
Abstract
The developmentally important hedgehog (Hh) pathway is activated in ischemic tissue, and exogenously administered Sonic hedgehog (Shh) supports tissue repair after cardiac ischemia. Hence, it is currently assumed that the endogenous increase in Shh during ischemia serves a beneficial role in limiting cardiac tissue damage. To prove or refute this hypothesis, we treated mice with the smoothened (Smo) inhibitor cyclopamine to block the Hh pathway during myocardial ischemia and reperfusion. The experimental induction of myocardial ischemia resulted in activation of the Hh pathway and hallmark features of myocardial damage, such as left ventricular dilatation and reduced cardiac output. Unexpectedly, cyclopamine treatment ameliorated left ventricular dilatation and cardiac output. As the beneficial effect of exogenous Shh was suggested to depend on reduced apoptosis, increased vascularization, and reduced fibrosis, we subsequently assessed the effect of cyclopamine on these processes. Vascularization was similar in cyclopamine-treated and control-treated animals, but increased apoptosis and reduced fibrosis were observed in the cyclopamine-treated animals. Thus, Hh seems to exert a dualistic action in cardiac ischemia in which high exogenous levels are able to foster tissue repair and endogenous Hh seems to be deleterious.
Introduction
The developmental hedgehog (Hh) protein family (1) is pivotal in many embryonic patterning events, and the number of processes in which Hh plays an essential role is expanding persistently. For instance, Indian hedgehog and the more thoroughly characterized homolog Sonic hedgehog (Shh) are critical to the development of the vasculature and of early erythrocytes (reviewed in Refs. 2 and 3). By virtue of its role in establishing left-right asymmetry, Shh is furthermore crucial to the looping of the cardiac tube, which later becomes the heart.
Recently, it has become clear that the Hh pathway is not only active in the developing embryo but also in the adult organism (4). Among others, Hh has been suggested to salvage ischemia-induced tissue damage. Two recent papers of Pola et al. established a strong role for Hh signaling in adult cardiovascular pathophysiology. Shh mediates a profound upregulation of Hh target genes such as vascular endothelial growth factor (VEGF) and the angiopoietins Ang-1 and Ang-2, which were found to induce neovascularization (5). The authors extended their findings and managed to salvage ischemic hind limbs in mice by injecting Shh into the afflicted muscle (6). In another experimental setup, Kusano and colleagues demonstrated that intramyocardial gene transfer of Shh promotes recovery and preservation of left ventricular function in both acute and chronic myocardial ischemia models (7). Hh exerts its beneficial effect by enhancing neovascularization, recruiting bone marrow-derived progenitor cells, and reducing fibrosis and cardiac apoptosis.
Interestingly, characteristic features of both the hind limb and the myocardial ischemia model mentioned above are the endogenous production of Shh and the activation of the associated signaling system. The physiologic importance of this endogenously expressed Shh is unexplored in the myocardial model, but it is thought that the endogenous increase of Shh during myocardial ischemia serves a beneficial role in limiting tissue damage as it does in hind limb ischemia models. To prove or refute this hypothesis, we set up experiments in which mice were treated with cyclopamine to block the Hh pathway before subjecting them to a model of myocardial ischemia and reperfusion (8). Cyclopamine is an alkaloid that specifically inhibits the activating receptor of the Hh pathway, smoothened (Smo) (9–11). In the model, the left anterior descending (LAD) coronary artery was ligated for 30 mins and then reperfused for 1 week. End points of this model were dimensional and functional markers of the integrity of the ventricular structure and function. In addition to these physiologic parameters, immunohistochemical analysis was performed to determine the effect of cyclopamine on apoptosis, vascularization, and fibrosis in the heart.
Our results show that in contrast to the prevailing notion that endogenous Shh is involved in a beneficial response after cardiac ischemia, endogenous Shh contributes to ischemia-reperfusion–induced injury. Hence, a dualistic role of Shh signaling in coronary disease is revealed, in which high levels of exogenous Shh as well as inhibition of endogenous production are associated with improved outcome of disease.
Materials and Methods
Animals.
Ten-week-old male C57Bl/6 mice (Harlan Sprague-Dawley Inc., Horst, the Netherlands), weighing approximately 25 g at the time of surgery, were provided free access to food and water and were maintained at the animal care facility of the University of Maastricht according to institutional guideline. Animal procedures were carried out in compliance with institutional standards for humane care and use of laboratory animals. The animal care and use committee of Maastricht University approved all experiments.
Experimental Setup.
Mice were anesthetized with ketamine (100 mg/kg given intramuscularly) and xylazine (5 mg/kg given subcutaneously). Body temperature, monitored with a rectal probe, was maintained at 37° C by using a warming pad and heating lamp. The trachea of each mouse was intubated per os with a stainless steel tube connected to a respirator (Hugo Sachs Electronic rodent ventilator type 845, March-Hugstetten, Germany) set at a stroke volume of 250 μ l and a rate of 210 strokes per minute. After left thoracotomy and exposure of the heart, the LAD coronary artery was ligated with a 6–0 (metric 0) polypropylene suture (Surgipro, Chicago, IL), just proximal to its main branching point. The suture was tied around a 3-mm-long polyethylene tube (PE-10) to induce ischemia. Under microscopic viewing, blood flow was reestablished by removal of the tube after 30 mins of ischemia.
Animals were given intraperitoneal injections of 2 mg/kg cyclopamine (a dose previously shown to completely block the Hh pathway [Ref. 12]; Biomol, Plymouth Meeting, PA) or solvent control (2-hydroxypropyl-β-cyclodextrin) 1 day before ligation of the LAD coronary artery. Immediately before the surgical procedure, echocardiography was performed to determine left ventricular dimension and function (before myocardial ischemia-reperfusion [MIR]). After the operation, animals were treated daily for 7 days with cyclopamine (or solvent) after which a second echocardiography was performed before the animals were sacrificed by cervical dislocation (after MIR). Upon sacrifice, hearts were removed for immunohistochemical analysis, whereas kidneys were collected for assessment of Hh pathway activity to determine the efficacy of cyclopamine treatment. The technicians performing the (randomized) treatment, operation, and analysis were blinded to the identity of the injected solutions.
Immunohistochemistry.
Hearts were fixed in 10% buffered formalin and embedded in paraffin. Slides (4-μ m-thick sections) were stained with hematoxylin and eosin according to routine procedures or incubated in 1% Sirius Red F3B in picric acid for 1 hr to assess fibrosis. For immunohistochemical analysis, paraffin was removed from the slides, and the slides were rehydrated. Endogenous peroxidase activity was quenched with 0.03% H2O2 in methanol, and antigen retrieval was performed by boiling for 5 mins in 10 mM citrate buffer (pH 6.0). Slides were blocked with TENG-T. Patched1 (Ptch1) staining was performed with a 1:250 dilution of anti-Ptch1 G-19 (Santa Cruz Biotechnology, Santa Cruz, CA). Specific active caspase-3 staining was performed with a 1:200 dilution of anti–ACTIVE Caspase-3 antibody (Promega, Madison, WI). TUNEL staining was performed according to the manufacturer’s protocol (DeadEnd Colorimetric TUNEL System, Promega). Smooth muscle actin was visualized by using anti–α-SMA-1 clone 1A4 (1:200 dilution, Sigma-Aldrich, St. Louis, MO). Slides were incubated with the appropriate horseradish pexoxidase-conjugated secondary antibodies, and 3,3′ diaminobenzidine (DAB) staining was used to visualize peroxidase activity. Slides were photographed on a Zeiss Axioskop (Zeiss, Jena, Germany) with a Sony DXC950P CCD camera (Sony, Tokyo, Japan). Caspase staining was quantified by measuring staining intensity; image colors were inverted in Photoshop 7.0 (Adobe Systems, San Jose, CA), and luminosity was determined in three separate areas in the left ventricle. For TUNEL staining, positive nuclei were counted per optical field at random locations in the left ventricle wall. For quantitation of vascularization, heart vessels stained with anti–α-SMA-1 were counted. For assessment of the amount of fibrosis in the ventricle wall, the fraction of Sirius Red–stained ventricle wall thickness was measured at three sites in the ventricle wall.
Western Blot Analysis.
Kidneys were collected and homogenized in Greenberger lysis buffer. Protein content was determined by using a BCA protein assay (Pierce Biotechnology, Rockford, IL) and was equalized, and samples were diluted with 2 × Laemmli buffer. After heating, samples were run on 10% sodium dodecyl sulfate–polyacrylamide gels, and subsequently transferred to blotting membranes. After blocking with 3% Protifar (Nutricia, Zoetermeer, the Netherlands) in TBS-T, membranes were incubated overnight in a 1:2000 dilution of anti–β-actin (Santa Cruz Biotechnology) or a 1:500 dilution of anti-Ptch1 (G-19, Santa Cruz Biotechnology). Membranes were incubated in anti-rabbit secondary antibody (1:2000 dilution, Dako Cytomation, Glostrup, Denmark) and imaged on a LAS3000 dark box (Fujifilm, Tokyo, Japan). Densitometry was performed in Photoshop by using the histogram function in a selected area of constant size for each band. Background values were subtracted, and values for Ptch1 were corrected for those of β-actin.
Echocardiography.
Animals were under light isoflurane anesthesia during transthoracic echocardiography. Transthoracic echocardiography of the left ventricle was performed by using a Hewlett-Packard 15-MHz linear array transducer (15–6 l) interfaced with a Sonos 5500 echocardiography system (Philips, Eindhoven, the Netherlands). Two-dimensional B-mode echocardiograms were captured at a rate of 90 to 120 Hz from parasternal long-axis views as well as from midpapillary short-axis views of the left ventricle. Data were obtained from at least three different images taken in end diastole and peak systole by using EnConcert software (Agilent Technologies, Andover, MA).
Statistical Analysis.
Depicted are the means ± SEM. After exclusion of nonresponders (see the Results section for details), seven animals remained in the control groups, and eight animals remained in the cyclopamine group. Comparison between two groups was done by using unpaired Student’s t test.
Results
The Hh Pathway Is Activated in Ischemic Myocardium.
Previous data suggest that the Hh pathway is activated in the heart of animals exposed to myocardial ischemia (7). To confirm these observations, we analyzed Ptch1 expression in the heart of mice subjected to myocardial ischemia and reperfusion. Ptch1, the inhibitory regulator of the Hh pathway, is a direct target gene of the Hh pathway, and Ptch1 levels are thus indicative of Hh pathway activity (13). As shown in Figure 1A, some baseline expression of Ptch1 was observed in the left ventricle of control animals. Ischemia and reperfusion, however, strongly induced the expression of Ptch1; this result confirmed that indeed the Hh pathway is activated.
Exclusion of Animals Unresponsive to Cyclopamine.
Cyclopamine inhibition of Hh signaling is notoriously variable in its efficacy, mainly because of the drug’s hydrophobicity (14). To eliminate superfluous variation in the experimental setup caused by inefficient cyclopamine bioavailability, we analyzed Hh pathway activity in kidney homogenates of cyclopamine-treated and solvent control–treated animals. Animals in which cyclopamine treatment was unsuccessful will not show decreased Hh pathway activity in organs with an active Hh pathway. As shown in Figure 1B, solvent-treated animals showed robust expression of Ptch1 (indicative of Hh pathway activity). Efficient cyclopamine treatment dramatically reduced Ptch1 expression levels, whereas inefficient cyclopamine treatment did not reduce Ptch1 levels (Fig. 1B). Cyclopamine-treated animals with levels of Ptch1 to β-actin (as determined by densitometry) at or above the average level of Ptch1 to β-actin in all animals were considered nonresponders and were excluded from the study (6 of 16 cyclopamine-treated animals).
Echocardiography.
To assess the effect of cyclopamine treatment on the function of the afflicted tissue, left ventricular dimensions and resultant functional parameters were obtained by echocardiography. The raw preoperative data are shown in Figure 2A, and the raw postoperative data are shown in Figure 2B. As evident from Figure 2A, left ventricular integrity and consequent function were similar between cyclopamine-treated and solvent control–treated animals before the induction of ischemia and reperfusion. These parameters are similar to those observed after 2 weeks in sham-operated animals (8). The induction of ischemia and reperfusion, however, severely compromised left ventricular integrity as evident from both a systolic and diastolic decrease in left ventricle wall thickness and an increase in the internal left ventricular diameter (Fig. 2B). In addition, ischemia and reperfusion severely reduced fractional shortening, the stroke volume, and the ejection fraction, a result indicating diminished ventricle contractility. The strong deterioration of these parameters highlights the severe effect of the 30-min ischemia-reperfusion procedure on cardiac function.
Figure 2C shows the relative values for the above-mentioned parameters as calculated by dividing postoperative values by preoperative values. Cyclopamine treatment reduced left ventricular dilatation as evident from increased systolic and diastolic left ventricle wall thickness (no significant difference) and from a reduced internal diameter. Furthermore, cyclopamine treatment mitigated the reduction of fractional shortening, stroke volume, and ejection fraction induced by ischemia and reperfusion, resulting in postoperative values more similar to preoperative values and thus sham-operated values. Taken together, the echocardiography data show that cyclopamine treatment significantly restored ventricular function inflicted by ischemia and reperfusion.
To assess the consequence of myocardial ischemia and reperfusion on the histologic level, slides were stained with hematoxylin and eosin. As shown in Figure 2D, the left ventricle of mice subjected to ischemia and reperfusion was rather severely dilated. In cyclopamine-treated animals, however, the left ventricle seemed to be almost unaffected, closely resembling the ventricle of controls that were not subjected to ischemia.
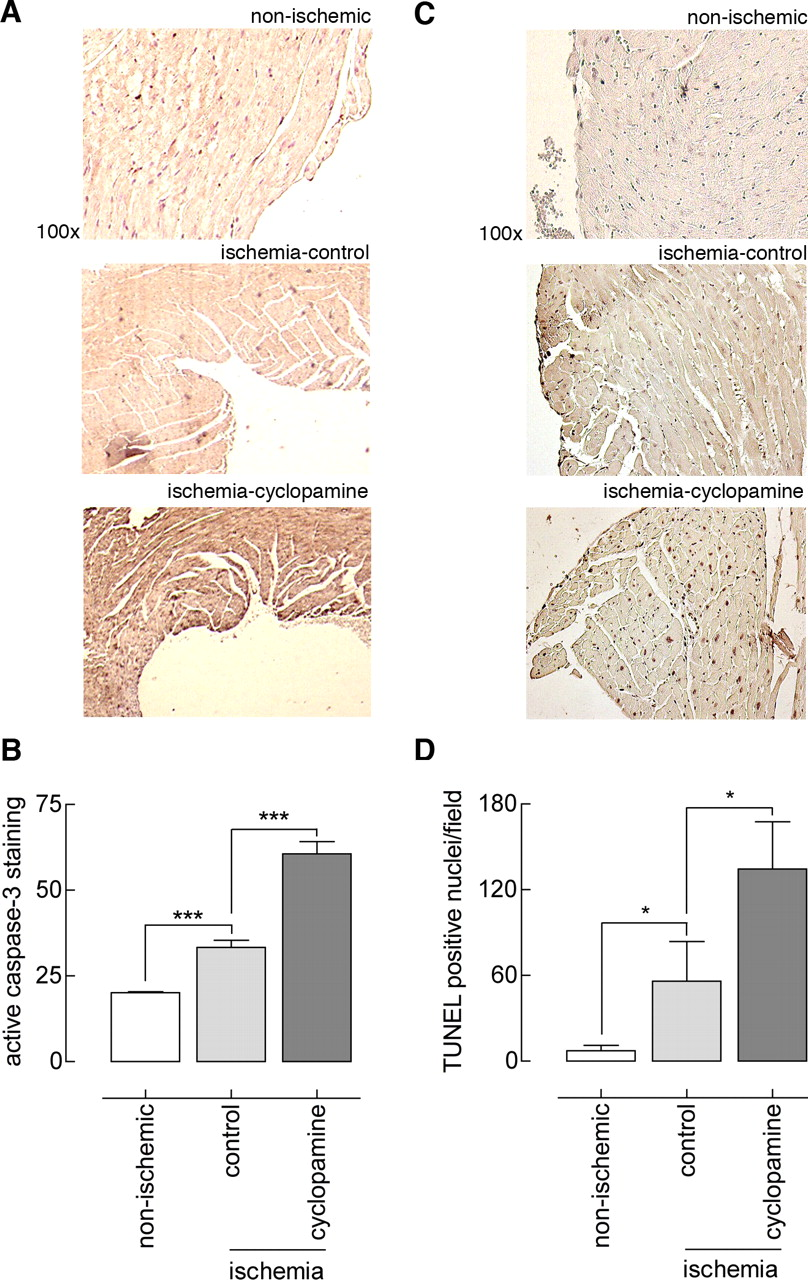
Apoptosis.
The previously described beneficial effect of exogenous Shh was partly attributed to reduced apoptosis in the area of infarction (7). To determine the effect of endogenous Hh on apoptosis, caspase-3 activity in the left ventricle wall was assessed by immunohistochemistry. As shown in Figure 3, some caspase-3 staining was observed throughout the left ventricle of control animals that did not undergo ischemia, whereas ischemia and reperfusion slightly but significantly increased the amount of caspase-3 staining. Caspase-3 staining and thus apoptosis was more strongly augmented in the cyclopamine-treated animals than in the nonischemic controls and solvent-treated animals (Fig. 3A and B). Thus, similar to exogenous Shh, endogenous Hh limited ischemia-reperfusion–induced apoptosis.
To confirm and consolidate the data obtained from the caspase-3 staining, we also employed TUNEL analysis (Fig. 3C and D). Hardly any TUNEL-positive nuclei were present in nonischemic control conditions, whereas the induction of ischemia and reperfusion increased the number of TUNEL-positive nuclei (ischemia-control). Compared with solvent control treatment, cyclopamine treatment further increased the number of TUNEL-positive nuclei—and thus apoptotic cells.
Revascularization.
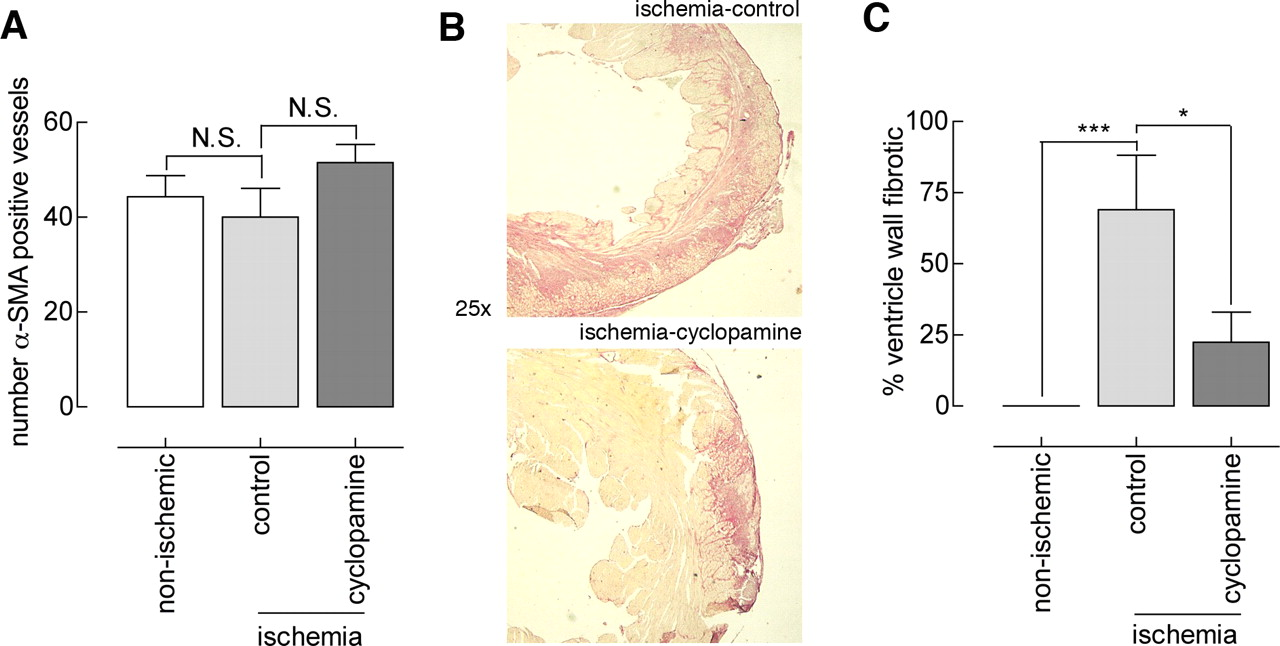
Another feature thought to be at least partly responsible for the salvaging effect of exogenous Shh in myocardial ischemia is increased neovascularization of the myocardium. To assess the role of endogenous Shh in vascularization after ischemia-reperfusion–induced injury, heart sections were immunohistochemically stained for α-SMA (Fig. 4A), and positively stained vessels were counted. Solvent-treated animals had α SMA-positive vessels throughout the heart. Cyclopamine treatment did not reduce the number of α SMA-positive vessels; this result indicated that endogenous Shh does not induce vascularization after ischemia and reperfusion. The number of α SMA-positive vessels in both groups did not differ from that in the nonischemic control group. It should be noted that this analysis did not include capillaries.
Fibrosis.
In the study performed by Kusano et al., exogenous Shh reduced cardiac fibrosis, which was suggested to be responsible for limiting ischemia-reperfusion–induced injury (7). To assess the role of fibrosis in the beneficial effect of cyclopamine treatment in our model, we evaluated fibrosis in the left ventricle by Sirius Red staining. This analysis included any present scar tissue. As shown in Figure 4C and D (quantitation), the left ventricle of solvent-treated animals contained large fibrotic lesions; the left ventricle of nonischemic controls did not contain any fibrotic lesions (Fig. 4D). Evidently, cyclopamine treatment dramatically reduced left ventricular fibrosis. Although preventing (or reducing) fibrosis is clearly beneficial in maintaining ventricular function, our data do not allow conclusions to be made about the cause or consequence of reduced fibrosis in improving left ventricular integrity by cyclopamine treatment.
Discussion
The developmental Hh pathway has previously been shown to be activated in ischemic tissue, and exogenously driven Shh expression exerts a protective effect in ischemia models (5–7). However, these studies did not address the role of endogenously expressed Shh. To investigate the role of endogenous Shh, we employed the Hh pathway inhibitor cyclopamine in a model of acute myocardial ischemia and reperfusion. Although a deleterious effect of cyclopamine treatment was expected, we instead observed a beneficial effect of cyclopamine given just before ischemia; this finding indicates that endogenous Hh might be deleterious during the early phase of myocardial ischemia and reperfusion.
Exogenous Shh has been shown to reduce ischemia-reperfusion–induced apoptosis, and the diminished apoptosis has been suggested to be partly responsible for the protective effect of exogenous Shh (7). In accordance with Shh limiting apoptosis, we show that the inhibition of an endogenously activated Hh pathway by cyclopamine aggravates apoptosis. As both exogenous Shh (and thus diminished apoptosis) as well as cyclopamine (and thus increased apoptosis) seem to limit ischemia-reperfusion–induced myocardial injury, it seems unlikely that apoptosis is a major determinant in myocardial injury. However, one should realize that apoptosis induced by cyclopamine is mechanistically different from Shh-induced inhibition of apoptosis.
Cyclopamine-induced apoptosis, well known to limit cancer cell growth, is dependent on inactivation of downstream Hh pathway components (15). In contrast, apoptosis inhibited by Shh is directly dependent on Ptch1, which serves as a dependence receptor, and does not necessarily involve downstream pathway signaling (16).
The beneficial effect of cyclopamine on myocardial injury as assessed by echocardiography was unambiguous, but we were unable to conclusively identify the underlying mechanism responsible. Cyclopamine did not significantly affect vascularization. Considering the relatively short time frame of the experiment and the fact that neovascularization is thought to be more important in chronic models of myocardial ischemia, our observation might not be that surprising (7).
Our study has some limitations that should be kept in mind when interpreting our data. For instance, the model we use did not allow us to discriminate between the effects of cyclopamine on acute myocardial injury and the effects on subsequent remodeling. Future studies in which the effect of cyclopamine on ischemia-reperfusion–induced myocardial damage is assessed at different time points should determine whether cyclopamine exerts its beneficial action in the acute or remodeling phase. Furthermore, we did not determine the area at risk (i.e., the ischemic area) in the individual animals. It is therefore difficult to formally exclude that cyclopamine-treated mice accidentally had smaller infarctions and thereby exhibited better postischemic functional recovery. However, the LAD coronary artery ligation was performed by a single experienced technician in a blinded fashion, minimizing variation in the area at risk (and infarct size), and we feel confident that infarct sizes were similar between groups.
Interestingly, the fibrosis data might shed additional light on the potential beneficial effect of apoptosis induction by cyclopamine. One could argue that the induction of apoptosis keeps fibroblast proliferation in check, thereby limiting ischemia-reperfusion–induced fibrosis and consequently limiting myocardial damage. Future experiments should test the validity of this hypothesis.
Whatever the mechanism responsible for the protective effect of cyclopamine, it is fascinating to speculate about the clinical implications of this finding. Systemic administration of an Hh-inhibitory molecule might be a more practical treatment option than the previously utilized injection of an Shh-expressing plasmid directly into the myocardium. As systemic administration of cyclopamine has so far not been successful in humans, (pro-)vitamin D3, which is well tolerated at high doses, might be an attractive therapeutic option (17). However, before such claims are made, studies addressing the actual potential of (pro-)vitamin D3 in experimental myocardial ischemia need to be performed. Currently, we are planning these experiments in addition to experiments in which Hh inhibitors are administered after induction of ischemia, which should more closely mimic the clinical situation.
In conclusion, we show that cyclopamine treatment limits myocardial ischemia-reperfusion–induced injury. Thus, Hh seems to exert a dualistic action in cardiac ischemia in which high exogenous levels are able to foster tissue repair and endogenous Hh seems to aggravate ischemic disease.
Hh pathway activity in the ischemic heart and inhibition by cyclopamine. (A) Hh pathway activity in sections of control and ischemic myocardium was analyzed by staining for the Hh pathway receptor Ptch1 (staining is denoted by arrows). Increased Ptch1 levels, indicating elevated Hh pathway activity, are seen in left ventricular tissue after 24 hrs of reperfusion that followed 1 hr of ischemia. Indicated is the original magnification. (B) Kidneys of control-treated and cyclopamine-treated animals were homogenized and immunoblotted for analysis of Ptch1 and β-actin. Densitometry was performed for all samples, and cyclopamine-treated animals that did not show diminished Ptch1 levels were considered to be nonresponders. A color version of this figure is available in the online journal. Preoperative and postoperative echocardiography. (A) Preoperative functional parameters of the left ventricle as determined by echocardiography were similar between solvent control–treated and cyclopamine-treated animals. Abbreviations: LVSs, systolic left ventricle wall thickness (cm); LVSd, diastolic left ventricle wall thickness (cm); LVIDs, systolic left ventricle internal diameter (cm); LVIDd, diastolic left ventricle internal diameter (cm); FS, fractional shortening (%); SV, stroke volume (μ l × 1000); EF, ejection fraction. (B) Postoperative functional parameters of the left ventricle showed improved outcome for cyclopamine-treated animals. The symbol * indicates P < 0.05. (C) Relative values calculated from data in (A) and (B) (the postoperative value divided by the preoperative value) confirmed the beneficial effect of cyclopamine. The symbol ** indicates P < 0.01. (D) Staining of sections with hematoxylin and eosin confirmed left ventricular dilatation in control-treated animals, which was not seen in nonischemic animals; left ventricular dilatation was strongly ameliorated in cyclopamine treated animals. A color version of this figure is available in the online journal. Immunohistochemical analysis of apoptosis after myocardial ischemia and reperfusion. (A) Hearts were collected and subjected to immunohistochemical analysis for active caspase-3. Shown are similar sections of left ventricle; caspase-3 activity in cyclopamine-treated animals was greater than that in control-treated and nonischemic mice. (B) Quantitation of active caspase-3 DAB staining in the left ventricle as performed by using the histogram function in Photoshop. Units on the y axis represent mean intensity in sample areas of a fixed size for all sections. The symbol *** indicates P < 0.005. (C) The method was similar to that described for (A) , but TUNEL staining was used. Shown are similar sections of left ventricle; the number of TUNEL-positive nuclei in the cyclopamine group is greater. (D) Quantitation of the number of TUNEL-positive nuclei per optical field (three fields counted in each left ventricle per animal, n = 4). The symbol * indicates P < 0.05. A color version of this figure is available in the online journal. Immunohistochemical analysis of vascularization and fibrosis after myocardium ischemia and reperfusion. (A) Hearts were stained for α-SMA, and positive vessels were counted. No difference in vessel number was observed between the groups. (B) Fibrosis of the left ventricle in ischemic solvent control–treated or ischemic cyclopaminetreated animals. (C) Quantitation of the average ratio of fibrotic area to left ventricular area showed decreased fibrosis in cyclopamine-treated animals after ischemia and reperfusion. The symbol * indicates P < 0.05; the symbol *** indicates P < 0.005. A color version of this figure is available in the online journal.

Footnotes
This work was funded by STW Technology Foundation.