
Abstract
Neonates with premature rupture of the membrane and oligohydramnios have an increased risk of acute respiratory morbidity. The aims of this study are to investigate the effects of experimental oligohydramnios on transforming growth factor (TGF)-β1 and connective tissue growth factor (CTGF) expressions and collagen level in fetal rat lungs. On day 16 of gestation, we anesthetized timed pregnant Sprague-Dawley dams, punctured the uterine wall and fetal membranes of each amniotic sac which resulted in oligohydramnios. Fetuses in the opposite uterine horn served as controls. On days 19 and 21 of gestation, fetuses were delivered by cesarean section. Rats exposed to oligohydramnios exhibited significantly lower lung weight/body weight ratios on days 19 and 21 of gestation than did the control rats. Lung type I collagen and TGF-β1 mRNA expressions and lung collagen levels were significantly decreased in rats exposed to oligohydramnios on days 19 and 21 of gestation. Type I collagen and inhibitors of metalloproteinase-1 (TIMP-1) proteins were decreased and matrix metalloproteinase-1 (MMP-1) was increased in oligohydramnios-exposed rats on days 19 and 21 of gestation. CTGF mRNA expressions were comparable between control and oligohydramnios-exposed rats on days 19 and 21 of gestation. These data suggest that downregulation of collagen might be involved in the pathogenesis of oligohydramnios-induced respiratory morbidity.
Introduction
Pulmonary hypoplasia is common in the perinatal period and is a significant cause of death in newborn infants and oligohydramnios is one of the most commonly associated abnormalities (1). Oligohydramnios may retard fetal lung growth and can result in pulmonary hypoplasia in experimental animals and in human fetuses with prolonged rupture of the membrane (2–6). Neonates exposed to oligohydramnios caused by premature rupture of the membrane have an increased risk of acute respiratory morbidity, including higher ventilator settings, increased incidences of hypoxemia and hypercapnia, pulmonary hypertension, and a trend to more air leaks (7). That study indicated that oligohydramnios has considerable impacts on short-term respiratory morbidity in infants. However, the exact mechanism by which oligohydramnios alters the respiratory system remains unknown.
Physical forces produced by fetal breathing movements and lung fluid in the airspaces play important roles in regulating fetal lung growth and maturation (8, 9). The fluid maintains the lungs in an expanded state and provides the tissue with the mechanical stretching necessary for normal lung development (10). Mechanical stretching increases growth factor expression and collagen synthesis and secretion in lung fibroblast culture (11–13). We found decreased lung tropoelastin gene expression and elastin level in oligohydramnios-exposed fetal rats (14). However, little is known about the effects of oligohydramnios on the lung collagen status in vivo. The accumulation of collagen in lung tissue is controlled by various growth factors and a dynamic equilibrium between synthesis and degradation of collagen is maintained. This balance is controlled by de novo synthesis of collagens, proteolytic degradation by matrix metalloproteinase (MMP), and inhibition of MMP activity by tissue inhibitors of metalloproteinase (TIMP) (15). Transforming growth factor (TGF)-β1 is a potent stimulant of collagen synthesis in fibroblast cultures; it activates collagen promoters and increases the production of types I and III collagen (16, 17). Connective tissue growth factor (CTGF) functions as a downstream mediator of TGF-β1 (18). CTGF has been implicated in fibroblast proliferation, cellular adhesion, and extracellular matrix synthesis (19). In this study, we hypothesized that experimental oligohydramnios would alter TGF-β1 and CTGF expressions and hence the collagen level in fetal rat lungs.
Materials and Methods
Animals and Treatment
This study was approved by the Animal Care and Use Committee of Taipei Medical University and was performed with timed pregnant Sprague-Dawley rats. On day 16 of gestation, pregnant dams were anesthetized with pentobarbital (50 mg/kg, i.p.). An abdominal midline incision was made, and the two uterine horns were exposed and kept moist with phosphate-buffered saline (PBS). The uterine wall and fetal membranes of each amniotic sac in one horn were punctured using a 19-gauge needle, which resulted in immediate visible leakage of amniotic fluid and a decrease in the size of the amniotic sac. Fetuses in the opposite uterine horn served as controls. The uterus was returned to the abdomen and the abdominal incision was repaired. On days 19 and 21 of gestation, the dams were anesthetized with pentobarbital (50 mg/kg, i.p.), and the fetuses were delivered by cesarean section. The fetuses were weighed, and then killed by pentobarbital (100 mg/kg, i.p.); the lungs were removed, dissected free, and weighed.
Reverse Transcription PCR
Left lung tissue was ground into a powder in liquid nitrogen, and gene expressions were measured with a reverse transcription polymerase chain reaction (RT-PCR). Total RNA was extracted using the TRIzol Reagent (Invitrogen Life Technologies, Paisley, UK) according to the manufacturer’s instructions. The PCR were carried out with the primers shown in Table 1. The PCR products were analyzed by electrophoresis on an agarose gel, stained with ethidium bromide, and photographed. Five samples on each gestational day were analyzed for each gene in each group.
Western Blot Analysis
Lung tissues were homogenized in Tris·buffer with protease inhibitor cocktail tablets. The samples were sonicated and centrifuged at 500 g for 20 min at 4°C to remove cellular debris. Proteins (40 μg) were resolved on 10% SDS-PAGE under reducing conditions and electroblotted to a polyvinylidene difluoride membrane. After blocking with 5% non-fat dry milk, the membranes are incubated with anti-collagen type I (1:500; Calbiochem, San Diego, CA, USA), anti-MMP-1 (1:400; Thermo Fisher Scientific Inc., Warm Springs Blvd, Fremont, CA, USA), anti-TIMP-1 (1:100; Santa Cruz Biotechnology, Santa Cruz, CA, USA) or anti-β-actin (1:10,000; Sigma-Aldrich, St. Louis, MO, USA) and subsequently with horseradish peroxidase-conjugated goat anti-rabbit IgG or anti-mouse IgG (Pierce Biotechnology, Rockford, IL, USA). Protein bands are detected using SuperSignal Substrate from Pierce.
Lung Collagen
Lung collagen levels were measured by a Sircol collagen assay (Biocolor, Belfast, UK) as per the manufacturer’s instructions and modifications from Philips et al. (20). The collagen concentration on each gestational day was obtained as an average of five samples.
Immunohistochemistry of TGF-β1 and CTGF
Immunohistochemical staining was performed on paraffin sections with immunoperoxidase visualization. Sections for antibody against CTGF were first preincubated for 1 h at room temperature in 0.1 M PBS containing 10% normal goat serum and 0.3% H2O2 to block endogenous peroxidase activity and nonspecific binding of the antibody before being incubated for 20 h at 4°C with a primary rabbit antibody against CTGF (1:500; Abcam, Cambridge, UK). Sections for antibody against CTGF were incubated in 10 mM citrate buffer (pH 6.0) for 10 min at 95–99°C in a water bath. Thereafter, sections were blocked in 5% normal goat serum in PBS/Triton for 60 min and sections were incubated for 20 h at 4°C with a primary rabbit antibody against TGF-β1 (1:100; Cell Signaling Technology, Boston, MA, USA). After washing all of the sections were then treated for 1 h at room temperature with biotinylated goat anti-rabbit IgG (1:200; Vector Laboratories, CA, USA). This was followed by reaction with the reagents from an ABC kit (Avidin-Biotin Complex, Vector Laboratories, Burlingame, CA, USA) as per the manufacturer’s recommendations. After thorough washing, the final detection step was carried out with the use of 3,3′-diaminobenzidine (Sigma) as the chromogen. Sections for antibody against TGF-β1 were counterstained with hematoxylin.
Quantification of TGF-β1 and CTGF Immunoreactivities
At least 3 immunohistochemically stained sections per animal were analyzed and a minimum of 4 random lung fields per section at ×400 magnifications were randomly selected and captured with a digital camera and imported into the computerized image analysis system Image-Pro Plus. The automatic object counting and measuring process was used to quantify the immunoreactivity in the sections (21). This generated a percentage of positive stained cells and the values were expressed as labeling index (%).
Statistics
Results are presented as the mean ± SEM. Comparisons between control and oligohydramnios groups at equivalent gestational ages were made using unpaired Student’s t test. Differences were considered significant at P < 0.05.
Results
Four and six pregnant dams were used on days 19 and 21 of gestation, respectively. Control fetuses were all alive on days 19 (n = 20) and 21 (n = 31) of gestation. Twenty-six of 30 (86.7%) and 24 of 50 (48%) oligohydramnios-exposed fetuses survived on days 19 and 21 of gestation, respectively. Only surviving fetuses were used for examination.
Body Weight, Lung Weight, and the Lung/Body Weight Ratio (%)
Rats exposed to oligohydramnios exhibited significantly lower lung/body weight ratios on day 19 of gestation and lower lung weights and lung/body weight ratios on day 21 of gestation when compared with control rats (Table 2).
CTGF, Collagen, and TGF-β1 mRNA Expressions
In this study, we measured only type I collagen mRNA expression because it constitutes greater than 65% of the total lung collagen. Rats exposed to oligohydramnios had significantly lower type I collagen mRNA expressions on days 19 and 21 of gestation when compared with control rats (Fig. 1). CTGF mRNA expressions were comparable between oligohydramnios-exposed and control rats on days 19 and 21 of gestation (Fig. 1). TGF-β1 mRNA expressions were significantly lower in oligohydramnios-exposed rats when compared to the control rats on days 19 and 21 of gestation (Fig. 2).
Western Blot Analysis of Type I Collagen, MMP-1, and TIMP-1
Lung type I collagen and TIMP-1 proteins were decreased and MMP-1 was increased in oligohydramnios-exposed rats when compared to the control rats on days 19 (Fig. 3A) and 21 (Fig. 3B) of gestation.
Lung Collagen Level
The lung collagen level was significantly lower in the rats exposed to oligohydramnios when compared to control rats on days 19 (218.5 ± 12.4 versus 425.2 ± 61.0 μg/mg protein, P < 0.05) and 21 (314.9 ± 14.6 versus 466.7 ± 41.5 μg/mg protein, P < 0.05) of gestation (Fig. 4).
Immunohistochemistry for CTGF and TGF-β1
The CTGF protein was localized mainly in airway epithelium, and the immunoreactivity had increased in control and oligohydramnios-exposed rats on day 21 of gestation when compared with rats on day 19 of gestation (Fig. 5A). CTGF immunoreactivities were comparable between control and oligohydramnios-exposed rats on days 19 and 21 of gestation (Fig. 5B). TGF-β1 was mainly immunolocalized in the airway epithelium and rarely in the parenchyma (Fig. 6A) and the immunoreactivity was significantly decreased in oligohydramnios-exposed rats on days 19 and 21 of gestation (Fig. 6B). Changes in the immunoreactivity of CTGF and TGF-β1 in all four groups were similar to those of their mRNA expressions.
Discussion
Oligohydramnios may retard fetal lung growth and result in pulmonary hypoplasia (2, 4–6). Pulmonary hypoplasia is defined as a low lung weight for a given body weight. Moessinger et al. performed amniocentesis on day 15 of gestation in fetal rat and found a significant reduction in lung weight and lung/body weight ratio, reduced DNA per lung, and the lung protein to DNA ratio on day 21 of gestation (2). Blachford et al. produced oligohydramnios from days 16 to 21 of gestation and found significant reductions in lung weight and lung/body weight ratio, and comparable DNA and protein concentrations (4). Kitterman et al. punctured fetal rat amniotic sacs on day 16 of gestation and found a significant reduction in lung weight, lung/body weight ratio, and total DNA and protein contents on day 21 of gestation (6). The common finding of pulmonary hypoplasia-induced by oligohydramnios is reduced lung weight and lung/body weight ratio in those studies. In the present study, we produced oligohydramnios on day 16 of gestation and found significant reductions in lung/body weight ratio and lung development on days 19 and 21 of gestation. Therefore, oligohydramnios did produce pulmonary hypoplasia based on lung/body weight ratio and histological findings.
TGF-β1 is a potent stimulant of collagen synthesis and production (16, 17). CTGF has been implicated in fibroblast proliferation and extracellular matrix synthesis (19). CTGF is a promoter of collagen deposition that acts downstream of TGF-β1, particularly with respect to its profibrotic effects (22). In this study, we found decreased TGF-β1 and type I collagen expressions and comparable CTGF expressions on days 19 and 21 of gestation. These data were compatible to the results of Kessler et al., who found that TGF-β did not stimulate CTGF expression when added to relaxed cells, and no temporal correlation was noted between the expressions of CTGF and TGF-β in stressed or relaxed fibroblasts (23). These results suggest that the signaling cascade activated by oligohydramnios does not involve an autocrine loop of CTGF.
Two major stimuli to fetal lung growth result from mechanical stretching produced by fetal breathing movements and a constant distension pressure (9). Oligohydramnios does not influence fetal breathing movements, but it does decrease the volume of fluid within the potential airways and airspaces (5, 24). Hyaluronan is the major interstitial matrix controlling water mobility and water balance in the lung. In this study, oligohydramnios produced by needle puncture of rat amniotic sacs is mainly due to membrane rupture rather than to changes in lung parenchymal matrix as hyaluronan secretion and production is stimulated by joint expansion and cell stretch in fetal lung fibroblasts, respectively (25, 26). The fluid maintains the lungs in an expanded state and provides mechanical stretching for the tissue that is necessary for normal lung development (10). Mechanical stretching increases growth factors expressions through intracellular signal transduction pathways (11). We speculated that oligohydramnios causes a reduction in amniotic fluid volume and a decrement in lung distension, thus producing a smaller amount of mechanical stretching-induced TGF-β1 activation. This speculation was supported by Hori et al. (27), who found that anti-TGF-β neutralizing antibodies blocked the mechanical stretching-induced type I collagen mRNA expression in cultured rat mesangial cells. Our findings suggest that CTGF does not mediate TGF-β1-induced collagen reduction in the hypoplastic fetal rat lungs due to oligohydramnios.
Collagen is vital for maintaining the normal lung architecture. Collagen fibrils are widely distributed in the interstitium of the bronchial tree, the interlobular septa, the bronchial lamina propria, and the alveolar interstitium. In this study, we found that oligohydramnios decreased lung collagen and TGF-β1 expressions in hypoplastic fetal rat lungs. This result is consistent with the observations of Ogata et al. (28), who found decreased TGF-β and collagen mRNA expressions after release of a urethral obstruction in rat kidneys. This study further supports our suggestion that pulmonary hypoplasia associated with oligohydramnios results from a reduction in fetal lung expansion. MMP-1 is an interstitial collagenase that specifically degrades type I collagen and TIMP-1 is a specific inhibitor of MMP-1. An imbalance of MMP and TIMP can result in metalloproteinase activation and a relatively higher level of MMP than TIMP may stimulate the degradation of collagen in the interstitial space. During the pathogenesis of oligohydramnios, the homeostasis deteriorates, resulting in a net decrease in deposited collagen content of the lung. This altered extracellular matrix might be responsible for the alteration of respiratory function associated with oligohydramnios.
In conclusion, maternal oligohydramnios during late gestation decreased TGF-β1 and collagen expressions in hypoplastic fetal rat lungs. These data suggest that down-regulation of collagen seems to be involved in the pathogenesis of oligohydramnios-induced respiratory morbidity and this effect was independent of CTGF.
Oligonucleotide Sequences of the Primers Used
Body Weight, Lung Weight, and Lung/Body Weight Ratio in the Control and Oligohydramnios-Exposed Rats

Effects of oligohydramnios on CTGF and collagen I mRNA expressions. Oligohydramnios-exposed rats exhibited significantly decreased collagen I mRNA expressions on days 19 and 21 of gestation and showed no significant effect on CTGF mRNA expressions when compared with control rats. ** P < 0.01 compared with the control group on each gestational day.

Effects of oligohydramnios on TGF-β1 mRNA expressions. Oligohydramnios-exposed rats exhibited significantly decreased TGF-β1 mRNA expressions when compared with control rats on days 19 and 21 of gestation. *** P < 0.001 and ** P < 0.01 compared with the control group on day 19 and 21 of gestation, respectively.

Representative Western blots for collagen type I, MMP-1, and TIMP-1 proteins in fetal rat lung exposed to oligohydramnios on days (A) 19 and (B) 21 of gestation.
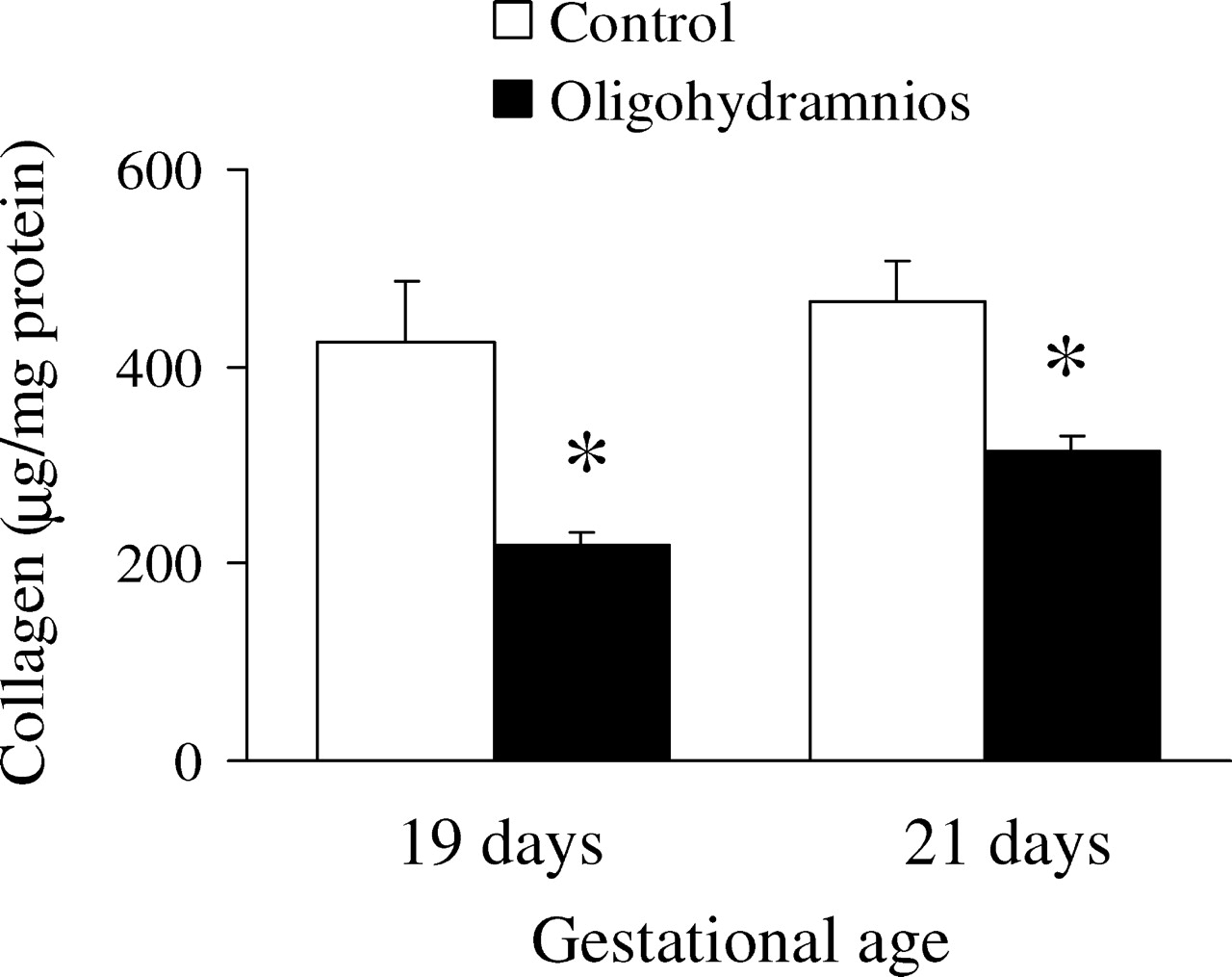
Collagen levels in fetal rat lungs. Oligohydramnios-exposed rats exhibited significantly decreased lung tissue collagen levels when compared with control rats on days 19 and 21 of gestation. * P < 0.05 compared with the control group on each gestational day.
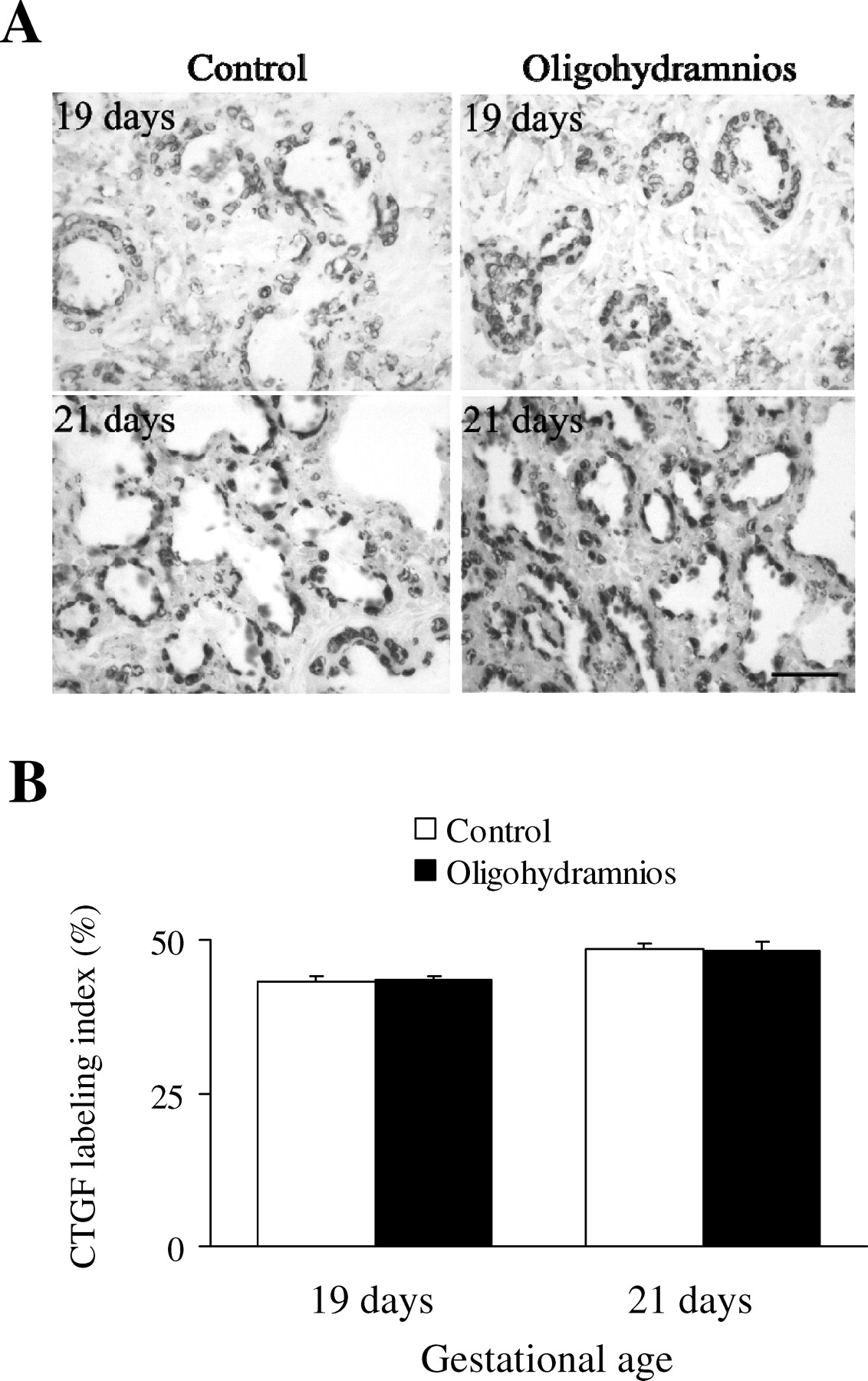
(A) Immunolabeling of CTGF in lung sections from control and oligohydramnios-exposed rats (×400). Positive staining is shown as brown. The CTGF protein was mainly localized in airway epithelial and the immunoreactivity was comparable between control and oligohydramnios-exposed rats on days 19 and 21 of gestation. Bar, 50 μm. (B) Quantitative analysis of CTGF immunoreactivity in the control and oligohydramnios-exposed rats. CTGF immunoreactivities increased in control and oligohydramnios-exposed rats on day 21 of gestation when compared with rats on day 19 of gestation and were comparable between control and oligohydramnios-exposed rats on days 19 and 21 of gestation.

(A) Immunolabeling of TGF-β1 in lung sections from control and oligohydramnios-exposed rats (×400). Positive staining is shown as brown. The TGF-β1 immunoreactivity was reduced in oligohydramnios-exposed rats on day 19 and 21 of gestation. Bar, 50 μm. (B) Quantitative analysis of TGF-β1 immunoreactivity in the control and oligohydramnios-exposed rats. TGF-β1 immunoreactivities were significantly decreased in oligohydramnios-exposed rats on days 19 and 21 of gestation when compared with control rats. *** P < 0.001 compared with the control group on each gestational day.