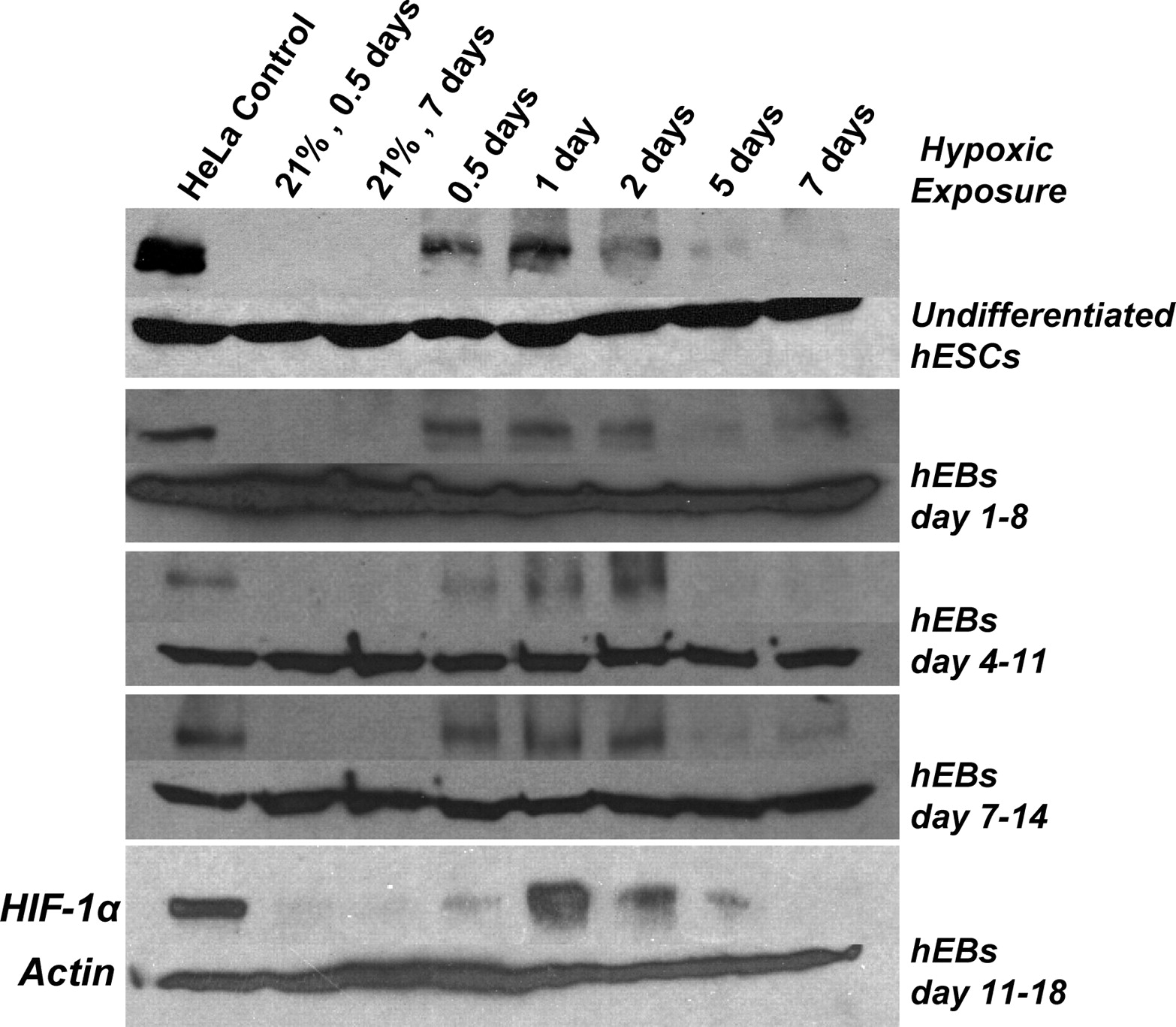
Transient accumulation of HIF-1α is independent of hypoxic exposure period. Western blot analysis of HIF-1α protein following 0.5, 1, 2, 5, and 7 days of exposure to 1% oxygen is shown. hESCs in maintenance conditions were subjected to 7 days of hypoxic exposure (top row) and hEBs were placed in hypoxia for 7 days starting on days 1, 4, 7, and 11 of differentiation. Control hEBs maintained in 21% oxygen throughout differentiation did not demonstrate HIF-1α accumulation at 12 hours (0.5 day) nor at 7 days of the respective hypoxic exposure period. HIF-1α from HeLa cells exposed to anoxia for 12 hours is shown as a positive control.
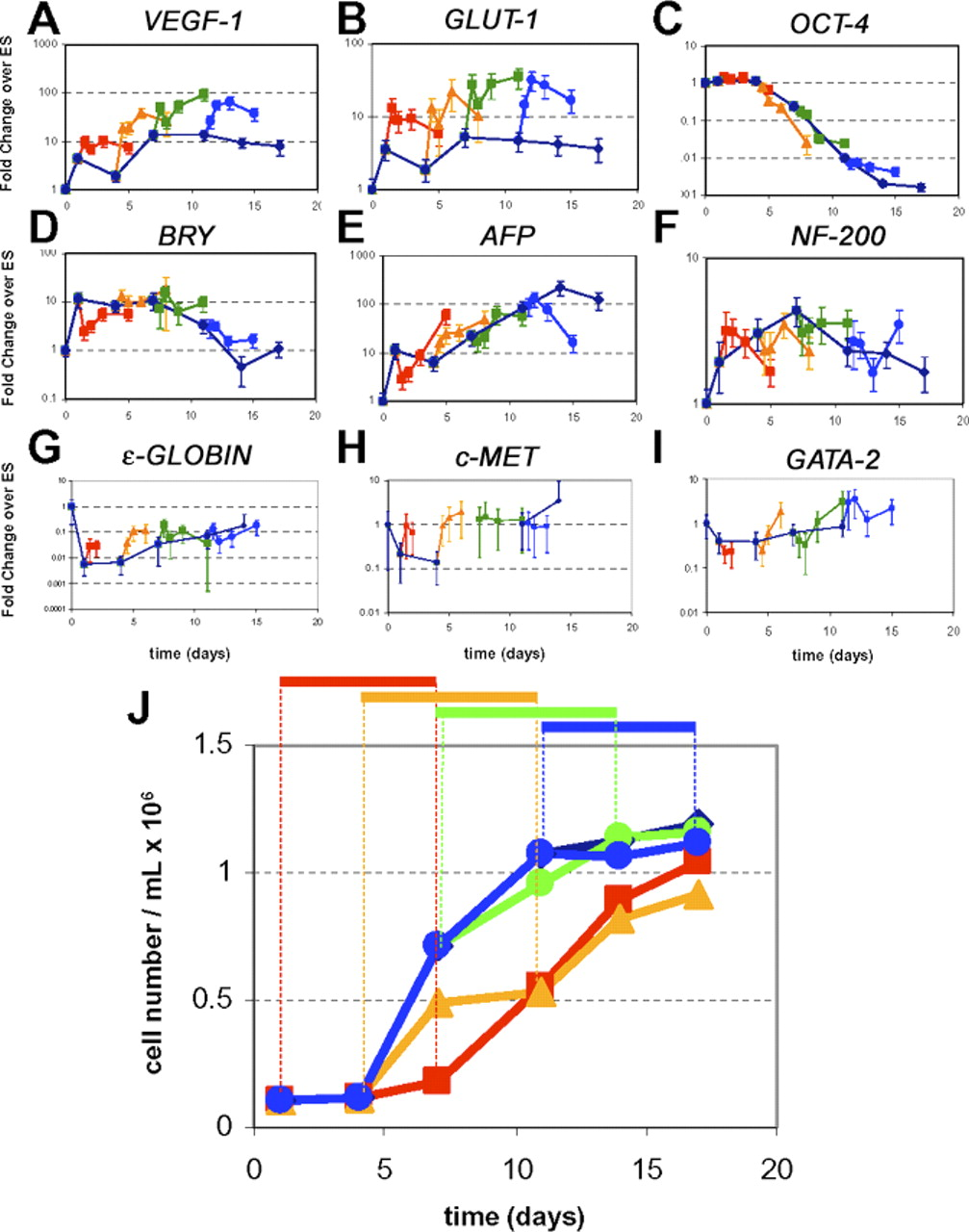
Hypoxic responsive gene expression in hEBs during different hypoxic exposure periods. Gene expression from hEBs subjected to 7 days of hypoxic exposure (1% oxygen) starting on days 1 (red square), 4 (yellow triangle), 7 (green square), and 11 (blue circle) of differentiation is shown. Expression from hEBs maintained in 21% oxygen (blue diamond) throughout differentiation is shown as a control. Hypoxic responsive genes (A) VEGF and (B) GLUT-1 were upregulated in response to 1% oxygen environments as early as 12 hours, and throughout the duration of hypoxic exposure in each exposure window. (C) Pluripotent cell marker OCT-4 did not have a significant change in expression during any 7-day hypoxic exposure period, as compared with 21% controls. Genes typical of differentiation to three embryonic germ layers (D) BRACHURY (BRY, mesoderm) (E) ALPHA-FETOPROTEIN (AFP, endoderm) (F) NEUROFILAMENT 200 (NF200, ectoderm) were not significantly upregulated in response to any 7-day hypoxic exposure period as compared with 21% controls, nor were genes typical of more mature differentiation, (G) ε -GLOBIN (mesoderm), (H) HEPATOCYTE GROWTH FACTOR RECEPTOR (C-MET, endoderm), or (I) GATA-2 (mesoderm). Error bars represent standard error of triplicate qPCR reactions. (J) hEBs growth kinetics in response to different hypoxic exposure periods. On day 1, hEBs were seeded at cell concentration of approximately 1 × 105 cells/mL. hEBs demonstrate attenuated growth kinetics in 1% oxygen, but hEB growth returns to near normal rates following removal from hypoxic conditions. A color version of this figure is available in the online journal.
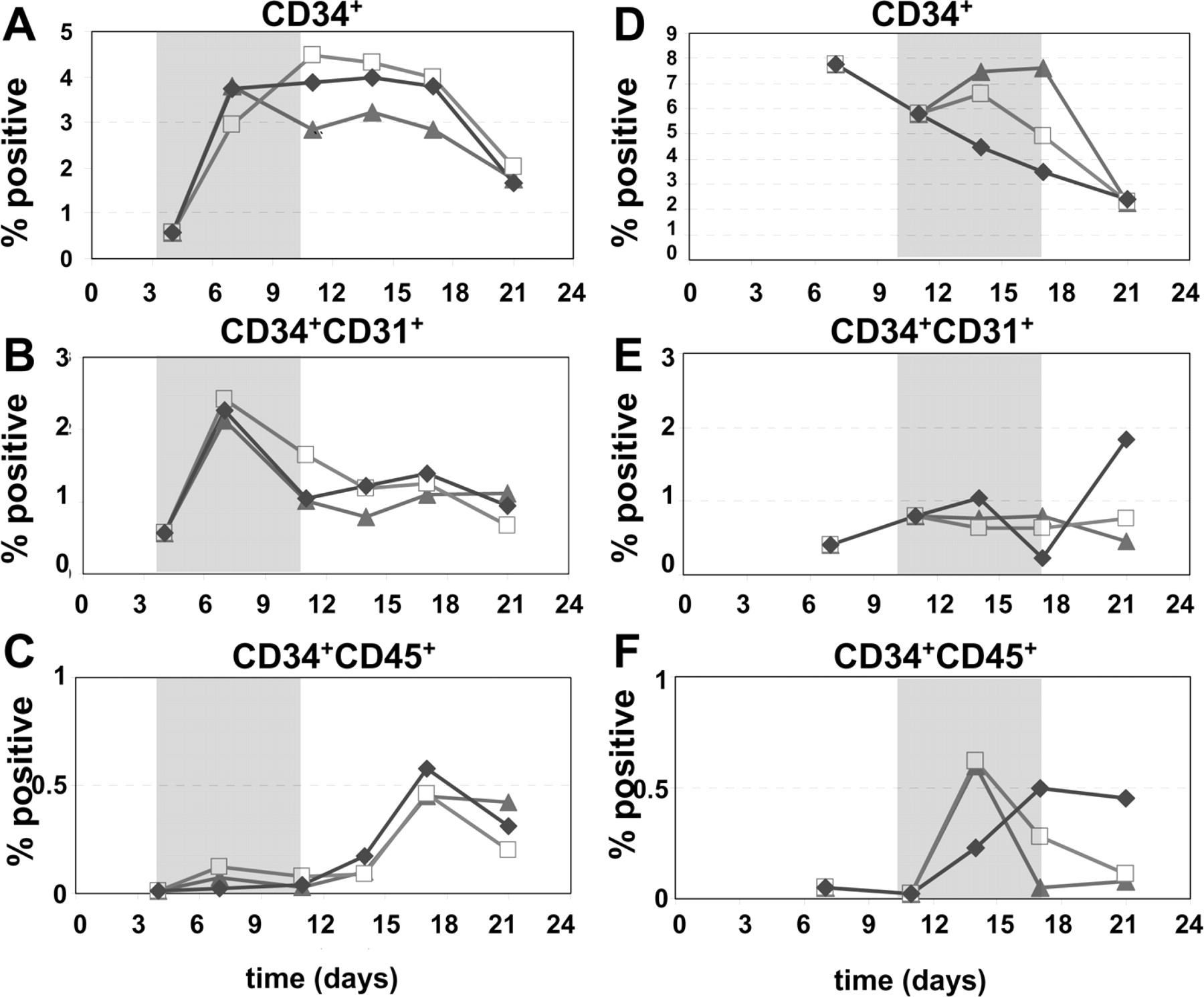
Kinetics of hematopoietic and endothelial differentiation. Differentiation potential was quantified by flow cytometry. Percentage of total cells being (A) CD34+, (B) CD34+CD31+, and (C) CD34+CD45+ in cultures exposed to hypoxia from days 4–11 of differentiation. Percentage of total cells being (D) CD34+, (E) CD34+CD31+, and (F) CD34+CD45+in cultures exposed to hypoxia from days 10–17 of differentiation in 1% (▴), 5% (□), and 21% (⋄) ambient oxygen cultures is shown. Shaded box designates period of hypoxic exposure.
Footnotes
This work was funded in part by NIH R21 HL72000 and R01 HL77923 (DSK). Funding was also provided by a grant from Chiron Corp. (now Novartis). CMC was supported by an NIH Biotechnology Training Grant.
Acknowledgements
Special thanks to Dr. Patrick Hossler for help with setting up, running, and analyzing data from bioreactor experiments. We thank Julie K. Morris for her help with flow cytometry and PCR, Drs. Lucas Chase and Terry Burns for assistance with selection of qPCR primers, and Dr. Nobuaki Kikyo and his lab members for guidance on Western blotting, and Tracy Bergemann (University of Minnesota Cancer Center) for consulting on biostatistics.