
Abstract
Magnolol, an active component extracted from Magnolia officinalis, has been reported to have protective effect on ischemia and reperfusion (I/R)-induced injury in experimental animals. The aim of the present investigation was to further evaluate the mechanism(s) by which magnolol reduces I/R-induced myocardial injury in rats in vivo. Under anesthesia, left anterior descending (LAD) coronary artery was occluded for 30 min followed by reperfusion for 24 h (for infarct size and cardiac function analysis). In some experiments, reperfusion was limited to 1 h or 6 h for analysis of biochemical and molecular events. Magnolol and DMSO solution (vehicle) were injected intra-peritoneally 1 h prior to I/R insult. The infarct size was measured by TTC technique and heart function was monitored by Millar Catheter. Apoptosis related events such as p-ERK, p-Bad, Bcl-xl and cytochrome c expression were evaluated by Western blot analysis and myocardial caspase-3 activity was also measured. Magnolol (10 mg/kg) reduced infarct size by 50% (P < 0.01 versus vehicle), and also improved I/R-induced myocardial dysfunction. Left ventricular systolic pressure and positive and negative maximal values of the first derivative of left ventricular pressure (dP/dt) were significantly improved in magnolol-treated rats. Magnolol increased the expression of phosphor ERK and Bad which resulted in inhibition of myocardial apoptosis as evidenced by TUNEL analysis and DNA laddering experiments. Application of PD 98059, a selective MEK1/2 inhibitor, strongly antagonized the effect of magnolol. Taken together, we concluded that magnolol inhibits apoptosis through enhancing the activation of ERK1/2 and modulation of the Bcl-xl proteins which brings about reduction of infarct size and improvement of cardiac function in I/R-induced injury.
Introduction
Ischemic heart disease is the single-most important cause of death in developed countries. Accumulating evidence indicates that apoptosis, a programmed cell death, contributes significantly to post-ischemic cardiomyocyte death, suggesting that therapeutic interventions that inhibit apoptotic cell death may attenuate ischemia and reperfusion (I/R)-induced heart injury (1). Despite significant therapeutic advances, I/R-induced myocardial injury is still a major unsolved problem. Therefore, cardiac myocyte apoptosis may be a good target for therapeutic modulation in the context of myocardial infarction and I/R injury.
In mammalian cells, so far three major mitogen-activated protein kinases (MAPKs) signaling pathways, including extracellular signal-related protein kinases (ERK1/2), p38 MAPK and c-Jun NH2-terminal protein kinases (JNKs)/stress-activated protein kinases (SAPKs) have been identified (2) and they are activated in response to myocardial I/R, and the activations of ERK1/2 (beneficial) and p38 MAPKs-JNKs (deleterious) exert opposite effects on post-ischemic myocardial apoptosis and cardiac function recovery (3, 4).
Magnolol has attracted great attention for protection on I/R-induced injury in many organs including myocardium in experimental animals (5, 6). However the protective molecular mechanism(s) of magnolol on I/R-myocardium, in particular, related to MAP kinase activation, remains unknown at the present time. Therefore, the aims of the present study were to determine whether administration of magnolol may activate ERK1/2 in myocardial I/R rat in vivo, and this may be responsible for the reduction of infarct size and improvement of myocardial function.
Materials and Methods
Experimental Animals.
Male Sprague-Dawley rats, 7–8 weeks of age (body weight 200–250 g; Samtako Inc., Osan, Korea) were used for the study. All animals were maintained in accordance with the Guide for the Care and Use of Laboratory Animals, published by the US National Institutes of Health (NIH publication NO. 85–23, revised in 1996). The protocol was approved before the animal study by Animal Research Committee of the Gyeongsang National University, Korea.
Experiment Protocol.
The animals were randomly divided into two sets of experiment each consisting of four groups. In the first set of experiments, animals were divided as follows: (1) Sham group (Sham), sham-operated rats in which no tightening of the left anterior descending (LAD) coronary artery sutures was performed (n = 6). (2) I/R control group (I/R), rats pretreated with vehicle (DMSO 0.25 ml, intraperitoneal injection 1 h before ischemia, n = 10). (3) I/R + magnolol 5 mg/kg group (I/R + Mag 5), rats were given magnolol at a dose of 5 mg/kg (intraperitoneal injection 1 h before ischemia, n = 10). (4) I/R + magnolol 10 mg/kg group (I/R + Mag 10), rats were given magnolol at a dose of 10 mg/kg (intraperitoneal injection 1 h before ischemia, n = 10). The animals were exposed to 30 min ischemia followed by 24 h of reperfusion after which infarct size and heart functions were estimated. In the second set of experiments, animals were divided as follows: (1) Sham group (Sham, n = 18). (2) I/R control group (I/R, n = 18). (3) I/R + Mag 10 mg/kg group (I/R + Mag 10, n = 18). (4) I/R + Mag 10 mg/kg + PD 98059 group (I/R + Mag 10 + PD, n = 18), rats were given PD 98059 at a dose of 1 mg/kg (intravenous injection 10 min before ischemia). The animals were exposed to 30 min ischemia followed by 1 h or 6 h of reperfusion after which tissue samples were used for Western blotting and apoptosis assay, respectively. The dose of magnolol (10 mg/kg) was selected from preliminary experiments. When used from 2 to 20 mg/kg (n = 6–10/dose), magnolol exerted direct negative inotropic effects (+ dP/dt: −21 ± 2%, P < 0.01) at higher concentrations such as 20 mg/kg.
Surgical Preparation.
The animals were anesthetized with ketamine (100 mg/kg, i.m.) and xylazine (10 mg/kg, i.m.), intubated through a midline tracheostomy and mechanically ventilated using a positive pressure respirator for small animals (Type 7025, Ugo Basile, Varese, Italy; 60–80 strokes/min, 8 ml/kg body weight) at a rate adjusted to keep blood pH and blood gases within normal range. The chest was opened via a left thoracotomy, followed by a pericardiotomy. A 4–0 black silk suture was passed around the LAD coronary artery, and the ends were pulled through a small vinyl tube to form a snare and then tightened. After 30 min of ischemia, the myocardium was reperfused by loosening the snare for 1 h or 6 h or 24 h. The body temperature was measured by an electric thermistor placed in the rectum and maintained at 37°C by a heating pad placed under the rats. Sham-operated rats were subjected to the same procedures without LAD occlusion. After surgery all animals were housed in cages and maintained on a 12 h night/day cycle and allowed free access to food and water at all times until the experiments began.
Hemodynamic Measurements.
Twenty-four hours after reperfusion, rats were anesthetized with ketamine (75 mg/kg, i.m.) and xylazine (7.5 mg/kg, i.m.). The right common carotid artery was cannulated with a 2 F Millar Catheter (Millar Instruments, Houston, TX), into the ascending aorta to measure systolic and diastolic blood pressure (SBP, DBP), mean arterial pressure (MAP) and heart rate (HR). The pressure transducer was then advanced into the left ventricle (LV) to measure LV systolic (LVSP), end-diastolic (LVEDP) pressure, and rate of rise and decline of LV pressure (±dP/dt).
Determination of Area at Risk and Infarct Size.
After measurements of heart function, before harvesting the hearts, the LAD was retired. Two ml of 1% Evans blue dye was injected into the femoral vein root to stain the normally perfused region blue and outline the area at risk. Right ventricle and atrium tissues were removed, and the LV was cut into transverse slices. The area at risk was separated from the nonischemic zone and incubated for 20 min at 37°C in a 1% solution of triphenyltetrazolium chloride (TTC) to visualize the infarct area. The infarct area (pale, white arrows) from the non-necrotic (red) area at risk was differentiated (Fig. 1). The area at risk as a percent of the LV mass (area at risk, %) and the infarct area as a percent of the area at risk mass (infarct size, %) were calculated.
Western Blotting.
Total proteins were extracted from area at risk zones of the heart. Western blot assay of Bcl-xl, Bad, phosphorylated Bad (p-Bad), extracellular signal-regulated kinase (ERK), and p-ERK proteins were performed as below. Briefly, 1 h after reperfusion (for the ERK and p-ERK) or 6 h after reperfusion (for the Bcl-xl, Bad, p-Bad, and cytochrome c) myocardial tissues were placed in lysis buffer containing protease inhibitors, homogenized and then centrifuged. After determining the concentration of protein in each sample using a modified Bradford assay, 50–80 μg of total protein was boiled in loading buffer and then loaded onto gradient SDS-polyacrylamide gels. Following electrophoresis, the proteins were transferred to an immobilon-polyvinylidine fluoride (PVDF) membrane. Non-specific binding sites on the membranes were blocked with 5% (w/v) nonfat milk in TBS-T (20 mM Tris-HCl, pH 7.5, 137 mM NaCl, 0.1% (v/v) Tween 20) for 60 min at room temperature. Subsequently, the membranes were incubated with a mouse monoclonal antibody recognizing p-ERK, Bcl-xl and Bad (Santa Cruz Biotechnology, Monterey, CA), a rabbit polyclonal antibody recognizing ERK1/2 and goat polyclonal antibody recognizing p-Bad (Santa Cruz Biotechnology) according to the manufacturer’s instructions. Bound antibody was detected by horseradish peroxidase conjugated anti-mouse IgG, anti-rabbit IgG and anti-goat IgG. Finally, enhanced chemiluminescence (ECL) detection reagents were employed to visualize peroxidase reaction products. Blots were quantified by laser scanning densitometry.
Measurement of Caspase-3 Activity.
Activity of caspase-3 was evaluated by using a colorimetric activity assay kit (Chemicon International Molecular, Temecula, CA) according to the manufacturer’s instructions. Myocardial tissue specimens were obtained from the area at risk zones. The sample was homogenized in ice-cold buffer provided in this kit and centrifuged. After determining protein concentration, the supernatant was incubated with caspase-3 substrate (Ac-DEVD-pNA) on a 96-well-plate. Activity of caspase-3 was determined using a spectrophotometer (Lambda EZ 210, Perkin-Elmer Inc., Boston, MA) at 405 nm, and the results were expressed as fold increase over the mean value of sham.
DNA Isolation and Gel Electrophoresis.
The appearance of DNA laddering was detected using agarose gel electrophoresis. Briefly, freshly frozen tissue samples (30–40 mg each, after 6 h reperfusion) from area at risk zones were minced in 500 μl lysis buffer (Puregene DNA isolation kit; Gentra Systems, Minneapolis, MN). Homogenized tissue samples were digested with a 100 μg/ml of proteinase K at 56°C for 2 h and incubated with RNase A at 37°C for 1 h. After incubation, tissues were precipitated and centrifuged at 13,000 g for 5 min. Supernatant containing DNA was precipitated with isopropanol. After centrifugation at 13,000 g for 5 min, the resulting DNA pellets were washed with 75% ethanol and dissolved in DNA hydration solution (Promega, Madison, WI). The content of DNA was detected at 260 nm by spectrophotometry (Lambda EZ 210), for which 10 μg DNA was loaded into 1.8% agarose gel containing 0.5 μg/ml ethidium bromide. DNA electrophoresis was carried out at 80 V for 1 h. DNA ladders, an indicator of tissue apoptotic nucleosomal DNA fragmentation, were visualized and photographed under ultraviolet transillumination.
Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick End Labeling (TUNEL) Assay.
Hearts were isolated from each group (after 6 h reperfusion) for the TUNEL assay. After the blood and the fixation were washed out, the heart was sliced transversely in 2.5 mm thicknesses. Paraffin-embedded, 3 μm-thick myocardial sections were used as described previously (7). Immunohistochemical procedures for detecting apoptotic cardiomyocytes were performed by using a TUNEL kit (Roche Applied Science, Indianapolis, IN). The slides were covered with the mounting medium containing DAPI for detection of total nuclei. Three sections from each myocardial sample were randomly selected and ten microscopic fields per section were evaluated. In each field, nuclei were counted and the percentage of TUNEL-positive nuclei was calculated (number of apoptotic myocytes/the total number of myocytes counted × 100), and the assays were performed in a blinded manner. Fluorescence staining was viewed with a fluorescent microscope (Fluoview 500, Olympus Optical Co. Ltd., Tokyo, Japan). Heart tissue samples for TUNEL analysis were obtained from the margins of non-infarcted but ischemic areas.
Chemicals.
The method of isolation and purity of magnolol from Magnolia officinalis was previously described by us (8) and it was dissolved in 0.1% DMSO.
Statistical Analysis.
All data were expressed as mean ± standard error of the mean (SEM). Differences between groups were assessed by one-way ANOVA followed by Student-Newman-Keuls test. Values of P < 0.05 were considered to be statistically significant.
Results
Magnolol Improves Cardiac Function in I/R Heart.
Hemodynamic parameters were measured in order to know whether magnolol improves myocardial function in I/R rats. At the end of reperfusion (24 h), I/R resulted in substantial left ventricular (LV) dysfunction as evidenced by increased left ventricular end diastolic pressure (LVEDP), decreased left ventricular systolic pressure (LVSP) and positive and negative maximal values of the first derivative of left ventricular pressure (±dP/dt) (Table 1). Magnolol significantly inhibited the decrease of LVSP and ±dP/dt (98 ± 2 vs. 85 ± 2 mmHg, P < 0.01; 3623 ± 89 vs. 3223 ± 104 mmHg/s, P < 0.05; −3010 ± 83 vs. −2579 ± 106 mmHg/s, P < 0.05; respectively). Moreover, the elevated LVEDP was significantly attenuated in the I/R + Mag 10 group (9 ± 1 vs. 6 ± 1 mmHg, P < 0.05). Consequently, magnolol significantly recovered the heart functions after I/R (Table 1). No significant difference of heart rates (HR) was found in all groups (data not shown). When the effect of vehicle and magnolol on normal hearts under the sham-operated condition was tested, there was no significant difference in the values of HRs and mean arterial pressures (MAPs) among the vehicle and Mag 10 groups. Furthermore, HRs and MAPs did not change significantly throughout the experimental period in the two groups (Table 2).
Magnolol Reduces Infarct Size.
To explore whether magnolol reduces infarct size, we measured area at risk and infarct size. As shown in Table 3, no significant differences were found in area at risk (% of total LV) among the groups. In the I/R group, the infarct size was 58% ± 3%, and that of the I/R + Mag 5 group was 48% ± 2% (P = 0.08 compared with the I/R group). However, the I/R + Mag 10 group the infarct size was significantly reduced to 29% ± 2% (P < 0.01 compared with the I/R group).
ERK1/2 Phosphorylation and Bcl-2 Family Protein Expression.
To investigate the effect of magnolol related with apoptosis, we measured apoptosis-related gene expression and related signals by Western blot analysis. The phosphorylation level of ERK1/2 was also determined by immunoblotting. As shown in Figure 2A, treatment with magnolol resulted in a 9.5 ± 0.6 fold (relative to sham) increase in ERK phosphorylation. Cotreatment with PD 98059 blocked ERK phosphorylation induced by magnolol (9.5 ± 0.6 vs. 6.1 ± 0.4 fold relative to sham, P < 0.01). There was no difference in total ERK, p-Jun N-terminal Kinase (JNK), p-JNK, p38 and p-p38 among the groups studied (data not shown). Western blot analysis of p-Bad, Bcl-xl and cytocrome c protein is shown in Figure 2A and 2B. I/R injury obviously reduced p-Bad and Bcl-xl expression (0.2 ± 0.1, 0.3 ± 0.1; respectively) and increased cytochrome c expression (9.0 ± 0.5) compared to sham as measured densitometrically (fold relative to sham). In contrast to the I/R control group, magnolol (10 mg/kg) significantly increased p-Bad and Bcl-xl expression (0.2 ± 0.1 vs. 0.5 ± 0.1, P < 0.05; 0.3 ± 0.1 vs. 0.7 ± 0.1, P < 0.05) but decreased cytochrome c (9.0 ± 0.5 vs. 2.0 ± 0.2, P < 0.01). Cotreatment with PD 98059 also significantly decreased p-Bad and Bcl-xl expression (0.5 ± 0.1 vs. 0.3 ± 0.1, P < 0.05; 0.7 ± 0.1 vs. 0.5 ± 0.1, P < 0.05; respectively) but increased cytochrome c expression (2.0 ± 0.2 vs. 6.0 ± 0.4, P < 0.05) compared with I/R + Mag 10 group. There was no difference in total Bad expression among the groups (data not shown).
Anti-Apoptotic Effect of Magnolol.
To further explore anti-apoptotic effect of magnolol, we measured caspase-3 activity, DNA strand break, and TUNEL analysis. As shown in Figure 3, treatment with magnolol (10 mg/kg) significantly reduced caspase-3 activity compared to I/R control group (1.8 ± 0.1 vs. 2.8 ± 0.2 fold relative to sham, P < 0.01), which was again reversed in I/R + Mag 10 + PD group (1.8 ± 0.1 vs. 2.3 ± 0.1 fold relative to sham, P < 0.05). Internucleosomal DNA fragmentation of myocytes is hallmark of apoptotic cell death. No DNA ladder was found in sham and sham + PD 98059 group. However, DNA ladders were clearly detected in the area at risk zones in I/R, and I/R + Mag 5 group after 6 h of reperfusion. Treatments with magnolol (10 mg/kg) significantly decreased the appearance of DNA ladders due to I/R, which reappeared after PD 98059 treatment (Fig. 4). As shown in Figure 5, apoptotic death was evident in I/R heart by showing an increased number of TUNEL-positive cells (sham, 3% ± 1%; I/R, 38% ± 6%, P < 0.01). The increased number of TUNEL-positive cells by I/R injury was significantly reduced by magnolol 10 mg/kg (38 ± 6% vs. 18 ± 4%, P < 0.05), which was again reversed by PD 98059 (18 ± 4% vs. 31 ± 5%, P < 0.05).
Discussion
In the present study, we showed that administration of magnolol improved cardiac function and reduced the size of myocardial infarction following myocardial I/R in rats in vivo. We further found that magnolol modified apoptosis resulting from myocardial I/R suggesting that the beneficial hemodynamic effects and cardioprotective effects of magnolol given prior to myocardial ischemia are explained in part by its modulation of apoptosis-related gene expression. That magnolol modulates genes associated with apoptosis in the I/R-rat heart in vivo has not yet been reported. In addition, there are no in vivo studies to date that explore the relationship between the putative anti-apoptotic effects of magnolol on the functional recovery of I/R myocardium. The present study demonstrated that magnolol treatment preserved left ventricular function, as reflected by a significant increase in the indices of contractility (+ dP/dt) and relaxation (−dP/dt) and a decrease in preload (LVEDP). Impairment of the hemodynamic and ventricular functions are the most common fatal complications secondary to ischemic heart disease. We confirmed similar observations in I/R control rats as evidenced by the lower ±dP/dt and higher LVEDP. Magnolol, however, markedly reduced the loss of these hemodynamic parameters which improved not only heart function but also reduced the infarct size. The pronounced cardioprotection by magnolol is unlikely to be due to its hemodynamic effects because no significant changes in the MAP and HR occurred during experimental periods when compared with the vehicle group. We believe that magnolol inhibits apoptosis and that such an inhibition could be translated into a reduction of infarct size and an improvement of heart function.
Apoptosis has been linked with reperfusion-induced myocardial injury after reversible coronary occlusion (9–11). A potential role of apoptosis in the development of infarction also suggested in cardiac myocytes underwent apoptosis during reperfusion after 30 min coronary occlusion in rats (12). The mitochondrial death pathway appears to play an important role in the execution of apoptosis in cardiac myocytes (1, 13). Caspase-dependent cytochrome c release has been reported in isolated chick cardiac myocytes subjected to stimulated I/R (13), and several parameters of mitochondria-mediated apoptosis were demonstrated during myocardial I/R injury in vivo (14, 15) and ex vivo (16). Now, the question is how magnolol regulates genes associated with apoptosis. The exact answer is not known at the present time, but we speculate that antioxidant action may modulate Bcl-2 family by redox-sensitive manner. Indeed, magnolol possessed strong antioxidant activity as potent as 1000 times that of α-tochoperol in isolated rat heart mitochondria, where lipid peroxidation was induced with adenosine diphosphate (ADP) and ferrous sulfate (FeSO4) (17). Magnolol also exhibited free radical scavenging activity, but was less potent than α-tocopherol (17). In rat I/R liver, magnolol affected the formation of Bcl-xS and Bcl-xl (18), which reinforces our present results that magnolol indeed modulates genes related to apoptosis, especially Bcl-xl. However, it can not be ruled out that the possibility of metabolites of magnolol may have affected expression of these genes, because we observed this only in vivo study. Many antioxidants have been shown to inhibit apoptosis by regulating genes such as Bcl-2 family (Bcl-xl) (19). During I/R injury, the return of oxygen to ischemic tissues is accompanied by an increased production of reactive oxygen species (ROS). Studies on whole hearts and isolated cells have shown that a burst of ROS generation occurs during the first minutes after hypoxia, and ischemic tissues are reoxygenated (20) due to the reactivation of mitochondrial respiration (21). ROS generation results in tissue lipid and protein oxidation (22, 23). Therefore, antioxidants scavenging ROS blunt or eliminate I/R injury in vivo and in vitro (22, 24). It is well known that anti-apoptotic Bcl-2 protein has potent cardioprotective effects. Bcl-xl has been shown to protect against I/R injury ex vivo and in vivo (25–27). Gene transfer of Bcl-xl reduced cardiac cell apoptosis following cold preservation and warm reperfusion of rat cardiac transplants (26). Furthermore, elevated Bcl-xl protein levels coincided with the onset of protection during myocardial stunning (28).
The most striking finding is that anti-apoptosis of magnolol in rat I/R heart is mediated by activation of ERK1/2 signal pathway as evidenced by DNA strand break and TUNEL experiment, where magnolol showed anti-apoptotic effect but all of these effects of magnolol were reversed by PD 98059, an ERK 1/2 inhibitor. Consistent with this finding, upregulated expression of p-ERK, p-Bad, Bcl-xl and downregulated expression of cytochrome c by magnolol in rat I/R heart were also antagonized by PD 98059. These findings suggest that magnolol modulates the Bcl-2 family (Bcl-xl) protein expression via ERK phosphorylation and Bad phosphorylation. Although data was not shown, magnolol did not influence the other MAP kinases in the present study. In fact, ERK1/2 is an established player in the anti-apoptotic defense network. Recent studies have demonstrated that ERK may inhibit apoptotic signaling via post-transcriptional protein phosphorylation. Specifically, activation of ERK1/2 has been shown to result in Bad phosphorylation (Ser112) both directly (29–32) and indirectly via ERK/p90RSK activation (33, 34) which specifically suppresses Bad-mediated apoptosis.
In conclusion, we provide evidence that magnolol reduces myocardial damage and improves cardiac function by inhibiting rat I/R-induced apoptosis through ERK 1/2 activation. Thus, it can be useful for the treatment of I/R-induced injury, such as myocardial infarction.
Hemodynamic Data Under I/R Condition a
Hemodynamic Data Under Sham-Operated Condition a
Area at Risk and Infarct Size a

Myocardial infarct size in I/R and I/R + Mag 10 groups subjected to 30 min ischemia followed by 24 h reperfusion. Evans blue stained area is area not at risk, triphenyltetrazolium chloride (TTC) stained area (red staining) is ischemic but viable tissue, and TTC staining negative (white arrows) areas depict infarcted myocardium.

Effects of magnolol with or without PD 98059 treatment on p-ERK, p-Bad, Bcl-xl and cytochrome c proteins in reperfused rat heart after ischemia 30 min followed by 1 h or 6 h. (A) p-ERK expression in area at risk zone of cardiac tissue after 1 h reperfusion, p-Bad expression in area at risk zone of cardiac tissue after 6 h reperfusion. The phosphorylation was quantified by densitometry using image analysis program. (B) Bcl-xl and cytochrome c (cytosolic fraction) expression in area at risk zone of cardiac tissue after 6 h reperfusion. Blot shown is representative of four experiments with similar results. * P < 0.05 vs. I/R; †P < 0.01 vs. sham; ‡P < 0.05 vs. I/R + Mag 10; ‡‡P < 0.01 vs. I/R + Mag 10.

Effects of magnolol with or without PD 98059 treatment on caspase-3 activity in area at risk zone of cardiac tissues after 30 min of ischemia followed by 6 h reperfusion. Results are mean ± SEM from four separate experiments. * P < 0.05 vs. I/R; †P < 0.01 vs. sham; ‡P < 0.05 vs. I/R + Mag 10.
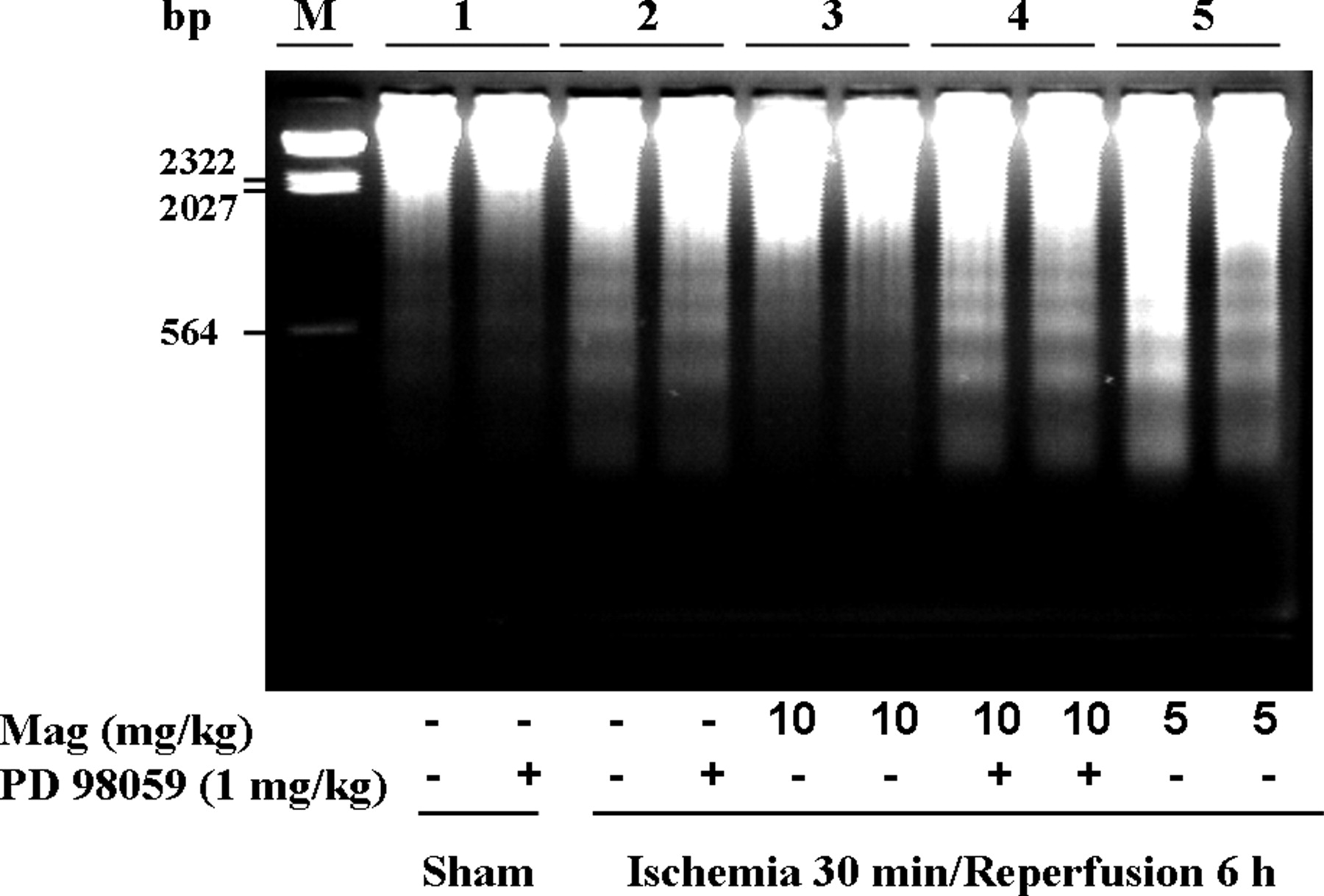
Detection of internucleosomal DNA fragmentation using agarose gel electrophoresis. M, marker lane; lane 1 represents sham tissue; lanes 2, 3, 4, and 5 represent area at risk zones tissue after 30 min ischemia followed by 6 h reperfusion in I/R control, I/R + Mag 10, I/R + Mag 10 + PD, and I/R + Mag 5 groups, respectively. Each lane represents a separate animal.

Detection of apoptotic myocytes using TUNEL technique in each group. Percentage of TUNEL-positive nuclei in rat hearts subjected to I/R injury with different treatment. Magnification × 100. Data are expressed as mean ± SEM from four separate experiments. * P < 0.05 vs. I/R; †P < 0.01 vs. sham; ‡P < 0.05 vs. I/R + Mag 10.
Footnotes
This work was supported by Korea Food and Drug Administration (S-06-02-2-CHM-230-0-C).