
Abstract
The purpose of the present study was to confirm the effectiveness of saponin hemolysis for concentrating ring-infected erythrocytes in Plasmodium falciparum cultures and to determine the actual numbers of the enriched parasites, not just percentage parasitemia. This is important because various molecular biology and vaccine development against malaria require useable quantities of pure culture with minimal number of uninfected erythrocytes at all stages. Synchronized cultures of three P. falciparum strains were exposed to 0.015% isotonic saponin solution for 30 minutes on ice. They were centrifuged and the pellets were treated again with saponin solution for 3–7 minutes. Initially, most of the cultures contained approximately 1010 erythrocytes and 1–7% parasitemia, but at the end of the enrichment up to 108 of erythrocytes containing 90–99.8% parasitemia were recovered (maximal enrichment). From microscopic examination of the cells it was calculated that the hemolysis rate of uninfected and infected erythrocytes was circa 27 to 1, which could account for the enrichment. Studies by other investigators have suggested that P. falciparum merozoite invasion decreases erythrocyte membrane lipids, and it has been reported that reduction of membrane cholesterol could make erythrocytes saponin-resistant. The possibility that merozoite invasion made erythrocytes partially resistant to saponin hemolysis was strengthened by the observation that the proportions of multiple infections increased significantly in the enriched cultures. However, mature asexual parasites could not be concentrated by this method, suggesting possible differences between the membranes of erythrocytes containing ring forms and those of trophozoites and schizonts. Ring-infected erythrocytes freshly from malaria patients could also not be concentrated by the method described here, suggesting that the ability to induce saponin resistance in erythrocytes was acquired by the parasites in vitro.
Introduction
In addition to the public health and economic misery that malaria inflicts upon human population in endemic areas (1, 2), the disease is increasingly being imported into non-endemic countries of Europe (3), the USA (4) and the affluent Gulf Region of the Middle East (5–7). There is no malaria transmission in Kuwait, but according to an article in a local newspaper (Khaleej Time, 30 March 2008), up to 69% of the population in 2007 was from foreign countries, mostly from malaria endemic areas. Plasmodium falciparum is responsible for most of malaria deaths with estimated annual clinical episodes of 300–660 million cases worldwide, based on the year 2002 data (8). It is therefore, not surprising that the majority of the research efforts now going on worldwide on molecular biology, chemotherapy and vaccine development in different laboratories have focused on P. falciparum, aided by in vitro cultivation of the parasite (9).
The stages in the life cycle of P. falciparum in humans are well defined, beginning with the sporozoites that initiate infection in the liver, merozoites that infect erythrocytes where the parasite goes through asexual multiplication (schizogony) via the ring, trophozoite and schizont stages, as well as sexual maturation from ring to I–V gametocyte stages (10). Unlike P. vivax, P. malariae and P. ovale in which all the intraerythrocytic stages in their life cycles are detectable in human peripheral blood, P. falciparum circulates mainly at the ring and mature gametocyte stages, except in relatively rare infections where all the stages could also be detected. A clear understanding of the different stages in the life of the blood forms of P. falciparum and the changes they induce in host erythrocytes has been tremendously facilitated by in vitro cultivation of the parasites. When first collected from infected human blood, the parasites that subsequently grow and multiply in vitro are all at the ring stage of development. However, after about four days in culture, all the asexual stages and maturing gametocytes could be identified in Giemsa-stained smears and by the end of the 4th–5th asexual cycle (day 8–10) mature gametocytes could also be detected among the different asexual forms (10).
The asynchronous growth of P. falciparum in culture was initially problematic for the scientists who wanted to work with defined stages of the parasite, until a synchronization technique was developed by Lambros and Vanderberg (11). Sorbitol destroys trophozoites and schizonts, leaving the ring-infected and uninfected erythrocytes. The parasites develop synchronously for 1–3 asexual cycles before the growth becomes asynchronous again. In addition, only 10–30% parasitemia can routinely be achieved in continuous cultures (12, 13) and uninfected erythrocytes could complicate some research experiments. Techniques that could enrich trophozoites and schizonts to 75% or higher parasitemia levels were introduced to reduce the number of uninfected erythrocytes (14). Still, there are some experiments that may require the parasites to be at the ring stage of development and also have minimal contamination by uninfected erythrocytes. For example, measurement of the rate of hemoglobin consumption and other metabolic activities could be complicated by large proportions of uninfected erythrocytes in the culture.
A saponin hemolysis technique for enriching P. falciparum ring-infected erythrocytes was first reported in a brief correspondence to the journal Lancet in 1994 (15). In that report it was shown that synchronized P. falciparum HB-3 culture containing ≥ 0.4% parasitemia could be enriched to 99% parasitemia by controlled saponin hemolysis, but the actual number of infected erythrocytes that could be recovered after the hemolysis was not determined. In the present study three different P. falciparum strains grown in vitro in normal (hemoglobin AA) and sickle cell trait (hemoglobin AS) erythrocytes, as well as blood samples directly from infected humans, were examined in order to define the conditions that are vital for successful enrichment of ring infected erythrocytes, including the actual number of parasites that could be harvested by the technique. Regarding the mechanism of the enrichment, we have used the information published by other investigators to suggest that merozoite invasion decreases cholesterol content of erythrocyte membrane, making infected cells relatively resistant to saponin hemolysis.
Materials and Methods
Malaria Parasites.
The malaria cultures tested included P. falciparum strains HB3, PB112 and Amiri7. Most of the experiments were done using HB3 strain which is well known internationally. In contrast, PB112 and Amiri7 were isolated and adapted to continuous cultivation here in Kuwait (16). Some blood samples collected directly from infected humans were also subjected to saponin hemolysis without prior in vitro growth. The blood samples were initially for routine malaria microscopy in hospital laboratories and they were collected in tubes containing EDTA anti-coagulant. The hospital laboratories detected only ring forms in the samples but submitted the “left over” blood specimens (1–2 ml) to the author of this paper for DNA analysis using polymerase chain reaction (PCR) in order to exclude mixed infections. Approximately 80% of the imported malaria cases in Kuwait are usually P. vivax and almost the rest are P. falciparum. Imported P. malariae and P. ovale are rare, but P. vivax and P. falciparum mixed infections are common.
In Vitro Culture of P. falciparum.
HB3, PB112 and Amiri7 strains were maintained in continuous culture, using stored transfusion blood from blood banks. Except where stated otherwise, the blood samples contained HbAA erythrocytes, as determined by sickling test (16). The standard method for continuous cultivation of P. falciparum in vitro was followed throughout, including incubation of all the cultures in a gas mixture containing 3% oxygen, 4% carbon dioxide and 93% nitrogen. RPMI 1640 powder, HEPES, hypoxanthine and gentamicin were obtained from GIBCO, BRL (Life Technologies). Each liter of the growth medium was supplemented with 2 g of D-glucose, 50 mg of hypoxanthine, 20 mg of gentamicin and 10% of human serum (complete medium) and the solutions were sterilized by membrane filtration. The parasites were routinely grown in approximately 4–7% hematocrit suspension of erythrocytes, unless stated otherwise.
Synchronization of Parasite Growth.
P. falciparum cultures were synchronized using 5% sorbitol solution in distilled water (16). The parasites were usually grown for 2–4 asexual cycles before being subjected to saponin concentration.
Saponin Solution.
The saponin powder used in this study was purchased from Sigma (St. Louis, MO, USA). Data obtained from the company about the product indicated that it was produced from Quillaja bark and it contained 11.1% sapogenin, 15.1% sulfate ash and 43.9% carbon. The working solution used for hemolysis was prepared by dissolving 15 mg of the saponin in 100 ml of modified Krebs/Ringer phosphate (MKRP) buffer containing the following salts (nmol/l): NaCl, 68; Na2HPO4, 50; KCl, 4.8; MgSO4, 1.2, and was adjusted to pH 7.4 with HCl. After preparation, the 0.015% saponin solution was sterilized by filtration through a membrane filter, 0.2 μm pore size, and was stored refrigerated until used. In most cases the saponin solution was used within 24 h, but a few experiments were also successfully done with saponin solution that was kept refrigerated for up to 3 months.
Saponin Hemolysis of P. falciparum Culture.
Sufficient aliquots of 2–6 days old P. falciparum culture containing 1–7% parasitemia were synchronized by sorbitol hemolysis and the pellets were pooled and centrifuged for 10 min in a 50-ml plastic centrifuge tube to obtain approximately 3 ml pellet containing 1–6 × 1010 erythrocytes. Twenty-eight ml of cold 0.015% saponin solution was added to the tube and the cell suspension was quickly and thoroughly mixed by hand. Four aliquots, each containing 7 ml of the suspension were prepared in 15-ml plastic centrifuge tubes and incubated on ice for 30 min with occasional mixing. The tubes were centrifuged for 5 min and the pellets were pooled in one tube and suspended in 7 ml of complete medium. After centrifugation for 5 min the supernatant was discarded and the pellet was suspended again in 5 ml of 0.015% saponin solution and incubated on ice for approximately 5 min. The suspension was centrifuged for 5 min and the pellet was washed 3 times by suspending it in 7 ml of complete medium and centrifuging for 5 min each time. Unless stated otherwise, all centrifugations were at 3000 rpm at room temperature. Throughout the procedures precautions were taken to avoid bacterial and fungal contamination of the parasite cultures.
Calculation of Hemolysis Rate.
The numbers of infected erythrocytes before and after the enrichment were calculated using the percentage parasitemia, and these were, respectively, subtracted from the total numbers of erythrocytes before and after saponin treatments. The number of hemolyzed infected erythrocytes was calculated by subtracting the number of infected erythrocytes after the enrichment procedure from the number of infected erythrocytes before the synchronized culture was first exposed to saponin. A similar calculation was made to obtain the number of hemolyzed uninfected erythrocytes. Finally, the hemolysis rate was calculated by dividing the number of hemolyzed uninfected erythrocytes by the number of hemolyzed infected erythrocytes.
In Vitro Growth of Enriched Parasites.
When necessary, the enriched parasites were allowed to continue their growth in vitro not only to monitor their viability microscopically but also to test their metabolic activity by measuring hemozoin production. For this purpose, the parasites were suspended in 2 ml of complete medium in high speed Oak Ridge centrifuge tubes with caps and were incubated in low oxygen environment as has been described previously (16). Usually the total numbers of cells in the cultures were in the order of 107.
Measurement of Hemozoin Production.
The method for quantitative measurement of hemozoin production by P. falciparum parasites has been described in another publication (16, 17).
Saponin Hemolysis of Trophozoites and Schizonts.
P. falciparum growth was synchronized through sorbitol hemolysis at the ring stage. The parasites were maintained in culture for 22 h to produce trophozoites and schizonts. The culture was centrifuged for 5 min and the pellet was suspended thoroughly in 7 ml of cold 0.015% saponin solution. After 30 min of incubation on ice the suspension was centrifuged for 5 min and the pellet was washed 3 times with 7 ml of complete medium. Smears of the final pellet were prepared on glass slides for staining with Giemsa stain and microscopic examination.
Saponin Hemolysis of Falciparum Malaria Blood Samples Directly from Infected Humans.
One ml of blood from human infected with P. falciparum was transferred to a sterile 15-ml plastic centrifuge tube with screw cap and centrifuged for 10 min to remove the plasma. The pellet, which contained both erythrocytes and leukocytes, was washed twice with 5 ml of saline. The volume of the pellet after the washing was usually 0.3–0.6 ml. It was loosened by gentle agitation before 4 ml of cold 0.015% saponin solution was added to the tube and quickly mixed by inverting and tapping at the bottom of the tube repeatedly until the cells have been suspended. The tube was then placed on ice and allowed to stand for 30 min, mixing occasionally to keep the cells in suspension and at the same time monitor the progress of hemolysis. After 30 min the tube was centrifuged for 5 min and the supernatant was quickly transferred to another sterile centrifuge tube. The pellet was suspended again in 2–5 ml of saponin solution, depending on the quantity of the cell pellet, and incubated on ice for up to 7 min. The suspension was centrifuged for 5 min, after which the supernatant was transferred to the tube containing the first supernatant and centrifuged for 30 min. Approximately 10 μl of the pellets were spread on glass slides and allowed to dry at room temperature for at least 1–2 h before fixing with methanol and staining with Giemsa stain.
Microscopic Examination for Malaria Parasites.
After the enrichment procedures, thin smears of the pellets were prepared and allowed to air dry at room temperature. The slides were then fixed with methanol and stained for 30 min with 10% Giemsa stain and after drying at room temperature the smears were examined under a microscope, using ×100 oil immersion objective. This was also the method used for examining blood samples and standard in vitro cultures for parasitemia. All parasitemias and parasite counts in this report refer only to parasites located in intact erythrocytes, unless stated otherwise.
Scanning Microscopy of Parasite in HbAS Erythrocytes.
One ml of P. falciparum HB3 culture in HbAS erythrocytes was injected into a vacutainer tube sealed with a rubber stopper. Inlet and outlet ports were created with two 18-gauge needles in the rubber stopper and the tube was flushed with the low oxygen gas mixture (3% oxygen, 4% carbon dioxide and 93% nitrogen) before removing the outlet and then the inlet needles. The tube was placed at a slant position in a modular incubator chamber and the chamber was sealed and filled with the low oxygen gas mixture as usual. After 24 h incubation at 37°C, and without exposure to air, the parasite culture was fixed with 2.5% (final concentration) of aqueous solution of glutaraldehyde for 60 min at room temperature as has been described (16). The suspension was centrifuged and thin smears of the pellet were prepared on glass slides. The smears were stored until electron microscopy could be done. For the scanning electron microscopy, a portion of the glutaraldehyde-fixed smear on glass slide was removed with a blade and mixed with 50% alcohol in distilled water on a mica sheet. The mixture was allowed to air-dry at room temperature and then was coated with gold, using Balzer’s sputter coater SCD50, after which the preparation was examined and photographed under Leo (Zeiss) 50 VP supra variable pressure field emission scanning electron microscope.
Statistical Analysis.
All statistical data were obtained using GraphPad PRISM® computer software.
Results
Parasite Cultures.
The P. falciparum HB3 strain used in this study did not produce any gametocyte throughout the period it was maintained in our laboratory, whereas both of the new isolates PB112 and Amiri7 consistently produced few gametocytes. Data presented here on HB3 and Amiri7 strains were obtained using HbAA erythrocytes, whereas PB112 strain was cultured in HbAS erythrocytes. As has been reported previously (16) and also shown in Figure 1, the low oxygen environment in which the parasites were grown caused some HbAS erythrocytes to sickle. However, scanning electron microscopy suggested that most of the heterozygous sickle cells apparently retained the morphology of normal erythrocytes (Fig. 1, left panel).
Enrichment of Ring-Infected Erythrocytes by Saponin Hemolysis.
Exposure to saponin was mostly carried out in two steps, the first step consistently lasting for 30 min, while the second step lasted in most cases 5 min. The results obtained with HB3 strain in a total of 32 experiments have been summarized in Figure 2. Parasitemia levels after the two-step hemolysis ranged from 76% to 99.8%. Specifically, 11 experiments yielded ≥ 99% parasitemia, 14 were between 90% and 98%, 6 were between 80% and 89%, and 1 experiment yielded 76%.
Hemolysis Rate and Parasite Yield.
The calculations of hemolysis rate were done using the data that were obtained with P. falciparum HB3 strain. From a total of 7 experiments it was determined that the mean hemolysis rate was 27 uninfected erythrocytes for 1 ring-infected erythrocyte hemolyzed. The percentages and absolute numbers of infected erythrocytes that were recovered in the experiments have been summarized statistically in Table 1.
Morphology of Enriched P. falciparum HB3 Ring Forms.
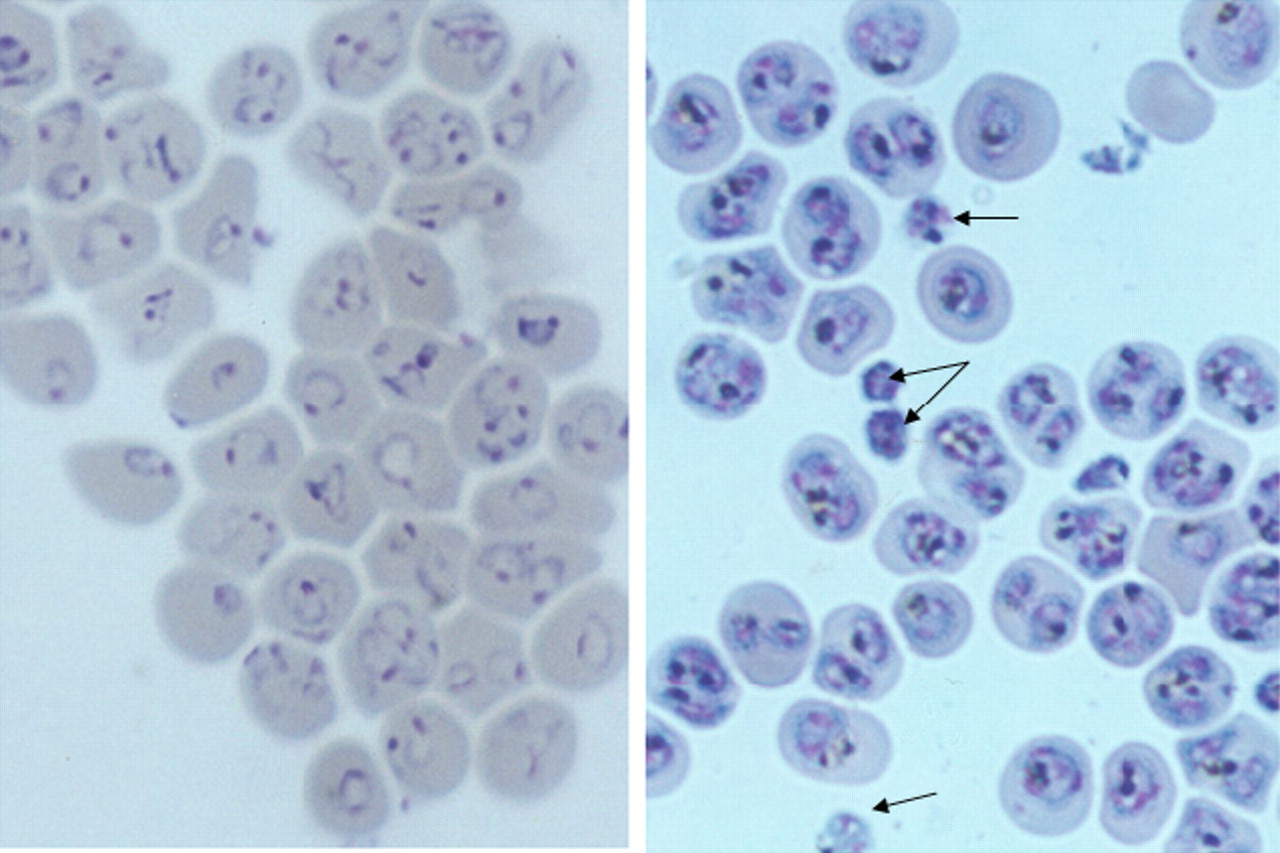
Microscopic examination of the culture in Giemsa stained thin smears did not reveal any identifiable abnormality of the parasites and the host erythrocytes after exposure to saponin. However, it was found that the mean proportion of multiply infected erythrocytes had increased from approximately 13% initially to 42% after the enrichment procedure (Table 2). Among the multiple infections the numbers of parasites in an erythrocyte ranged from 2 to 4 (Fig. 3), although a few erythrocytes containing 5 or even 6 parasites were detected (not included in the figure).
Viability and Metabolism of Saponin Enriched P. falciparum HB3 Ring Forms.
Saponin enriched intraerythrocytic ring forms were suspended in 2 ml of complete medium containing a total of 2–7 × 107 erythrocytes and incubated for up to 44 h in the standard conditions for in vitro growth, except that the cultures were in Oak Ridge centrifuge tubes to permit centrifugation and quantitative measurement of hemozoin production (17). After 24 h of incubation it was observed by microscopy that most of the parasites had matured to trophozoites and schizonts (Fig. 4). After a total of 42 h of growth the mean quantity of hemozoin produced by the parasites contained 975 pmol FP/106 infected erythrocytes, which was somewhat less than the quantity produced in unenriched cultures (Fig. 5). In other experiments in which enriched rings were first grown for 24 h and then mixed with normal erythrocytes and incubated for additional 24 h, it was determined that each of the enriched ring forms developed to a schizont which produced merozoites that infected approximately 5 new erythrocytes (data not shown).
Saponin Hemolysis of P. falciparum Trophozoites and Schizonts.
In 3 experiments with P. falciparum HB3 strain, the cultures were synchronized at the ring stage and allowed to grow for 24 h. They were then subjected to saponin hemolysis for 30 min, after which it was found by microscopic examination that the mean parasitemia levels had decreased from 4.5% before to 0.3% after the exposure to saponin. It must be stressed that only intraerythrocytic parasites were counted in these experiments. The few trophozoites and schizonts that survived the exposure to saponin appeared partially dehemoglobinized and did not grow further when the cultures were maintained for additional 24 h (data not shown).
Saponin Hemolysis of Erythrocytes Infected In Vivo with P. falciparum.
Blood samples were collected from 4 expatriate male adults who were probably infected in their home country, India, before coming to Kuwait. The blood samples contained only the ring forms, and polymerase chain reaction (PCR) with species specific primers confirmed that only P. falciparum was in the blood samples (18). The parasitemia levels were between 0.6% and 7.3% before saponin. Massive quantities of dehemoglobinized parasites were recovered from the supernatants of the saponin-treated blood samples (Fig. 6A), but the proportions of intact parasitized erythrocytes remained essentially the same as they were before saponin hemolysis (Fig. 6B). Detailed information about the usefulness of saponin hemolysis in malaria diagnosis will be published separately.
Enrichment of P. falciparum Rings in HbAS Erythrocytes.
In vitro cultures of P. falciparum PB112 strain were synchronized with sorbitol and subjected to saponin hemolysis as described in Materials and Methods. In the experiment presented in Figure 7, the parasitemia levels before and after synchronization with sorbitol were 7% and 4.5%, respectively. Immediately after the enrichment by saponin hemolysis the parasitemia increased to 82% (Fig. 7C), and was 79% after it was allowed to grow to mainly trophozoites and schizonts during 30 h incubation under the low oxygen conditions for in vitro growth (Fig. 7D).
Enrichment of P. falciparum Amiri7 Strain.
A synchronized culture of Amiri7 containing 2.7% parasitemia was exposed to saponin in two steps as usual, but the final parasitemia level was only 63%. The strain usually produced a few gametocytes during continuous cultivation and more of the gametocytes were detected after the treatment with saponin, indicating that the method also enriched the sexual form (Fig. 8).
Discussion
Saponin hemolysis for concentrating viable ring-infected erythrocytes was first developed in 1994, using in vitro culture of P. falciparum HB3 strain (15). Since then, several investigators have communicated with the author, stating that they encountered some difficulties while trying to use the method, but they did not specify what the difficulties were (personal communications). The questions that came to mind were whether resistance to saponin was strain-specific or whether the difficulties experienced by other investigators were perhaps because they apparently did not recover adequate number of parasites even though the percentage parasitemia increased dramatically after the saponin concentration procedure. To address the question of strain specificity, attempts were made with blood samples collected directly from 12 falciparum malaria patients and none of them could be significantly enriched by saponin hemolysis (Fig. 6). However, the enrichment was successful when in vitro cultures of two newly isolated P. falciparum strains, PB112 and Amiri7 were used. These new isolates had been continuously in culture for approximately two months before the enrichment experiments. PB112 was initially isolated from a sickle cell trait carrier and was since then cultivated continuously in HbAS erythrocytes. In addition to establishing that saponin hemolysis could also be used to enrich other P. falciparum strains, the experiment with PB112 has further confirmed that P. faciparum could be grown continuously in HbAS erythrocytes at low oxygen tension and synchronized by sorbitol hemolysis, just like the parasites in HbAA erythrocytes. On the issue of yield, it was determined with HB3 strain that up to 108 of apparently intact erythrocytes, containing up to 99% parasitemia and could support the completion of P. falciparum asexual cycle, could be recovered after exposure of circa 1010 erythrocytes, initially containing ≥1% parasitemia, to saponin hemolysis. The quantities of enriched infected erythrocytes were sufficient for studies such as quantitative measurement of hemozoin production and the level of hemoglobin consumption, as well as chloroquine accumulation by the parasites (data not shown).
Regarding why ring-infected erythrocytes were less susceptible than uninfected erythrocytes to saponin hemolysis, it has been determined by other investigators that during erythrocyte invasion P. falciparum merozoites translocate cholesterol-rich membrane components (detergent-resistant membrane, DRM) complexes from erythrocyte membrane for use in formation of the parasitophorous vacuole membrane (PVM) separating the parasite from the cytosol of the host cell (19–21). Other studies have reported that saponin makes cells permeable to macromolecules by forming complexes with membrane cholesterol (22) and that the susceptibility to saponin hemolysis was decreased by reducing erythrocyte membrane cholesterol level (23). Taken together, it is reasonable to suggest that P. falciparum ring-infected erythrocytes become partially resistant to saponin hemolysis because of the invasion by plasmodial merozoites and the removal of cholesterol-rich DRM from host erythrocyte membrane. The association of merozoite invasion with saponin resistance was made more plausible by the observation that the proportions of multiply infected erythrocytes increased after saponin hemolysis (Fig. 2), suggesting that multiple invasion decreases host cell membrane lipid level more than invasion by one parasite. Nevertheless, further studies are necessary to clearly establish the connection between merozoite invasion and resistance of host erythrocytes to saponin hemolysis. Such studies would seek to explain why only a relatively small proportion of the ring forms were resistant to saponin hemolysis and why the resistance was ineffective at the trophozoite and schizont stages. Furthermore, the role of the infected erythrocytes still has to be clearly defined because it has not yet been ruled out that the population of erythrocytes that survived the exposure to saponin already possessed that capability before parasite invasion. Under the conditions of the present experiments it was found that not all of the 1010 normal erythrocytes that were suspended in 0.015% saponin solution became completely hemolyzed within 30–35 min of incubation (data not shown).
For practical purposes, the enrichment technique described here could be summarized as follows: Ring-infected and uninfected erythrocytes in P. falciparum culture are hemolyzed at a rate that continuously increase the proportion of infected to uninfected cells until a point where only parasitized erythrocytes are left. For example, if 99.9% of the erythrocytes in a synchronized P. falciparum culture were hemolyzed, most of the remaining 0.1% would contain ring forms of the parasite, even when the initial parasitemia was about 1%. The actual number of infected erythrocytes after the procedure would depend on prompt termination of saponin action (by washing with serum-medium mixture, i.e., complete medium) to prevent further hemolysis. Among the things that should also be considered when using this technique is whether it is necessary to remove virtually all uninfected erythrocytes in the culture (maximal enrichment) or whether the intended experiments could be done with cultures containing about 50% parasitemia, in which case the saponin hemolysis procedure should be limited to the first 30 min, thereby reducing the loss of parasites.
Statistical Analysis of Parasite Yield After Saponin Enrichment a
Effects of Saponin Hemolysis on Parasitemia and Multiple Infections a
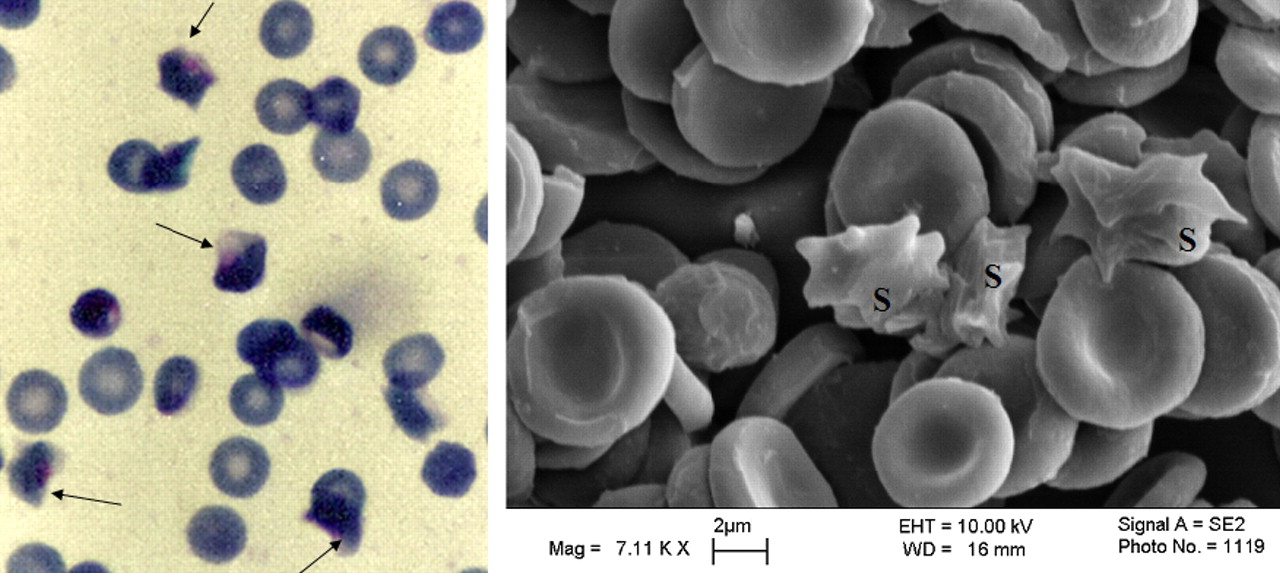
Effect of low oxygen environment on HbAS erythrocytes. P. falciparum culture in HbAS erythrocytes was incubated for 24 h in a mixture of 3% oxygen, 4% carbon dioxide and 93% nitrogen as described above, after which a solution of 2.5% glutaraldehyde was added to the culture to fix the cells before exposure to air. Thin smears of the fixed cells were stained with Giemsa stain and examined by light (left panel) or scanning electron microscopy (right panel). Although most of the sickled cells contained parasites in the stained smear, the parasites were not identifiable by scanning electron microscopy. The arrow at left panel and the S at right panel indicate erythrocytes that sickled during incubation at low oxygen tension. A color version of this figure is available in the online journal.
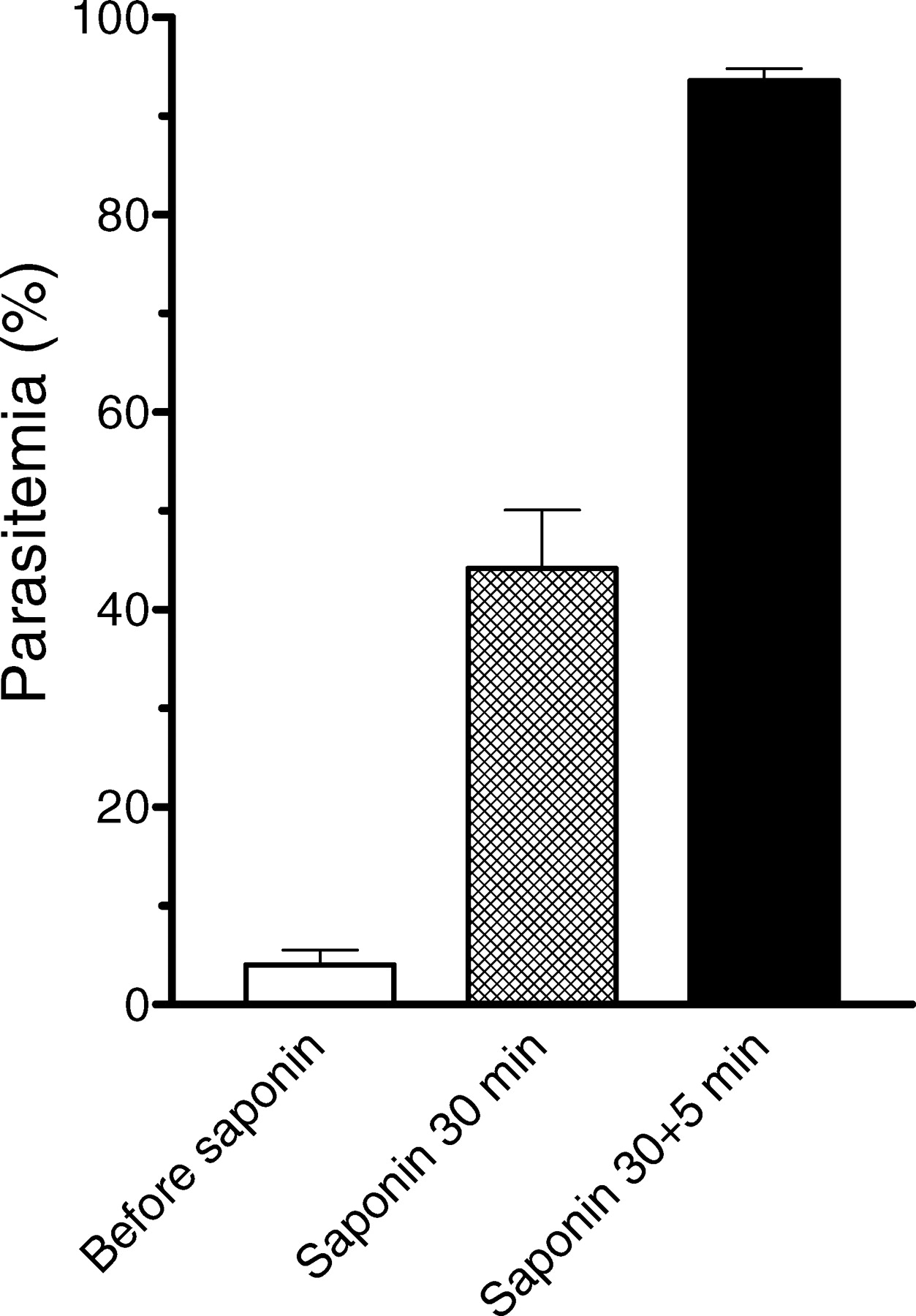
Enrichment of P. falciparum HB3 ring-infected erythrocytes by saponin hemolysis. Bars indicate mean ± SEM of 32 experiments, except in saponin for 30 min which contained 14 experiments.

Distribution of multiply infected erythrocytes. In a total of 13 experiments, cultures of P. falciparum HB3 strain were synchronized at the ring stage and enriched by two step exposures to saponin. Thin smears of the enriched preparations were stained with Giemsa stain and examined under the microscope. In the figure here only erythrocytes containing 2 or more parasites were counted.
Viability of saponin enriched P. falciparum HB3 strain. A synchronized culture was enriched at the ring stage to 99.5% parasitemia (left panel) and then cultured for additional 24 h to obtain trophozoites and schizonts (right panel). Some parasites that were apparently hemoglobin-free after the saponin treatment still matured to some extent during the 24 h incubation (arrows). A color version of this figure is available in the online journal.

Hemozoin production in normal (unenriched) and saponin enriched P. falciparum HB3 cultures. The parasitemia levels in the normal and enriched cultures were 2–5% and 97–99%, respectively. There parasitemia levels were determined after synchronization (normal culture) and saponin hemolysis (enriched culture). Bars indicate mean ± SEM of 6 experiments. The difference between the normal and enriched cultures was not significant at 0 h (P value was 0.7557), but was significant after 42 h of growth, suggesting that the parasites in the normal cultures produced more hemozoin than those in the enriched cultures (P value was 0.0372).

Effect of saponin on in vivo P. falciparum infected erythrocytes. Blood samples collected directly from patients were washed and subjected to saponin hemolysis. Most of the infected erythrocytes were hemolysed leaving “free” parasites in the supernatant (A) and a few intact ring-infected erythrocytes (B). The parasitemia of intracellular rings was 2.7% before and 3.3% after saponin hemolysis. A color version of this figure is available in the online journal.

Saponin enrichment of HbAS erythrocytes infected with P. falciparum rings. The experiments were done with P. falciparum PB112 strain. The initial culture (A) was synchronized with sorbitol to obtain B (4.5% parasitemia), which was enriched by saponin hemolysis to obtain C (82% parasitemia), which was grown in complete medium for 30 h to obtain D (79% parasitemia). A color version of this figure is available in the online journal.

Saponin enriched P. falciparum Amiri7 strain. Gametocytes (crescent shape) were also enriched. Parasitemia was 2.7% before and 63% after the enrichment. A color version of this figure is available in the online journal.
Footnotes
This work was supported by Kuwait University, Research Grant No. NM 01/02.
Acknowledgements
The technical contribution of Ms. Preethi Cherian is acknowledged. The scanning electron micrograph shown in this paper was kindly produced by the Electron Microscopy Unit, Faculty of Science, Kuwait University.