
Abstract
Although considered to be generally safe, a number of β-lactam antibiotics have been associated with epileptic seizures in humans. Furthermore, some β-lactam antibiotics, including ceftriaxone, are used to evoke convulsions under experimental conditions. Recently it was demonstrated that ceftriaxone increased expression of the glutamate transporter (GLT1) and its biochemical and functional activity in the brain of rodents. GLT1 regulates extracellular concentrations of glutamate, an excitatory amino acid involved in the pathogenesis of seizures and epilepsy. Because of its rapid transfer of glutamate into neurons and adjacent glial cells, GLT1 diminishes glutamate toxicity. We investigated whether ceftriaxone (200 mg/kg body wt) administered intraperitoneally (ip) for 6 days could modify the convulsant effects of pentylenetetrazole (PTZ, 100 mg/kg ip) in inbred male BALBcAnNCR and C57 black (BL)/6 mice aged 4 and 12 weeks. Ceftriaxone pretreatment provided significant protective effects against PTZ-evoked generalized clonic convulsions (GCCs), generalized clonic-tonic convulsions (GCTCs), and convulsion-induced mortality during a period of 30 mins after PTZ administration. The incidence of GCCs, GCTCs, and death was statistically significantly lower for BALBcAnNCR mice of both ages, particularly younger mice. The latency time for each of the three parameters was significantly greater, with the exception of GCCs in adult mice. Protective effects of ceftriaxone were also noticed in adult C57BL/6 mice but not in prepubertal C57BL/6 mice. This is the first demonstration of anticonvulsant effects of ceftriaxone or any other β-lactam antibiotic, which are not uniform across the mouse population. Our results provide new insight into the effects of ceftriaxone, which need further investigation.
Introduction
Epilepsy is a huge medical and economic problem around the world. Successful control of epilepsy is hindered by its complex pathogenesis. To improve our understanding of neurochemical processes in the brain, chemicals and other convulsant sources are used in experimental models of epilepsy.
Epileptic seizures are associated with a number of prescribed drugs. Therefore, many prescribed drugs are not recommended, especially for epilepsy-prone patients. Although the β-lactam family of antibiotics are mostly safe in the central nervous system, epileptic activity has been reported as a consequence of their use in human subjects (1). The exact incidence of this complication is not known. However, it is believed to be low if therapeutic antibacterial doses are prescribed.
Convulsive effects were first associated with penicillin, the first β-lactam antibiotic that was discovered. Some new generations of this class of antibiotics are not totally free of convulsant effects, although some exceptions have been noted (2). β-Lactam antibiotics can evoke not only convulsive reactions but also nonconvulsive reactions; in particular, nonconvulsive status epilepticus in humans associated with cephalosporin use is probably underestimated (3). Under certain conditions, β-lactam antibiotics are able to evoke different patterns of convulsions in diverse animal species (4, 5). Therefore, they are useful in the study of epilepsy under experimental conditions.
Epileptic seizures associated with β-lactam antibiotics have been ascribed to the inhibition of the γ-aminobutyric acid (GABA) signaling system, resulting in inhibition of postsynaptic GABA ion channels and a subsequent a decrease in GABA-mediated inhibitory transmission (6, 7). Dose-dependent convulsive activity of cephalosporins has been noted (7). These drugs compete only with GABA at the GABAA receptor site and do not have any affinity for the N-methyl-
To a certain degree, epileptic convulsions evoked by β-lactams are related to the chemical structure of these drugs. De Sarro et al. discovered a resemblance between the chemical structure of β-lactam antibiotics and that of GABA receptor antagonists such as bicuculline and the chemical convulsant pentylenetetrazole (PTZ) (8).
In addition to the discovery that GABAergic antagonistic effects are capable of triggering epileptic seizures, recent research has demonstrated that β-lactam antibiotics are able to modulate glutamate activity in the brain. By testing 1040 drugs and nutritionals approved by the U.S. Food and Drug Administration (FDA), Rothstein and coworkers found that some β-lactam antibiotics, including ceftriaxone, administered during 6 consecutive days at doses routinely used to treat central nervous system infections “increased both the expression of GLT1 (glutamate transporter) and its biochemical and functional activity in vitro and in vivo in the brains of rats and mice” (9).
Excitatory amino acids (EAA), as well as GABA, are involved in the pathogenesis of seizures and epilepsy. Under physiologic conditions, glutamate is continuously released from nerve terminals into the synaptic cleft. Because glutamate does not appear to be significantly metabolized outside cells, transporters are the most important mechanism for removing it from the extracellular space. Transporters of EAA are located presynaptically, postsynaptically, and extrasynaptically, in cell plasma membranes, and within the cells. In addition, transporters play a key role in regulating extracellular concentrations of EAA, particularly in the synaptic cleft. The rapid transfer of EAA into neurons and adjacent glial cells diminishes the opportunity for EAA-mediated toxicity.
Glutamate is primarily transported by GLT1, which is particularly abundant in astrocytes, where glutamate is converted to glutamine. For this reason, astrocytes are the main regulators of the extracellular concentration of neurotransmitters such as glutamate and GABA (glutamate-glutamine cycle).
Glutamate transporters are involved in physiologic and pathologic signaling in the brain. Their dysfunction has been found in some animal models of neurologic disorders, including epilepsy (10). Tanaka et al. demonstrated a greater susceptibility to seizures in mice lacking GLT1 (11). In GLAST (glutamate-aspartate transporters) knockout and wild-type C57 black mice, Watanabe and colleagues noted similar occurrences (12). Furthermore, Milh et al. concluded the following: “our data suggest that a deficient transport of glutamate (in the cortex of immature rats) may be a potential cause for early onset of epilepsy” (13).
An opposite result can be expected in the case of increased GLT1 activity. For example, in an experimental mouse model of amyotrophic lateral sclerosis, Rothstein et al. found protective effects of ceftriaxone treatment (9).
When all of the previously reported observations are taken into account, it is clear that β-lactam antibiotics can influence the functions of the two main neurotransmitters involved in seizures: β-lactam antibiotics decrease the inhibitory effects of GABA and diminish glutamate transmission. The epileptic effects of β-lactam antibiotics mediated through the GABAergic system have already been thoroughly examined. However, it is not known whether β-lactam antibiotics are able to prevent seizures induced by convulsant chemicals. Therefore, the aim of our study was to investigate whether a therapeutic dose of ceftriaxone administered during 6 successive days could influence generalized motor convulsive responses to a single PTZ treatment in two strains of mice not susceptible to seizures.
Materials and Methods
Animals.
The experiments were performed on inbred male mice belonging to BALBcAnNCR (10 to 16 in each experimental group) and C56 BL/6 strains (8 to 10 in each experimental group). Mice exactly 4 weeks old and 12 weeks old at the beginning of the experimental treatments were used. All mice had free access to standard food and tap water and were kept under humane and environmentally controlled conditions at a room temperature of 23° ± 2°C and a 13:11-hr light:dark cycle. All mouse handling took place between 0900 hrs and 1300 hrs. The mice were weighed daily because the doses were based on body weight. Five mice were routinely housed in one cage. Each mouse was used only once. The whole research protocol strictly followed the rules laid out in the “Guidelines for Animal Study No 282–12/2002” (proposed by the Ethical Committee of the Military Medical Academy, Belgrade, Serbia).
Treatments.
Control mice (mice of both strains and of both ages) received 0.9% NaCl intraperitoneally (ip) once daily during 6 consecutive days. Ceftriaxone (Azaren, Panfarma, Vrsac, Serbia) was dissolved in sterile distilled water and was given in the same manner to the other groups. The daily dose of ceftriaxone was 200 mg/kg body wt. The protocol of ceftriaxone administration was the same as that used by Rothstein and coworkers (9).
On the seventh day both control mice and ceftriaxone-treated mice were randomly selected for intraperitoneal administration of a single dose of PTZ (100 mg/kg body wt; Sigma, St. Louis, MO) dissolved in 0.9% NaCl.
Observation of Convulsive Responses.
After a PTZ injection, each mouse was placed into a transparent cage for behavioral monitoring lasting 30 mins. Clinical responses to the administered substances were classified as follows (in successive order of their appearance): generalized clonic convulsions (GCCs, i.e., clonic seizures that involved the entire body with loss of righting ability), generalized clonic-tonic convulsions (GCTCs, i.e., the appearance of a tonic response of the hind limb extensor muscle [hind limb extension followed GCC]), and mortality as a consequence of the convulsions.
We recorded the time interval between the PTZ injection and the onset of any of the two convulsion patterns and death (latency time), and we also determined the incidence of convulsion patterns and death. A time of 1800 secs was recorded if convulsions and/or death did not occur during the 30-min monitoring period.
Statistics.
The latency times of GCCs, GCTCs, and death were calculated in seconds and expressed as the mean ± SD. The incidences of the convulsion patterns and death were expressed as percentages. The former results were analyzed by the Mann-Whitney U test, whereas the latter were analyzed by the Fisher exact probability test. Differences between experimental groups were considered significant when P < 0.05.
Results
Effects of PTZ on Convulsions and Death.
When PTZ was administered after 6 days of 0.9% NaCl pretreatment, we observed GCCs, GCTCs, and death in both strains of mice. The incidences of GCCs, GCTCs, and death were similar in 4-week-old and 12-week-old mice of both strains (Fig. 1A and B).
For 4-week-old BALBcAnNCR mice, the latency time of PTZ-evoked GCCs was greater than that for 12-week-old mice (P < 0.01; Table 1). The latency time of PTZ-evoked GCTCs and death for 4-week-old C57BL/6 mice was greater than that for 12-week-old mice of the same strain (P < 0.05 for both parameters).
For 4-week-old mice of the B57BL/6 strain, the latency times of PTZ-evoked GCTCs and death were greater than those for the BALBcAnNCR mice (P < 0.05; Table 1). In addition, the latency time of PTZ-evoked GCCs was greater in 12-week-old C57BL/6 mice than in BALBcAnNCR mice (P < 0.05).
Effects of Ceftriaxone on the Incidence of PTZ-Evoked Convulsion Patterns and Death.
Comparison Between 4-Week-Old and 12-Week-Old BALB-cAnNCR Mice.
Compared with the NaCl + PTZ–treated group, the group of 4-week-old mice pretreated with ceftriaxone had lower incidences of PTZ-evoked GCCs, GCTCs, and death (P < 0.01 for all parameters), whereas the group of 12-week-old mice had only a significantly decreased incidence of death (P < 0.05; Fig. 1A).
Ceftriaxone’s protection against GCCs and GCTCs was more pronounced in 4-week-old mice than in 12-week-old mice (P < 0.01 and P < 0.05, respectively).
Comparison Between 4-Week-Old and 12-Week-Old C57BL/6 Mice.
Ceftriaxone decreased the incidences of GCCs, GCTCs, and death only in 12-week-old mice when they were compared with those of the NaCl + PTZ–treated group (P < 0.05 for all three parameters; Fig. 1B).
In the ceftriaxone pretreated groups, the incidence of GCCs differed according to the age of the mice: the 12-week-old mice were more susceptible to ceftriaxone’s protective effects than were the 4-week-old mice (P < 0.01).
Comparison Between BALBcAnNCR and C57BL/6 Mouse Strains.
The protective effects of ceftriaxone were more prominent in 4-week-old BALBcAnNCR mice than in C57BL/6 mice (Fig. 1A and B). Lower incidences of GCCs (P < 0.01) and death (P < 0.05) were recorded for the BALBcAnNCR strain. Opposite effects in 12-week-old mice were recorded: ceftriaxone-mediated protection against GCCs and GCTCs in C57BL/6 mice was greater than that in BALBcAnNCR mice (P < 0.01 and P < 0.05, respectively).
Effects of Ceftriaxone on the Latency Times of PTZ-Evoked Convulsion Patterns and Death.
Comparison Between 4-Week-Old and 12-Week-Old BALBcAnNCR Mice.
The latency times of GCCs, GCTCs, and death were significantly more prolonged in the ceftriaxone pretreated mice of both ages than in the control (NaCl +PTZ) experimental groups (P < 0.01 for all but one parameter; Table 1). The exception was the latency time for GCCs in 12-week-old mice; this time was also prolonged, but the difference exhibited borderline significance (P = 0.053).
The protective effects of ceftriaxone, which were indicated by the latency times of GCCs, GCTCs, and death, were more notable in 4-week-old BALBcAnNCR mice than in 12-week-old mice (P < 0.01, P < 0.01, P < 0.05, respectively).
Comparison Between 4-Week-Old and 12-Week-Old C57BL/6 Mice.
The latency times of GCCs, GCTCs, and death were increased by ceftriaxone pretreatment in 12-week-old mice compared to NaCl +PTZ treated mice of the same age (P = 0.003, P = 0.03, and P = 0.03, respectively; Table 1). Four-week-old mice were not sensitive to the protective effects of ceftriaxone.
The latency time of PTZ-evoked GCCs differed according to age. The latency time was greater in 12-week-old mice pretreated with ceftriaxone than in 4-week-old mice pretreated with the same drug (P < 0.01).
Comparison Between BALBcAnNCR and C57BL/6 Mice.
The latency time of GCCs in 4-week-old BALB-cAnNCR mice was greater than that in C57BL/6 mice of the same age (P < 0.01; Table 1). The opposite finding was noted when the latency time of GCCs of 12-week-old C57BL/6 mice was compared with that of the BALB-cAnNCR mice of the same age (P < 0.01).
Discussion
Anticonvulsant effects of β-lactams, including the anticonvulsant effects of therapeutic doses of ceftriaxone, are not expected and have not been previously investigated. To the best of our knowledge, our research is the first demonstration of the anticonvulsant effects of ceftriaxone or of any β-lactam antibiotic.
We found that therapeutic doses of ceftriaxone administered over 6 consecutive days protected some BALBcAnNCR and C57BL/6 mice against PTZ-evoked convulsions and death. The protection resulted in a decreased incidence of convulsion patterns and convulsion-induced mortality and in an extension of the latency time of both convulsions and death (Fig. 1 and Table 1).
Ceftriaxone-pretreated mice did not homogeneously react to PTZ. This finding partially agrees with those of studies that demonstrated nonuniform responses by immature and mature rats, with particular regard to the latency time for convulsive reactions toward PTZ (14). Furthermore, De Sarro et al. registered different susceptibility to audiogenic seizures among mouse strains (15). Kosobud et al. found that strains of mice differed in their susceptibility to PTZ-evoked convulsions. Under some circumstances, C57BL/6J mice were more resistant to PTZ-evoked convulsion patterns than were BALB/cJ mice (16). Such differences in susceptibility reflect age and strain heterogeneity of neurochemicals in brain regions involved in convulsive processes (17, 18).
The exact mechanism of PTZ action is not well understood. It is believed that its main action is mediated by its binding to the postsynaptic GABAA receptors (19). By competitive inhibition, PTZ antagonizes the GABA-activated current and reduces the frequency of chloride channel opening in a concentration-dependent manner. Thus, PTZ lowers the depolarization threshold.
Beside GABA involvement, the data about involvement of glutamate in PTZ action are also in accordance. Furthermore, Lukomskaya and coworkers suggested that glutamatergic synaptic transmission plays an important role in the weakening of GABAergic inhibitory processes induced by PTZ (20). Currently, PTZ is a useful substance for epilepsy research, for screening new compounds with potential antiepileptic activity, and for use in studies regarding anxiety.
Ceftiaxone belongs to the third generation of cephalosporins. Its pharmacokinetics is somewhat specific to the brain. It does not penetrate into the cerebrospinal fluid (CSF) as much as penicillin G; the large molecular size of ceftriaxone plays a part in limiting its ability to cross the blood-brain barrier. However, ceftriaxone attains its effective bactericidal concentrations in the CSF because it has less affinity for the transport system in the chorioid plexus than does penicillin and it is slowly transported from the CSF to blood (21). Therapeutic doses of ceftriaxone are usually administered once daily, except in some severe diseases for which it can be administered twice daily.
Of the three types of GABA receptors (GABAA, GABAB, and GABAC), the GABAA receptor is the one whose inhibitory effects are influenced by ceftriaxone and PTZ. According to the results of Sugimoto et al. and other research groups that have documented the convulsant activity of β-lactam antibiotics, PTZ given to animals pretreated with ceftriaxone should produce more pronounced convulsive effects (22). However, we found the exact opposite effects of ceftriaxone: ceftriaxone provided protection against convulsive responses and death induced by PTZ (Fig. 1 and Table 1).
There are several reasons for such a disagreement. For instance, predisposition and the presence of risk factors in patients (and in animals) contribute to the epileptic activity of β-lactam antibiotics. These factors include accompanying diseases (especially those of the brain and the kidney) and concurrent use of other drugs. Such factors can increase the brain’s susceptibility to seizures and/or change the pharmacokinetics and the pharmacodynamics of β-lactam antibiotics (23, 24). Such a predisposition and the presence of risk factors for convulsant activity induced by ceftriaxone were eliminated in our experiments, as our research was performed with healthy mice.
The discrepancy between our results and those present in the literature might also be due to the administered dose of β-lactam antibiotic. In studies of β-lactam antibiotic–induced epileptic seizures in humans and animals, the administered doses were (intentionally or accidentally) higher than those used for therapeutic purposes. For example, severe and treatment-resistant epileptic seizures appeared in a patient after the unintentional administration of a high dose of mezlocillin in a brain ventricle (1). In another report Marangoz and Bagirici applied 3 × 106 units of penicillin G per kilogram of body weight to provoke epileptic activity in rats, whereas Jimenez et al. stereotaxically administered 250 to 500 units of penicillin into multiple brain structures and registered motor and electroencephalographic seizures in awakened cats (25, 26). Jin et al. lowered the PTZ convulsive threshold in rodents by intravenously administering 800 mg/kg body wt cefazolin but not by intravenously administering 400 mg/kg (27). In accordance with Sugimoto et al., De Sarro et al. recorded dose-dependent convulsant effects of some β-lactam antibiotics, including ceftriaxone, given intracerebroventricularlly (7, 8). Also, Williams et al. registered a decreased PTZ convulsive threshold by a single intravenous application of subtoxic doses of β-lactam antibiotics (5).
Unlike the experiments described above, the daily dose of ceftriaxone used in our experiments was within the antibacterial therapeutic range. Furthermore, we injected ceftriaxone during 6 consecutive days, whereas the previously mentioned researchers applied a single dose of this drug. According to Rothstein and associates, a period of 6 days is sufficient to increase GLT1 expression and activity, and these effects of ceftriaxone appear early after the beginning of the treatment (within 2 days) and persist for a considerable time (up to 3 months; Ref. 9). Because glutamate is involved in the pathogenesis of PTZ-evoked convulsions, ceftriaxone administration could therefore modify the effects of PTZ through modulation of the GLT1 transporter and glutamatergic activity.
Our observations regarding age-dependent and strain-dependent susceptibility to ceftriaxone’s protective effects could have been related to the activity of ceftriaxone on GLT1. It has been documented that glutamate transporters exhibit age-dependent morphologic and biochemical characteristics in the brain (9, 13, 28). Also, there are differences between glial and neuronal architecture in the brain during development and maturation, including developmental changes in the levels of GLT protein and the susceptibility to endogenous excitatory and inhibitory neurotransmitters (29). Furthermore, the distribution of GLT1 in the brain is not uniform: it is region-dependent and species-dependent (30, 31). All of the above mentioned findings could contribute to the nonhomogeneous effects of ceftriaxone demonstrated in our research, which is supported by Lipski et al.: “the beneficial effects of treatment with β-lactams do not appear to be universal and may confined to specific developmental stages/ages and the type of stressor (e.g., hypoxia, excitotoxicity) and experimental model” (32).
Contrary to a number of agonists and antagonists for different types of glutamatergic receptors, no specific drugs capable of modulating the activity of glutamate transporters are known. Therefore, substances that can selectively modulate GLT1 may be of interest for developing antiepileptic and neuroprotective drugs, which would be important for clinical practice and for pharmacotherapy.
In summary, by administering ceftriaxone we demonstrated a protective effect against PTZ-evoked convulsions and death (the latter as a consequence of convulsions). The protective effects were registered in prepubertal adolescent and adult BALBcAnNCR mice as well as in adult C57BL/6 mice. Our results suggest the involvement of the glutamatergic system in the action of PTZ. Further studies focusing on the anticonvulsant effects of ceftriaxone are required in other animal species and in different experimental models of convulsions to gain a broader knowledge of the drug’s anticonvulsant effects.
The Influence of Ceftriaxone on the Latency Time for PTZ-Evoked Convulsions and Death in 10 to 16 BALBcAnNCR Mice and 8 to 10 C57BL/6 Mice a
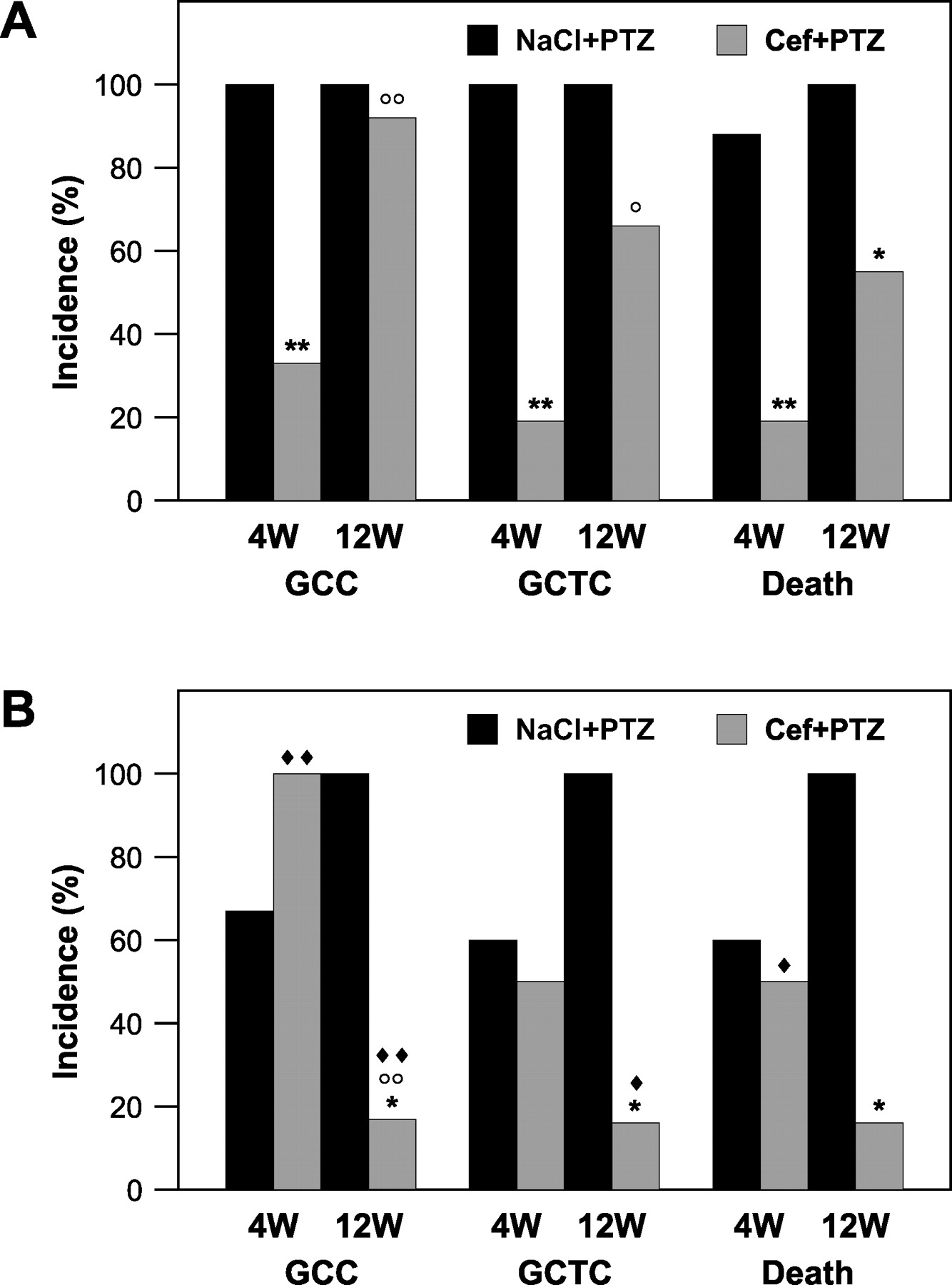
(A) The influence of ceftriaxone on the incidence of PTZ-evoked convulsions and death of BALBcAnNCR mice (10–16 mice). (B) The influence of ceftriaxone on the incidence of PTZ-evoked convulsions and death of C57BL/6 mice (8–10 mice). Ceftriaxone (Cef), 0.9% NaCl, and PTZ were applied ip; NaCl and Cef were administered over 6 consecutive days (Cef in a single daily dose of 200 mg/kg body wt), and PTZ was given as a single dose of 100 mg/kg body wt. The abbreviation “W” refers to weeks of age at the beginning of experiments. The Fisher exact probability test was used to test differences in the incidences of seizure and death.
Footnotes
The Ministry of Science of the Republic of Serbia (contract 143057) supported this research.