
Abstract
Activation of T cells is a critical event in the pathogenesis of concanavalin A (Con A)-induced liver injury, and facilitating apoptosis of activated T cells may provide a strategy for the treatment. Here, we found that the ethanol extract from the stem parts of Dregea volubilis (DVE) inhibited cell proliferation and induced apoptosis, which was selective for Con A-activated, rather than nonactivated, lymph node cells. Administration of DVE prevented mice from Con A-induced elevation of serum transaminases, liver necrosis and increased TNF-α, IFN-γ, IL-2 and IL-4 in mice sera. DVE also caused apoptosis of in vivo activated T cells. In addition, increased active fragments of caspase-3 were found in the DVE-treated cells. But DVE-induced apoptosis was Fas-independent, as it was still observed in T cells from Fas ligand-mutated gld/gld mice. These results suggest that DVE may have great potential to treat T cell-mediated diseases through facilitating apoptosis of activated T cells.
Introduction
T cell-mediated immunity is one of the dominant causes in a variety of liver diseases involving autoimmune and viral hepatitis (1). Increasing evidence suggests that activation of T cells is a critical initial step in the pathogenesis of liver damage (2, 3). In mice, T cell-dependent hepatitis and acute liver injury can be modeled using in vivo administration of concanavalin A (Con A), a plant lectin and T cell mitogen. This mitogen induces polyclonal T cell activation in vitro and causes severe immune-mediated hepatitis characterized by increased serum levels of transaminases and infiltration of T cells and eosinophils into the liver (4–6). Upon Con A injection, increased TNF-α, IFN-γ, IL-2, IL-4, and IL-12 expression are detectable in the circulation, and these cytokines are known to be produced by activated T cells, macrophages and NKT lymphocytes. Nude mice, which lack immunocompetent T lymphocytes, were protected from Con A-induced liver injury, whereas lymph node cell transfer from responder mice resulted in susceptibility to Con A in the nude mice (7). Loss of the adhesion molecule LFA-1 on activated hepatic T cells also caused resistance to Con A-induced hepatic damage (8). In addition, selective immunosuppression by cyclosporine A or FK506 that inhibits IL-2 signaling in T cells prevented the liver injury (9). All these findings indicate that activated T cells play a pivotal role in the liver injury.
Apoptosis is an essential mechanism used to remove activated T cells during shutdown of an immune response and to maintain immune homeostasis (10). It is thought that to reduce or even eliminate the unwanted activated T cells through facilitating apoptosis may provide a strategy for the treatment of T cell-dependent diseases. The advantage of this approach is to cause the immunosuppression that is specific for pathogenic T cells before they function as effector cells, without damaging the normal immunocompetence.
During screening of a variety of plant sources, it was found that the ethanol extract prepared from stem parts of Dregea volubilis (DVE) had potent immunomodulatory activities. Dregea volubilis, belonging to the family Asclepiadaceae, is a tall woody climber distributed in various countries including the southern area of China. The leaves are edible and used as a green vegetable, while the extract from the whole plant or stem parts has been used traditionally as an anti-inflammatory medicine to treat several diseases including eye ailments, tracheitis and stomachache (11, 12). Nevertheless, the mechanism underlying is still unclear. In the present study, DVE was found to selectively induce apoptosis of activated T cells in a caspase-dependent, but Fas-independent manner. The effects of DVE contributed to the improvement of Con A-induced liver injury.
Materials and Methods
Animals and Reagents.
Eight- to ten-week-old male BALB/c and C57BL/6 mice were purchased from the Experimental Animal Center of Jiangsu Province (Jiangsu, China). Eight-week-old male gld/gld mice with a C57BL/6 background were provided by Professor Zichun Hua. Mice were maintained with free access to pellet food and water in plastic cages at 21 ± 2°C and kept on a 12-hr light/dark cycle. Animal welfare and experimental procedures were carried out in accordance with the Guide for the Care and Use of Laboratory Animals (Ministry of Science and Technology of China, 2006) and the related ethical regulations of Nanjing University. Anti-mouse antibodies against CD3 (clone145-2C11), CD28 (clone 37.51), CD25 (clone 7D4), Fas ligand (clone MFL3) and Fas (clone Jo2) were purchased from BD Pharmingen (Becton Dickinson, San Diego, CA).
Preparation of Dregea volubilis Extract.
The stems of Dregea volubilis (Asclepiadaceae) were collected in November 2006 from the province of Yunnan, China. The plant was identified and authenticated by associate professor Chaofeng Zhang. A voucher specimen (No. 200612A) was deposited at China Pharmaceutical University. The stems were harvested and air dried at room temperature in shade. Each 1 kg of the stems was pulverized and extracted three times with 6 liters of 70% ethanol. The ethanol was removed under reduced pressure by rotary evaporation and the water residue was removed by lyophilization. The yield was 9.4%. For testing, the extract was dissolved in dimethyl sulfoxide (DMSO) and diluted to the desired concentrations in RPMI 1640 medium (GIB-COBRL, NY). DMSO final concentrations were kept to below 0.1% and did not exert any detectable change in the cell growth or apoptosis. The test samples of DVE were 0.2 μm sterile filtered (Millipore, Billerica, MA) and the presence of endotoxins was analyzed using the Limulus assay (kinetic gel-clot assay; ACC Inc.). All endotoxin levels were below 0.25 EU/mL in the test samples. We have tested multiple batches of the material and obtained similar results for each experiment.
Cell proliferation.
Lymph node cells isolated from mice were activated by Con A (5 μg/ml) for 24 hrs, which were indicated as activated cells, while those without Con A stimulation were used as nonactivated cells. The cells were further incubated in 96 well-plate with or without various concentrations of DVE at a density of 5 × 105 cells per well for 12 hrs. For MTT assay, 20 μl of MTT (Sigma, MO; 4 mg/ml in PBS) was added per well 4 hrs before the end of the incubation. MTT formazan production was dissolved by DMSO replacing the medium. The optical density at 540 nm (OD540) was measured by a microplate reader. Cell proliferation was also assayed by incorporation of [methyl-3H] thymidine (ICN Pharmaceuticals, Costa Mesa, CA) at 0.5 μCi/well during the last 8 hrs of incubation, and the uptake was measured as counts per minute (c.p.m.).
Measurement of Apoptosis.
Cell apoptosis was determined by several methods. The morphological features of the cells after DVE treatment were observed by fluorescence microscopy after staining with DAPI (Roche, Germany) at 5 μg/ml as previously described (13). For DNA fragmentation assay, DNA preparation were obtained and processed for electrophoresis in 2% agarose gel, as described previously (14). For terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling (TUNEL) assay, In Situ Cell Death Detection Kit was used according to the manufacturer’s instruction (Roche), followed by flow cytometric analysis. For annexin V staining, the cells were measured by flow cytometry after addition of FITC-conjugated annexin V (Bender, Germany), as previously described (15). Samples were analyzed by flow cytrometry on a FACScan (Becton Dickinson). Nuclear fragmentation and/or marked condensation of the chromatin with reduction of nuclear size were considered as typical features of apoptotic cells.
Induction of Con A-Induced Liver Injury and Treatment.
Mice received an intravenous Con A injection (20 mg/kg). Subsequently, sera were collected to measure the serum levels of cytokines and aminotransaminases at 2 and 24 hrs, respectively. Livers were also excised for the histological analysis at 24 hrs. In some experiment, spleens were obtained for apoptosis measurement at 12 hrs. For treatment, mice were administered with DVE at 150 or 300 mg/kg by oral gavage three times in an 8 hr interval before Con A injection. As the control, one injection of 30 mg/kg CsA was given intraperitoneally 16 hrs before.
Assay for Serum Transaminase Activities.
Blood samples were obtained by retro-orbital puncture. Sera were prepared by centrifugation at 4 °C and stored frozen at −70°C until use. Serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were assessed using the commercial kit as recommended by the manufacture (Nanjing Jiancheng Bio-engineering Institute, Nanjing).
Histology and Cytokine Assay.
Livers were fixed in 10% formalin, embedded in paraffin, sectioned, and stained with hematoxylin-eosin for morphological evaluation. The sections were graded for the degree of hepatocyte necrosis and inflammation in hepatic lobules. The histological changes were read on a scale of 0–4 (0, no change; 1, mild; 2, moderate; 3, severe; and 4, most severe) and expressed as an average score. A pathologist unaware of our experimental treatment evaluated the liver sections. Serum levels of cytokines were determined using Cytometric Bead Array (CBA) cytokine assay kit according to the manufacturer as recommended by BD Pharmingen.
Western Blot Analysis.
In brief, cells were washed with phosphate-buffered saline and lysed in the lysis buffer containing 50 mM TrisCl, 150 mM NaCl, 1% NP40, 100 μg/ml PMSF. After 10,000 × g centrifugation for 10 mins, the protein content of the supernatant was determined by a BCA™ protein assay Kit (Pierce, Rockford, IL). The protein lysates were separated by 10% SDS-PAGE and subsequently electrotransferred onto a polyvinylidene di-fluoride membrane (Millipore Corp., Bedford, MA). The membrane was blocked with 5% nonfat milk for 2 hrs at room temperature. The blocked membrane was incubated with the indicated antibodies. Primary antibodies used were against cleaved caspase-3 (1:1000 dilution, Cell Signaling Technology, Beverly, MA) and gapdh (1:1000 dilution, Santa Cruz Biotechnologies, Santa Cruz, CA), and the secondary antibodies were horseradish peroxidase-conjugated goat anti-mouse or anti-rabbit immunoglobulin (1:10,000, KPL, Gaithersburg, MD). Protein bands were visualized using Western blotting detection system according to the manufacturer’s instructions (Cell Signaling Technology).
Statistical Analysis.
Data are expressed as means ± SD. Statistical analyses were performed using one-way analysis of variance (ANOVA), followed by orthogonal contrasts. The data of liver histologic changes were analyzed using non-parametric test, as applicable. P < 0.05 was considered significant. In addition, the data analyzed by flow cytometry are expressed as means ± SEM.
Results
DVE Induces Apoptosis of Con A-Activated But Not Nonactivated T Cells.
Figure 1A illustrates that DVE showed a dose-dependent enhancement of non-activated cell proliferation. In contrast, cell proliferation was significantly inhibited by DVE in a dose-dependent way in Con A-activated cells (Fig. 1A and B). A similar observation was made with lymph node cells activated by anti-CD3/anti-CD28 monoclonal antibodies (data not shown). To investigate the reason of the selective inhibitory activity potentiated by DVE, apoptosis was measured by DAPI staining in fluorescence microscopy. As shown in Figure 1C, typical chromatins condensation and blebbing nuclei were only displayed in DVE-treated activated T cells. Such apoptosis was also verified by studying internucleosomal DNA fragmentation (Fig. 1D). Moreover, apoptosis of activated T cells was confirmed by the TUNEL assay. DVE dose-dependently induced apoptosis of Con A-activated T cells, which was demonstrated by the presence of DNA breaks (Fig. 1E).
DVE Prevents Mice from Con A-Induced Liver Injury by Inducing Apoptosis of Activated T Cells.
Figure 2A illustrates that the treatment of 300 mg/kg DVE significantly inhibited the elevation of serum ALT and AST in mice with Con A-induced liver injury and 150 mg/kg DVE showed a tendency to reduce the transaminases elevation, which was almost comparable to CsA. Pathological evidence of liver injury was correlated with serum ALT and AST levels (Fig. 2B), where a marked decrease in lymphocyte infiltration and massive hepatocyte necrosis was shown in 300 mg/kg DVE-treated mice compared with untreated model mice, and the mice treated with 150 mg/kg DVE or 30 mg/kg CsA showed a moderate reduction. It should be noted that less lymphocyte infiltration was exhibited in the 150 mg/kg DVE-treated mice than in those treated with CsA. For the histopathological grading, the groups treated with 150 or 300 mg/kg DVE showed a significantly lower score compared with untreated control (Table 1).
Since Con A-induced liver injury is associated with increased TNF-α, IFN-γ, IL-2 and IL-4 expression (16), CBA cytokine assay was used to determine the effects of DVE on serum cytokine levels 2 hrs after Con A injection. All serum levels of those cytokines were significantly lower in the mice treated with 150 or 300 mg/kg DVE compared with untreated control and were similar to those in the CsA-treated group (Fig. 2C).
Next, apoptosis measurement was made by Annexin V staining on spleen cells freshly isolated from the regent-treated mice 12 hrs after Con A injection. Compared with the control, the percentages of apoptotic T cells among gated CD3+ cells were increased in the groups treated with 150 and 300 mg/kg DVE (Fig. 3). In contrast, few apoptotic cells were shown among gated CD3− cells in all the groups (Fig. 3).
DVE Facilitates Apoptosis of Activated T Cells in a Caspase-Dependent, But Fas-Independent Manner.
Because activated T cells express Fas ligand (Fas L), and Fas-mediated signaling plays important parts in the induction of lymphocyte apoptosis (17), the contribution of Fas/FasL interaction in the apoptotic effect of DVE was studied. As shown in Figure 4A, FasL expression at the surface of Con A-activated lymph node cells was increased upon 12 hrs of exposure to DVE. However, apoptosis induced by DVE was not altered in lymph node cells from FasL–mutated gld/gld mice, in which apoptosis can be induced by the anti-Fas antibody (Fig. 4B). When apoptotic signaling pathway was examined by Western blotting, the active fragment of executor caspase-3 was observed to appear more intensively in DVE-treated activated lymph node cells than untreated cells (Fig. 4C). Similar result was also found in Con A- activated spleen cells, indicating that DVE increased caspase activation (Fig. 4C).
Discussion
In traditional medicine, foods from plant sources like vegetables and fruits have drawn increased attention. The present study demonstrates that DVE, extracted from the stem parts of a green vegetable, has the potential for immunomodulatory activity. While DVE stimulated the proliferation of nonactivated lymph node cells, cell proliferation was markedly and rapidly inhibited by DVE once pre-activated by Con A or anti-CD3/anti-CD28 monoclonal antibodies. Such inhibition is mostly due to the induction of apoptosis instead of cytotoxity or overgrowth, where apoptosis was achieved after 12 hrs of exposure to DVE in Con A-preactivated lymph node cells but not nonactivated lymphocytes (Fig. 1 C–E). It should be emphasized that the extract showed an increase, rather than inhibition, in the proliferation of the lymph node cells from naïve mice (Fig. 1A and B). These findings suggest that DVE selectively induces apoptosis of activated, but not nonactivated T cells.
To examine whether the DVE-induced activated T cell apoptosis identified in vitro could account for the prevention from Con A-induced liver injury, we performed an in vivo experiment. Against this liver injury, DVE showed a dose-dependent inhibition on serum transaminases as well as liver histological changes (Fig. 2A and B, Table 1). This finding suggests that the immunomodulatory activity of the extract could be used for the treatment of liver disease. In addition, DVE treatment dramatically inhibited the pathogenic cytokine production 2 hrs after Con A injection (Fig. 2C). Previous studies have indicated that the early increase in cytokine levels is pivotal in triggering liver damage, because maximal levels of most cytokines are reached before infiltrating of the activated T lymphocytes occurs, but liver damage has already begun (9, 18). TNF-α and IFN-γ directly contribute to liver damage via activation of apoptotic signaling in hepatocytes (7, 19). And IL-4 plays a role in recruiting eosinophils into liver (6). It is not possible that the reduction of these pathogenic cytokines in the DVE-treated mice resulted from a failure in activation of T cells, because DVE seems not to suppress T cell activation like CsA, supported by the evidence that the expression of active marker CD25 in T cells was not decreased by DVE in the presence of Con A (supplementary data). Apoptosis of activated T cells could provide a rational explanation because apoptosis can lead to loss or dysfunction of activated T cells that are essential in the initial step of liver damage. In the DVE-treated mice, high percentage of apoptotic CD3+ T cells was found in the spleen at 12 hrs after Con A injection (Fig. 3). In agreement with this finding, earlier studies indicated that activated T cells undergoing apoptosis were of dysfunction, including having low hepatotoxicity (20).
Fas-mediated apoptosis is known as one of mechanisms by which activated T cells undergo death (21). The sensitivity of T cells toward Fas-mediated apoptosis can be modulated by the level of FasL expression upon activation. In the current study, DVE was able to up-regulate the expression level of FasL at the surface of activated T cells (Fig. 4A). However, the increased FasL seems not to be involved in the enhancement of Fas-dependent apoptosis signaling, as potent apoptosis induced by DVE was not inhibited in the gld/gld mice deficient in FasL (Fig. 4B). Similar result was found in the study of methotrexate, which selectively induced apoptosis of mitogen-activated peripheral blood T cells through a Fas-independent pathway in spite of its property to stimulate FasL mRNA expression in activated T cells (22). Resistance to Fas-induced apoptosis is likely due to short-term exposure to T cell stimuli. In peripheral blood T cells, their sensitivity to Fas-dependent apoptosis progressively develops from day 3 to day 6 of mitogenic activation (23). Naïve CD4+ or CD8+ T cells become sensitive to Fas-induced apoptosis after approximately 6 days of TCR stimulation in vitro (24). In this study, T cells were fully susceptible to DVE-induced apoptosis only after 24 hrs of Con A stimulation. These results indicate that factors other than FasL expression control cell susceptibility to Fas-mediate apoptosis, and that in addition to the Fas system, there are other apoptotic pathways involved in DVE-induced apoptosis.
Caspases have a central role in the regulation of most types of apoptotic pathways (21). Activation of the caspase cascade results in the cleavage of cellular substrates that induces the morphological and biochemical features of apoptosis. Caspase-3 locates in the downstream of the caspase cascade and functions as a key executor upon activation. In the present study, exposure to DVE increased the amount of the active fragment of caspase-3 in activated T cells (Fig. 4C), suggesting that DVE facilitates apoptosis of activated T cells likely through promoting caspase activation with irrelevance of Fas-dependent pathway.
There are many plants and plant products discovered with immunomodulatory activity such as some triterpenoids (25). However, little attention has been given to the mechanism underlying them. In our study, the bioactivities of DVE indicated its immunomodulatory potential. DVE tended to stimulate nonactivated lymph node cells while suppressing already activated T cells by selectively facilitating apoptosis through a caspase-dependent, but Fas-independent pathway. This facilitation of apoptosis was suggested to account for DVE efficiency in T cell-mediated Con A-induced liver injury. So far, Dregea volubilis has been subjected to phytochemical analysis and found to mainly contain triterpenoid glycosides and aglycones, called dregeosides and drevogenins, respectively (26). It has been recently reported that Drevogenin-D, one of the aglycones, prevented selenite-induced oxidative stress and calpain activation in cultured rat lens (27). Further studies are in progress to identify the functional compounds responsible for the immunomodulatory activity of DVE.
Effect of DVE on Liver Histology a
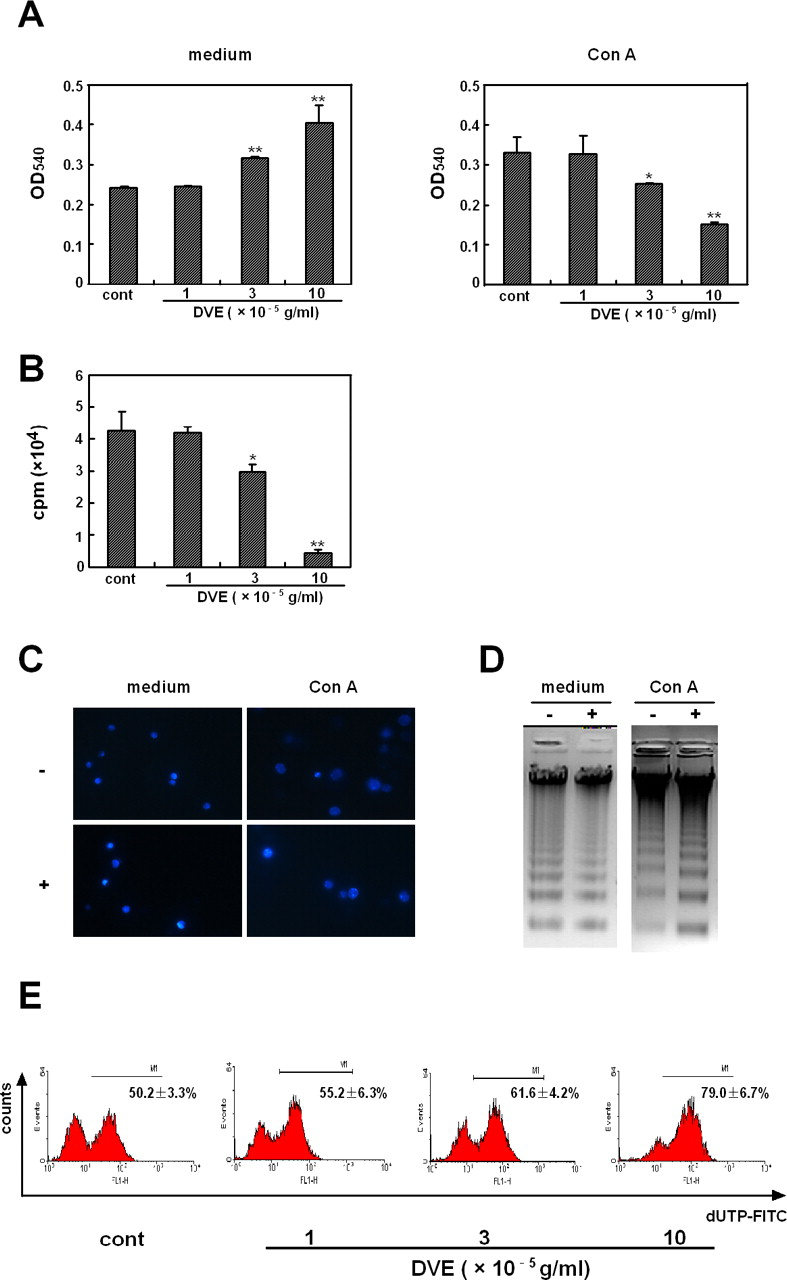
DVE selectively induced apoptosis of Con A-activated T cells. Lymph node cells isolated from BALB/c mice were incubated in medium or in the presence of Con A for 24 hrs. Then the cells were further incubated with or without various concentrations of DVE for 12 hrs. (A) MTT method was used to detect cell proliferation. (B) Incorporation of [3H] thymidine was used to assay the proliferation of Con A-activated cells. Values are shown as means ± SD of three independent experiments. *P < 0.05, **P < 0.01 vs the control. (C) Cells were stained with DAPI and examined by fluorescence microscopy (medium and Con A, without (−) or with (+) 10 × 10−5 g/ml of DVE). Original magnification, ×400. (D) DNA fragmentation was evaluated by electrophoresis. (E) DNA strand breaks were detected by TUNEL assay. Numbers in each histogram refer to the percentage of cells above the threshold of normal controls, and are shown as means ± SEM of three independent experiments. A color version of this figure is available in the online journal.

Effects of DVE on Con A-induced liver injury. Liver injury was induced by intravenous Con A injection (20 mg/kg). Mice were administered with DVE at 150 or 300 mg/kg by oral gavage three times in an 8 hr interval before Con A injection. As the control, one injection of 30 mg/kg CsA was given intraperitoneally. Serum transaminase activities and liver histologies were assessed at 24 hr and serum cytokine levels were analyzed at 2 hr after Con A injection. (A) Serum levels of ALT and AST. (B) Representative photographs of H&E-stained liver sections. a, Normal; b, Model; c, 150 mg/kg DVE; d, 300 mg/kg DVE; e, 30 mg/kg CsA. Original magnification ×100. (C) Serum levels of TNF-α, IFN-γ, IL-2 and IL-4. Values are shown as means ± SD, n ≥ 8 per group. ## P < 0.01 vs Normal; *P < 0.05, **P < 0.01 vs Model. A color version of this figure is available in the online journal.

DVE induced apoptosis of in vivo activated T cells. Mice were treated as described in Figure 2. Spleens were obtained 12 hrs after Con A injection, and percentages of apoptotic CD3+T cells and CD3− cells were determined by Annexin V binding. Numbers in each histogram are shown as means ± SEM of three independent experiments.
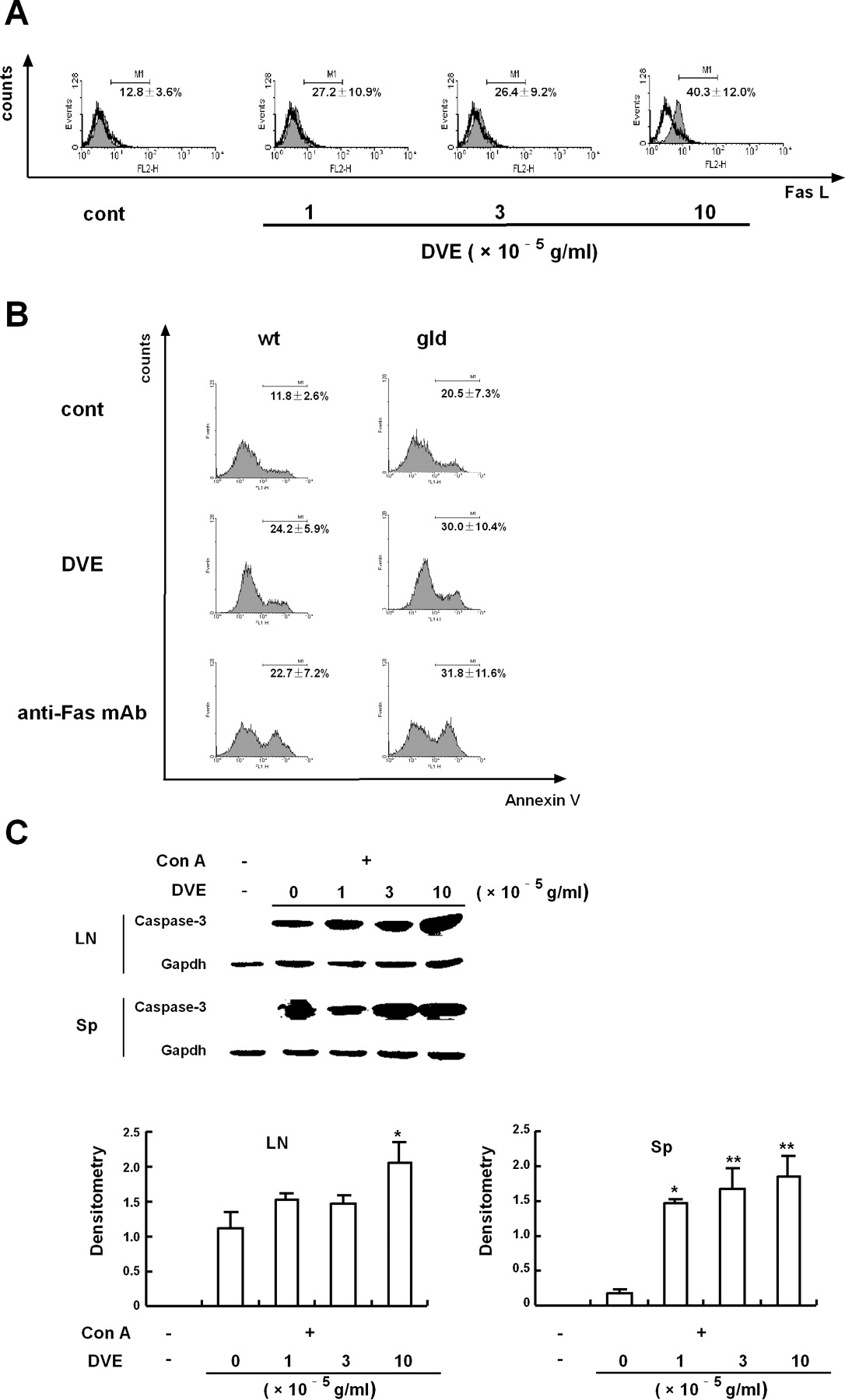
DVE induced apoptosis of activated T cells in a Fas-independent, but caspase-dependent way. (A) Lymph node cells were activated by Con A (5 μg/ml) for 24 hrs, and then incubated with various concentrations of DVE for 12 hrs. FasL expression on the surface of the cells was evaluated by flow cytometry. Open histogram, isotype antibody; filled histogram, anti-FasL PE. (B) Lymph node cells isolated from C57BL/6 wide type mice (wt) and gld/gld mice (gld) were activated by Con A (5 μg/ml) for 24 hrs. Then the cells were exposed to DVE (10 × 10−5 g/ml) or anti-Fas monoclonal antibody (1 μg/ml) for 12 hrs. Percentages of apoptotic T cells were determined by Annexin V binding. Numbers in each histogram are shown as means ± SEM of three independent experiments. (C) Cells were treated as described in Figure 4A. The expression of active fragment of caspase-3 were determined by Western blot analysis (upper). Gapdh was used as a loading control. The relative expression of active fragment of caspase-3 was quantified densitometrically using the software LabWorks 4.0, and reported as fold relative to the reference band of gapdh (lower). Values are shown as means ± SD of three independent experiments. *P < 0.05, **P < 0.01 vs the activated control (without DVE).
Footnotes
This study was supported by National Natural Science Foundation of China (No. 30730107 and 30701025).