
Abstract
The discovery of drugs that can be used for the treatment of allergic disease is important in human health. Arctium lappa Linne (Compositae) (AL) has been used as a traditional medicine in Brazil and throughout Asia and is known to have an anti-inflammatory effect. In this study, the inhibitory effects of AL on degranulation and the release of mediators as well as on inhibition of cys-leukotriene biosynthesis by basophils were investigated. AL was selected out of 10,000 herbal extracts in a set-up for high throughput screening in which the degree of degranulation was monitored by the release of β-hexosaminidase from rat basophil leukemia (RBL-2H3) cells. The AL extract significantly reduced degranulation and biosynthesis of cys-leukotrienes of human basophils in peripheral blood mono-nuclear cells (PBMCs) (50% inhibitory concentration [IC50] = 8.3 and 11.4 μg/ml, respectively). Viability and metabolic activity of the PBMCs were not affected. Although arctiin, the active component of AL that has been described in the literature, was not able to reduce degranulation in RBL-2H3 cells, a single high-performance liquid chromatography (HPLC) fraction from the AL extract inhibited β-hexosaminidase release (IC50 = 22.2 μg/ml). Topical administration of an aqueous extract of AL (5 mg/ear) on the ear of whey-sensitized mice 4 hrs before challenge with whey in the ear inhibited acute ear swelling by 50% in an in vivo cow’s milk allergic model. The extract had no effect in this model when administered orally. In conclusion, the active component present in the active HPLC fraction of the AL extract was able to significantly reduce the release of inflammatory mediators through inhibition of degranulation and cys-leukotriene release in vitro. In addition, this active component was able to inhibit acute skin response in mice in vivo, indicating that AL is a very promising natural component for use in anti-allergic treatment.
Introduction
The prevalence of allergic diseases in Western countries has risen substantially over the last few decades (1). Allergy may be defined as the potential for development of immunologically mediated reactions to allergens which, in 80% of the allergy-based clinical diseases in man, is mediated by IgE antibodies (2). Exposure to an antigen sets off an immune-mediated cascade of inflammatory events. First, the allergen is broken down into smaller peptides and exposed to T cells by antigen-presenting cells. The T cells produce cytokines such as interleukin 4 (IL-4) that induce B cells to produce antigen-specific IgE, which binds to high-affinity FcεRI receptors on basophils and/or mast cells (3). On a second encounter with the same allergen, the allergen cross-links the IgE bound to FcεRI receptors, activating them and causing the release of inflammatory mediators such as histamine, prostaglandins, and leukotrienes (4). Preformed mediators, such as histamine, neutral proteases and other enzymes, and chemotactic factors, are present in granules and released by fusion of the granule membranes with the cell membrane. Other substances, such as cytokines, and lipid mediators, such as prostaglandins and cysteinyl leukotrienes (LTC4, LTD4, and LTE4) are newly synthesized and secreted following cross-linking. Current treatment of allergic symptoms consists of antihistamines (5), leukotriene receptor antagonists (6, 7, 8, 9), mast-cell stabilizers (10, 11), and corticosteroids (12). Some of these treatments are known to have severe adverse effects (5, 13, 14, 15).
Arctium lappa Linne (AL; burdock) is a biennial member of the Asteraceae (Compositae) family, and its carrot-like root is commonly cooked and eaten as a vegetable in parts of Asia. In traditional Brazilian and Asian folk medicine, AL is used for the treatment of almost all infectious diseases. Several studies have been performed to screen plants for medicinal activity. Holetz et al. (16) studied 42 plants with regard to their in vitro antimicrobial activity. AL leaves showed moderate activity against gram-positive and gram-negative bacteria. Lin et al. (17) demonstrated that the crude root extract of AL has anti-inflammatory effects by inhibiting carrageenan-induced rat paw edema, which might be due to its free radical–scavenging activity. Iwakami et al. (18) found that a hot aqueous extract of AL had an inhibitory effect on the binding of platelet-activating factor, a compound released from activated basophils, causing platelet aggregation, which is relevant for inflammatory processes such as those seen in allergy. Biologically active components isolated from parts of the AL plant are arctiin and its metabolite arctigenin (19, 20, 21, 22, 23, 24), pectin (25), fructooli-gosaccharides (26), inulin (27), and fructofuranan (28) from the root.
The present study examined the pharmacologic activity of the AL extract as an anti-allergic agent, using both in vitro and in vivo models. In summary, the data indicate that AL has anti-inflammatory properties and might be useful in the treatment of allergic inflammation.
Materials and Methods
Cell Culture.
The rat basophil leukemia cells RBL-2H3 (ATCC CRL-2256) and RBL cells transfected with the α-chain of the human Fcε receptor type 1 complex (RBL-hEIa-2B12 cells) (29) were cultured in Dulbecco’s Eagle MEM (Life Technologies, Merelbeke, Belgium), supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Greiner, Alphen a/d Rijn, The Netherlands) and 100 IU/ml penicillin/streptomycin (Life Technologies).
For RBL-hEIa-2B12 cells, another 50 mg/ml of geneticin (Life Technologies) was added. Cells were grown in a 75-cm2 culture flask and passed twice a week using trypsin-EDTA (Life Technologies).
Peripheral blood mononuclear cells (PBMCs) were freshly isolated by Ficoll-Paque (Pharmacia Biotech, Diegem, Belgium) density-gradient centrifugation from buffy coats of healthy donors (Sanquin, Amsterdam, The Netherlands). The PBMCs were cultured overnight in RPMI-1640 (Life Technologies) with 2 mM
Herbal Extracts and Controls.
Complete AL plants, including roots and leaves but without seeds, were harvested and extracted by PhytoMyco Research Corporation (South Carolina).
For high throughput screening, all herbal extracts were tested in a final concentration of 100 μg/ml EtOH 20%. Extracts that reduced the release of β-hexosaminidase by > 55% were studied for their dose-response effects in concentrations of: 200, 100, 50, 25, 12.5, 6.3, 3.1, and 1.6 μg/ml. The flavonoid quercetin (Sigma-Aldrich Chemie) was used as a positive control for inhibition of β-hexosaminidase release in RBL cells in a concentration of 50 μM (30). In the RBL degranulation assay with cells containing the human receptor, AL extract concentrations of: 320, 160, 80, 40, 20, and 10 μg/ml were used. For human degranulation studies, AL extract concentrations of: 0.5, 1, 2.5, 5, 7.5, and 10 μg/ml were used. Nordihydroguaiaretic acid (Fluka, Zwijndrecht, The Netherlands), a well-known 5-lipoxygenase inhibitor, was used as a positive control for inhibition of the production of cys-leukotrienes by PBMCs in concentrations of: 0.35, 0.7, and 1.4 μg/ml (31).
Whey-sensitized mice were orally treated three times with 5 mg AL in 0.5 ml phosphate-buffered saline (PBS). For topical administration, a vehicle cream was used for application of AL to the ear. Three concentrations of AL (0.5, 1.0, and 5.0 mg/ear) were either directly added to the cream or first dissolved in water before adding to the cream.
Degranulation Assay for High Throughput Screening (RBL-2H3 Cells).
Confluent growing RBL-2H3 cells (1 × 105/well) in a 96-well, flat-bottom, white culture plate were sensitized by 100 ng/ml mouse IgE α-dinitrophenyl (DNP; Sigma-Aldrich Chemie) in a 100-μl culture medium. After 18 hrs of culture at 37°C/5% CO2, the cells were washed with 100 μl Tyrode’s buffer (130 mM NaCl [Merck], 190 mM KCl [Merck], 1.4 mM CaCl2 [Sigma-Aldrich Chemie], 1 mM MgCl2 [Sigma-Aldrich Chemie], 5.6 mM glucose [Merck], 10 mM HEPES [Merck], and 0.1% bovine serum albumin [BSA; ICN, Zoetermeer, The Netherlands] in distilled water, pH 7.4).
Twenty-five microliters of the herbal extracts (stock: 6 mg/ml in ethanol 20%) and 25 μl of DNP-BSA 100 ng/ml (Molecular Probes, Breda, The Netherlands) were supplemented to the adhered cells, according to Hoffmann et al. (32).
Maximum release was measured using 10 μl 1% v/v Triton X-100 (Sigma-Aldrich Chemie) in Tyrode’s buffer. Cells were incubated for 1 hr at 37°C/5% CO2. Fifty microliters of 4-methyl umbelliferyl-N-acetyl-β-
Degranulation Assay with RBL Cells with Human Receptor (RBL-hEIa-2B12 Cells).
Confluent-growing RBL-hEIa-2B12 cells (1 × 105/well) in a 96-well, flat-bottom culture plate were sensitized with human IgE purified (Chemicon) (5 μg/ml) for maximum release with IgE or specific IgE from cow’s milk–allergic (CMA) (β-lactoglobulin–specific IgE 74.6 Ku/l) or peanut-allergic (peanut-specific IgE < 50 Ku/l) patients, diluted 1:50 in 100 μl culture medium. After 18 hrs of culture at 37°C/5% CO2, cells were washed with 100 μl Tyrode’s buffer and incubated with α-hIgE (BD Pharmingen, Franklin Lakes, NJ) (5 μg/ml) for maximum release and AL extract in a serial dilution (320 μg/ml to 10 μg/ml), together with either 500 ng/ml milk powder (Numico Research BV, Wageningen, The Netherlands) or 50 ng/ml peanut allergen (TNO, Zeist, The Netherlands) to induce cross-linking. Measurement of β-hexosaminidase release was performed as described above.
Degranulation Assay with Human PBMCs.
After coculture of PBMCs with IL-3 and the AL extract for 18 hrs, 5 × 104 PBMCs/ml were incubated for 4 hrs with tetrazolium salts (WST-1, Boehringer Mannheim, Mannheim, Germany) to determine metabolic activity. PBMCs (3 × 106/ml) were stimulated with 50 μl α-IgE receptor antibody (CAST-ELISA, DPC) in 200 μl stimulation buffer (CAST-ELISA; DPC, Breda, The Netherlands) for 40 mins to induce cross-linking.
The biosynthesis of cys-leukotrienes (LTC4, LTD4, and LTE4) was analyzed in the supernatant of the stimulated cells by the use of CAST-ELISA (DPC) (34). α-CD203c-PE and α-CD63-FITC (Coulter Immunotech, Fullerton, CA) were diluted 1:200 in PBS supplemented with 2% FBS and incubated for 30 mins in the dark on ice. These markers were used to determine basophils and degranulation, respectively, by flow cytometry on the Coulter Epics XL (Beckman Coulter, Fullerton, CA) (35). The fluorescent DNA stain 7-amino-actinomycin D (7-AAD) (Coulter Immunotech) can penetrate the membranes of dying or dead cells and was used for identification of necrotic/late apoptotic cells (36). Degranulation of human basophils was defined as 7-AAD−/CD203c+ /CD63+ cells.
Oral Sensitization and Challenge of Mice with Whey.
Three- to five-week-old specific pathogen-free female C3H/HeOuJ mice (n = 4–6 per group) were purchased from Charles River Laboratories (Maastricht, The Netherlands), maintained on cow’s milk protein–free mouse chow (Special Diets Services, Witham, Essex, UK), and housed in the animal facility at University Medical Center, Utrecht. Animal care and use were performed in accordance with the guidelines of the Dutch Committee of Animal Experiments. As previously described (37), the mice were sensitized using intragastric administration at days 0, 7, 14, 21, and 28 with 0.5 ml homogenized whey (40 mg/ml PBS) (DMV International, Veghel, The Netherlands), with cholera toxin (CT, 20 μg/ml PBS) (Quadratech Diagnostics, Surrey, UK) as an adjuvant, using a blunt needle. Control mice received CT. Booster doses were administered weekly for 6 wks; 1 wk after the last sensitization, the mice were challenged, using intragastric administration, with 100 mg whey in 1 ml PBS. Mice that received oral treatment with AL were administered 1 dose of AL 5 mg in 0.5 ml PBS on days 27, 30, and 33.
Allergen-Specific Skin Response.
The acute allergen-specific skin response was measured after intra-dermal injection of the specific protein into the ear pinnae. In topical administration, AL was applied to the ears 4 hrs before challenge. Before intragastric challenge (t = 0), the control and whey-sensitized mice were injected intradermally in the left ear with 20 μl homogenized whey (0.5 mg/ml in PBS). The CT mice also received a whey ear challenge as control. Ear thickness was measured in duplicate, using a digital micrometer (Mitutoyo, Veenendaal, The Netherlands), at t = 0 and 1 hr after challenge. The allergen-specific net ear swelling was calculated by subtracting the basal thickness (0 h) from the thickness measured at 1 hr after injection. In addition, the control swelling measured at the same time points was subtracted. The ear swelling is expressed as delta μm.
Activity-Guided Fractionation of AL.
Analytical high-performance liquid chromatography (HPLC) (Akta Purifier, Pharmacia) was performed to analyze the chemical fingerprint of the AL extract according to the AGI method, which is an in-house method developed by Numico Research. A Superspher 100 RP18 4 μm 125 × 3 mm column, with precolumn 10 × 3 mm (Bischoff, Leonberg, Germany), bedvolume analytical column 0.9 ml, was employed using two solvent systems: 0.04% (v/v) trifluoro-acetic acid (TFA) in demiwater (A) and 0.04% (v/v) TFA in acetonitrile (B). The elution gradient profile was set as follows: 2 CV 100% A, in 20 CV to 25% B, in 5 CV to 50% B, in 5 CV to 100% B, 2 CV 100% B, while flow rate was set at 0.45 ml/min and the detector wavelength was set to 220 and 254 nm.
Analysis.
For all parameters, the positive control in which all conditions were comparable with the supplemented samples, except for supplementation, was set at 100%. Samples were related to the control samples. IC50 values were determined by calculating a statistically validated nonlinear regression curve through the plotted acquired values. The x values (i.e., dosage) corresponding to the maximal y value divided by 2 were calculated from the mathematical regression curve formula. Ear swelling was expressed as mean ± SEM. Analyses were performed using the statistical software package GraphPad Prism, version 4.03. For comparison with the control group, values of P < 0.05 were considered significant.
Results
High Throughput Screening Degranulation (RBL-2H3) Cells.
A high throughput study of approximately 10,000 herbal extracts dissolved in EtOH 20% was performed to investigate their inhibitory effects on the release of β-hexosaminidase from RBL-2H3 cells after FcεRI cross-linking by α-DNP-IgE and DNP-BSA. During the set-up of the assay it was determined that EtOH 20% has only a very slight inhibiting effect on degranulation (data not shown); this effect was corrected by use of the same amount of EtOH in the controls. Ninety-nine extracts were able to reduce degranulation by > 55% (Fig. 1), which was confirmed as positive in a dose-response study (data not shown). Among these 99 extracts, 10 extracts appeared to be applicable for use in nutrition. The most prominent inhibitor of degranulation, as indicated by the release of β-hexosaminidase, was the food grade extract of AL.
Different AL Extracts.
Of the eight different AL extracts (extracts A–H) among the 10,000 screened herbal extracts present in the herbal library, only one (extract H) exhibited inhibition of β-hexosaminidase release. These eight AL extracts were studied in a dose-response fashion (Fig. 2). As EtOH 20% solvent previously showed minimal effect on the release of β-hexosaminidase, the extracts were dissolved in water and then compared to EtOH 20%. Extract H showed a strong inhibitory effect when dissolved in water (IC50 = 12.6 μg/ml), which was comparable to the inhibition of the extract dissolved in EtOH 20% (IC50 = 12.4 μg/ml, data not shown). However, water alone showed no effect on β-hexosaminidase release. None of the other extracts from different suppliers was found to decrease β-hexosaminidase release. This finding suggests that the extraction method used by PhytoMyco Research Corporation maintained the anti-allergic activity of AL.
Degranulation of RBL Cells Containing Human IgE Receptor (RBL-hEIa-2B12 Cells).
RBL cells transfected with the α-chain of human Fcεreceptor have the important property of binding human IgE from the sera of allergic individuals; the cells can be activated in an allergen-specific manner. Sera used in this assay were obtained from individuals suspected to have food allergy and were assessed using double-blind, placebo-controlled food challenge tests (38, 39). The RBL cells were incubated with serum from either CMA individuals or peanut-allergic individuals. After this incubation step, the sensitized RBL cells were incubated with milk powder or peanut extract, either in the absence or presence of different dosages of AL extract. The AL extract was able to inhibit the β-hexosaminidase dose dependently in cells incubated with both CMA serum and milk powder (IC50 = 63.3 μg/ml), as well as in cells incubated with serum from peanut-allergic individuals and peanut extract (IC50 = 55.8 μg/ml; Fig. 3).
Degranulation in Human PBMCs.
Human PBMCs from 6 healthy donors were cocultured with AL extract and IL-3, which is critical for the development, survival, and function of tissue mast cells and blood basophils (40, 41, 42). Stimulation of basophils was induced by cross-linking of the FcεRI receptor with α-IgE antibody. Significant decrease (IC50 = 3.45 μg/ml) in degranulation was ascertained by a decrease of 7-AAD−/CD203c+ /CD63+ cells (Fig. 4a) and strikingly, biosynthesis of cys-leukotrienes was also decreased (IC50 = 3.54 μg/ml) (Fig. 4c). The release of β-hexosaminidase was inhibited, but not as much as degranulation and cys-leukotriene release, with IC50 = 13.9 μg/ml (Fig. 4b).
In vitro toxicologic aspects of AL were analyzed by 7-AAD, using flow cytometry (viability) and WST-1 (metabolic activity) to indicate the condition of the PBMCs. Viability was decreased to 80%, compared to the viability of untreated cells with 10.0 μg/ml of AL, but no IC50 value could be calculated (Fig. 4d). The metabolic activity of PBMCs was decreased (IC50 = 12.5 μg/ml) (Fig. 4e), but this value was significantly higher than the IC50 values of degranulation or cys-leukotriene release. Thus, the AL extract strongly decreased degranulation and cys-leukotriene excretion of human basophils without affecting the condition of the PBMCs negatively at the same dosage.
Activity-Guided Fractionation and Identification of the Active Component.
Thirteen fractions of AL extract H (fractions 1–13) were tested with the RBL-2H3 cells for their ability to inhibit degranulation. Only fraction 11, with retention volume 22.9 ml, was found to inhibit β-hexosaminidase release, with IC50 = 22.2 μg/ml (Fig. 5). Lyophilized complete extract run through the HPLC column and the complete extract without any processing was able to inhibit the release of β-hexosaminidase, with IC50, respectively 23.6 and 19.1 μg/ml (data not shown). The other extracts (extracts A–G) from different suppliers were also run through the HPLC column, and the peak found in fraction 11 of extract H was absent from all of the other extracts (data not shown). Attempts to identify the active component from fraction 11 by liquid chromatography-mass spectrometry (LC-MS) and nuclear magnetic resonance (NMR) were unsuccessful. One of the active components of AL described in the literature as being immunomodulatory is arctiin (19, 20, 21, 22, 23, 24). However, arctiin (Calbiochem, Darmstadt, Germany), in concentrations of 0.39, 0.78, 1.56, 3.13, 6.25, 12.5, 25, and 50 μg/ml, was not able to reduce the release of β-hexosaminidase in stimulated RBL-2H3 cells (data not shown).
Effect of AL on Acute Ear Swelling.
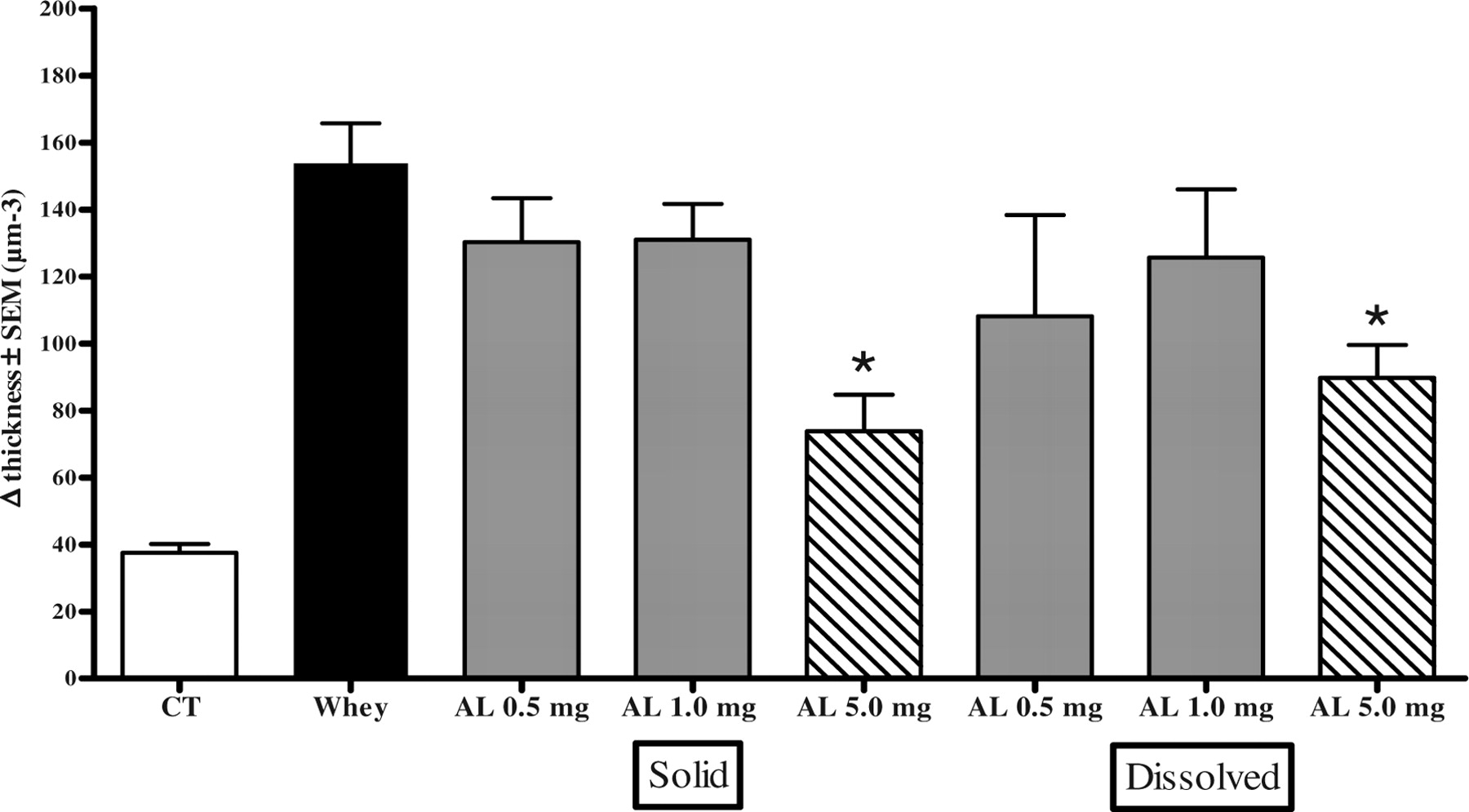
Topical administration of AL extract, both directly added to the vehicle cream as well as first dissolved in water and then added to the cream, in a concentration of 5 mg/ear, significantly inhibited acute mouse ear edema (reduction of swelling was 52% and 42%, P = 0.0011 and P = 0.0033, respectively) in whey-sensitized mice that were challenged with whey (Fig. 6). Oral administration of AL, in a concentration of 5 mg/0.5 ml PBS, showed no effect on ear swelling (data not shown).
The results of the current study demonstrate that the H extract of AL, selected by high throughput screening, displayed marked anti-allergic activity in vitro and in vivo. Topical treatment with AL inhibited acute mouse ear edema in whey-allergic sensitized mice that were challenged with whey. In contrast, oral administration of AL showed no effect on the acute allergic response in allergic animals. Moreover, the AL extract was able to inhibit IgE-mediated β-hexosaminidase release from RBL-2H3 and RBL-hEIa-2B12 cells and significantly reduced degranulation and cys-leukotriene production from activated basophils in human PBMCs. It is noteworthy that AL extract has a wide range of anti-inflammatory effects distinct from these anti-allergic effects, including potent inhibition of carrageenan-induced rat paw edema (17) and an inhibitory effect on platelet-activating factor (18).
In previous experiments (data not shown), no differences were found in degranulation when cells were incubated with AL solely during incubation of the cells with IgE, during cross-linking, or during the entire experiment. This indicates that the AL component does not interfere with either the FcεRI receptor or the IgE molecules themselves.
In this study, we found that AL extract has an inhibitory action on both mast cell degranulation and cys-leukotriene production. The signaling cascade triggered upon FcεRI cross-linking is induced by activation of protein-tyrosine kinases (PTKs) of the Src family. This activation is one of the earliest events detectable after antigen cross-linking of the IgE receptor. At least two families of PTKs are implicated as being responsible for the increase in tyrosine phosphorylation seen after FcεRI activation. Lyn, a member of the Src-like family of kinases, is reported to be associated with the receptor and to be activated following aggregation of FcεRI. In addition, the Syk tyrosine kinases induce tyrosine phosphorylation, and their activity is increased following receptor activation (43). Downstream from Syk activation, the mitogen-activated protein kinase pathway leads to phospholipase A2 activation, an initial step in the production of arachidonic acid metabolites such as LTC4 and prostaglandin D2 (44). It has been demonstrated that Syk activation in the absence of FcεRI cross-linking is sufficient to induce degranulation, leukotriene production, and synthesis of cytokines IL-3 and tumor necrosis factor-α (45). Therefore, it might be suggested that the current AL extract is likely to inhibit mast cell activation by inhibition of Syk activation, for this would block degranulation as well as leukotriene production.
Several different extracts of AL were studied for their effects on the release of β-hexosaminidase by RBL-2H3 cells, of which only extract H proved to be highly effective. Out of the 13 HPLC fractions of AL extract H, only fraction 11 showed an inhibitory effect on degranulation. This fraction contains only one peak, which indicates that one structure is responsible for the inhibitory effects. Additionally, the lyophilized and reconstituted extract was still able to reduce β-hexosaminidase release, demonstrating that the active unknown structure is not volatile. Unfortunately, attempts to identify the active component from fraction 11 by LC-MS and NMR were unsuccessful.
Our data suggest that toxic effects of AL are not expected. The viability and metabolic activity of PBMCs after 18 hrs of coculture with this herbal extract were hardly affected. The assessed risk for the development of anaphylactic shock on exposure to AL is extremely small. Four cases of anaphylaxis due to the use of anti-inflammatory root plasters and the consumption of AL have been described (46, 47), but when taken into account that this plant is frequently used in many—especially Asian—countries, this number is negligible. Other members of the Compositae family were found to be carcinogenic in rats; however, no carcinogenic activity was observed with use of AL (48). In vivo animal studies should be conducted to determine whether high concentrations of the active component in AL also have no toxic effects.
The present in vivo experiments showed that topical administration of AL extract was able to inhibit acute mouse ear edema when oral administration seemed to be ineffective. This might be due to digestion of the active compound in the gastrointestinal tract, resulting in AL losing its anti-allergic action. However, only one oral dose was tested; further in vivo experiments, using different dosages, are required to exclude AL extract as an oral anti-allergic agent.
In conclusion, AL extract is able to strongly reduce in vitro, and significantly reduce in vivo, the release of inflammatory mediators by inhibition of degranulation and cys-leukotriene release. Therefore, AL seems to be a very promising natural component for the treatment of allergy.
Of approximately 10,000 herbal extracts (black dots), 99 reduce the release of β-hexosaminidase in RBL-2H3 cells for > 55% (cut offset as 45% of degranulation in comparison with untreated cross-linked cell, dashed line). Effect of eight AL extracts (A–H), extracted by different methods and derived from various suppliers, on the release of β-hexosaminidase in RBL-2H3 cells. α-DNP-IgE bound to FcεRI receptors of RBL cells were cross-linked by DNP-BSA followed by the induction of degranulation. The release of β-hexosaminidase by the AL extracts was calculated from degranulation of the maximal release (Max. release). Nonsupplemented cells that were not cross-linked were used as negative controls, indicating spontaneous release (Spont. release) of β-hexosaminidase. Effect of AL extract on the release of β-hexosaminidase in RBL-hEIa-2B12 cells. IgE from allergic sera binds to the human IgE receptor of RBL cells and was crosslinked with α-IgE (serum + α-hIgE) as a control for the presence of IgE in the serum or with an allergen: milk powder 500 ng/ml or peanut extract 50 ng/ml (serum + allergen). Different concentrations of AL extract were added to the cells with serum + allergen. The release of β-hexosaminidase of the AL extract was calculated from the degranulation of purified human IgE crosslinked with α-human IgE (hIgE + αhIgE). Spont. release of β-hexosaminidase was measured with untreated cells, and the influence of serum on β-hexosaminidase is shown (serum 1/50). Effects of AL extract on isolated human PBMCs. PBMCs were cocultured with IL-3 and AL extract for 18 hrs. Metabolic activity was determined by WST-1. After 40 mins of FcεRI cross-linking, part of the cells were analyzed by flow cytometry. Markers used were α-CD203c-PE to indicate basophils and α-CD63-FITC as a marker for degranulation. Supernatants were analyzed for the presence of β-hexosaminidase and cys-leukotrienes. The average results of six donors are plotted. (A) Degranulation of basophils. (B) Release of β-hexosaminidase of basophils. (C) Release of newly synthesized cys-leukotrienes in supernatant after FcεRI cross-linking. (D) Viability of PBMCs measured by 7-AAD staining by flow cytometry. (E) Metabolic activity of PBMCs after coculture with IL-3 and herbal extract, but before FcεRI cross-linking. (F) Table of concentration at which IC50 can be calculated; values > 10 μg/ml were extrapolated. Effect of 13 HPLC fractions of AL extract on the release of β-hexosaminidase in RBL-2H3 cells. α-DNP-IgE bound to FcεRI receptors of RBL cells was cross-linked by DNP-BSA, followed by the induction of degranulation. The release of β-hexosaminidase of the HPLC fractions was calculated from the Max. release. Nonsupplemented cells that were not cross-linked served as negative controls, indicating Spont. release of β-hexosaminidase. Effect of AL extract (0.5, 1.0, and 5.0 mg/ear) in whey-induced ear edema in mice. AL was administered to the ear under two different conditions: AL extract directly added to the vehicle cream (Solid) and AL extract first dissolved in water before adding to the cream (Dissolved). Negative controls were animals receiving only CT and positive controls were whey-sensitized animals (Whey). Each data point represents the mean reduction of ear thickness of 5 animals ± SEM. * P < 0.01.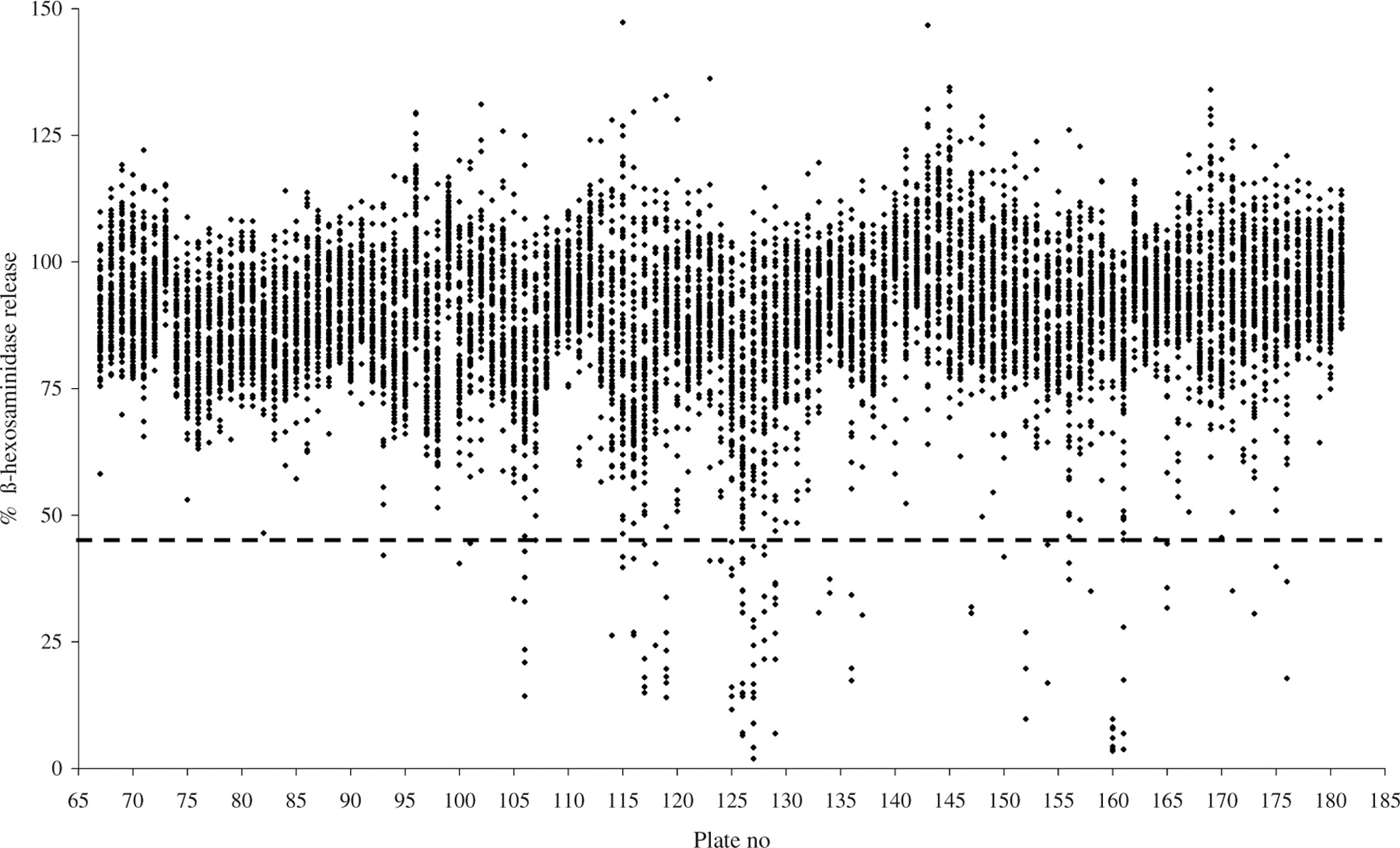




Footnotes
This work was supported by SenterNovem, TSIN 1055.
Acknowledgements
We thank D. Smeenge for her excellent assistance in developing and performing the high throughput screening, and G. Witte and E. Voogd for performing the HPLC experiments.