
Abstract
Orthopaedic tissues, such as bone, cartilage, intervertebral disc and tendon, contain cells that are difficult to culture and stimulate in vitro for repair of damaged tissue. Stem cells have the ability to self-renew and differentiate into many tissue types. Recent progress in stem cell research has led to an enthusiastic effort to utilize stem cells for orthopaedic tissue regeneration. Due to ease of harvest and abundance, adipose-derived mesenchymal cells (ASC) are an attractive, readily available adult stem cell that has become increasingly popular for use in many stem cell applications. Recent progress has been made in characterizing ASC and looking mechanistically at gene expression and cellular pathways involved in differentiation. This review focuses on (i) the characterization of ASC through expression of appropriate surface markers; (ii) modulation of in vitro differentiation of ASC through different scaffolds, growth factors, and media; and (iii) the use of ASC in orthopaedic tissue repair. Strategies for repair involve the use of differentiated or undifferentiated, fresh or passaged ASC, in conjunction with appropriate choice of media, growth factors and scaffolds. Recent in vivo studies utilizing ASC are discussed giving results on defect repair and potential for clinical orthopaedic tissue regeneration.
Introduction
Stem cells are characterized by the ability to differentiate into many lineage-specific cell types. Despite the pluripotency of embryonic fetal stem cells, ethical, regulatory and availability concerns have driven the search for adult multipotent mesenchymal stem cells (MSC) for use in tissue repair and regeneration.
For many years, bone marrow derived stem cells (BSC) were considered the major source of stem cells for tissue engineering applications. However, a recent study in mice showed that MSC reside in virtually all organs and tissues (1). Due to ease of harvest and abundance, adipose-derived mesenchymal cells (ASC) are an attractive, readily available adult stem cell that has become increasingly popular for use in many applications in orthopaedics (Table 1). ASC are readily harvested from adipose-tissue derived from subcutaneous surgery; stem cells are then isolated using enzymatic digestion, filtration and centrifugation of the stromal vascular fraction (SVF) that contains the stem cells along with non-adherent cells such as red blood cells. The SVF can be used directly as a source of stem cells or expanded in culture through many passages.
Although comparison between BSC and ASC is not a focus of this review, some examples are cited wherein ASC act in a way similar to the more extensively studied BSC. Both BSC and ASC derive from embryonic mesoderm and contain a stromal cell population that includes stem cells. Proliferation rates and similar gene expression pathways for BSC and ASC are found for osteogenic-, chondrogenic-, or lipogenic-like differentiation (2–4).
Here we discuss (i) current characterization of ASC through expression of appropriate surface markers; (ii) modulation of in vitro differentiation of ASC through different scaffolds, growth factors, and media; and (iii) the use of ASC in orthopaedic tissue repair.
Characterization of ASC
Adipose tissue is a highly complex mixture consisting of adipocytes, fibroblasts, vascular smooth muscle cells, endothelial cells, and immune cells as well as stem cells. Three criteria have been proposed to define MSC (5): a) adherence to plastic; b) multipotent differentiation potential to osteoblasts, adipocytes and chondroblasts; and c) specific surface antigen expression.
Surface antigen expression has been used extensively in immunology. Typical categories of surface marker proteins expressed in MSC are: adhesion molecules such as integrins (CD29, CD49e), receptor molecules such as hyaluronate (CD44), cadherins (CD144), surface enzymes (CD73), extracellular matrix proteins (CD90, CD105), intercellular adhesion molecules (CD54), vascular adhesion molecules (CD106), complement regulatory proteins, and histocompatibility antigens (6, 7). MSC should characteristically show positive localization of CD105, CD73 and CD90, but no localization of haematopoietic markers CD45, CD34, CD14 or CD11b, CD79α, CD19, or HLA-DR (5). However, CD90, CD34, CD 106, CD105, and Stro-1 are variably expressed in both human and animal ASC, especially in fresh or early passage cells (Table 2) (8–12). Comparison of initial fresh SVF and passaged and expanded ASC shows that the SVF contains stem cell markers CD166, CD44, CD29, CD73, CD90 and CD105. Expression of these markers increases significantly with increasing passage number, reaching 98% by P4 (passage 4). The SVF also expresses CD11, CD14, CD45 and CD34 (haematopoietic stem cell markers), which then decrease or are lost with increasing passage number, suggesting that adherence to plastic and subsequent expansion will select for a relatively homogeneous cell population compared with the SVF (11). Even with these differences in phenotype, tissue repair using freshly isolated SVF is possible (9).
It is important to note that no clear discrimination in surface markers between ASC, BSC and fibroblasts is currently available (13). ASC and BSC were shown to have very similar marker expression (14) (Table 2). Although some functional similarities between adult human fibroblasts and mesenchymal stem cells exist, such as characteristic surface markers and some differentiation ability (13), extensive characterization shows many functional differences (15, 16).
Characterization of marker expression for animal-derived MSC is challenging because commercial markers may not be available for a particular species, and not all human or murine antibodies cross-react with other species.
Effect of Media on ASC
Media used for tissue culture have an important impact on growth and differentiation of ASC. ASC are often plated and expanded in classical culture media containing balanced salt solutions such as minimal essential media (MEM), Dulbecco’s Modified Eagle’s Medium (DMEM), RPMI-1640 (developed at Roswell Park Memorial Institute), Basal Medium Eagle (BME), Dulbecco’s Modified Eagle’s Medium/Ham’s Nutrient Mixture F-12 (DMEM:F-12) with added serum (usually 10–20% fetal bovine serum). Serum concentrations can affect the proliferation of ASC. For example, increasing serum concentrations from 10% to 15% increases proliferation of MSC (17). This is not surprising since serum contains many cytokines and growth factors such as platelet derived growth factor (PDGF) and epidermal growth factor (EGF). Concerns have arisen about the possibilities of disease transmission through serum and supplements added to media leading to the creation of serum free media formulations. Formulations such as reduced serum media, containing nutrients to lower serum requirements; serum free media containing added proteins (18); protein free media; and chemically defined media comprising solely of defined low molecular weight constituents are now available. In order to test the effect of individual growth factors, a serum-free media has traditionally been used to test growth and differentiation of ASC (19). Transforming growth factor beta (TGFβ) and fibroblast growth factor-2 (FGF-2) was found to enhance growth but not differentiation of ASC. Epidermal growth factor (EGF) was shown to be an anti-differentiation factor. Enhanced proliferation of ASC was seen with a commercial growth medium when compared with DMEM. Only FGF-2 significantly promoted proliferation of ASC; the proliferative effect of FGF-2 alone was less than that seen with media only; thus it is possible that synergism among the other supplement factors may exist (17). Further work is needed to fully characterize these important factors which influence ASC growth and differentiation.
Specific osteogenic, chondrogenic or adipogenic media, containing various tailored differentiating supplements such as growth factors, are commercially available for differentiation of ASC into osteogenic, chondrogenic and adipogenic-like lineages respectively. These are recommended for characterization of the multipotent potential of various stem cell extractions using the specific media required. Considerable similarities between the differentiation potential of BSC and ASC were seen (20) (Figs. 1, 2). Non-differentiating commercial medium is also available to maintain an undifferentiated phenotype, or to enhance proliferation. Chondrogenic differentiation is usually accomplished by high density plating or micromass pellet formation (Fig. 1A, B) with TGFβ, dexamethasone and ascorbate supplemented media (21). The use of other growth factors such as BMP-6 has been investigated (22, 23). Osteogenic induction of ASC occurs after 3–4 weeks of tissue culture in an osteogenic media (Fig. 1C) containing dexamethasone, b-glycerophosphate, L-glutamine and ascorbate (24, 25).
Effect of Scaffolds on ASC
The field of tissue engineering involving ASC is rapidly advancing. Tissue engineering involves cell transplantation, materials science and engineering efforts directed toward development of biological replacement tissues. The source of donor cells may be a different species (heterologous), the same species but a different individual (allogenic), or the same individual (autologous).
Most mammalian cells are anchorage dependant and need an appropriate scaffold for gene-expression appropriate for the tissue of interest. Many different scaffolds exist, both as support for cells and to promote and maintain lineage differentiation. An ideal scaffold facilitates cell attachment, growth and differentiation, and ultimately, tissue formation and organization. Scaffolds are optimized for a permanent support for new cells and tissue, or may serve as a temporary resorbable cell delivery system.
Collagen is a readily available scaffold easily purified from tissue. Collagen implants are biodegradable through digestion by collagenase and the MMPs. The matrix of a collagen sponge slowly dissolves or is resorbed to allow the integration of the added cells and their matrix into the surrounding tissue. Scaffolds also allow for co-culture of two different cell types. The co-culture environment permits cell communication that may promote differentiation of the stem cell to the cell of interest, and exposure to secreted cell products. Varying scaffold density will affect the available size of the interstitial space where cells attach and proliferate. A 3-dimensional (3D) matrix with an open, porous structure (100–1000 μm) allows space both for cells to attach for nutrients and waste to permeate, and also room for extracellular matrix (ECM) accumulation (26). Collagenous microbeads have been used as a scaffold for binding ASC; bound cells were shown to proliferate and differentiate to an adipogenic or osteogenic phenotype (27). Scaffolds may also be coated with specific ECM components optimized for individual tissue types. Distinct ECM proteins (such as collagen types I or II) deposit from serum onto matrices and differentially mediate adhesion.
Other scaffolds, such as alginate, have been used as an injectable cell delivery vehicle. An injectable scaffold can form-fit to fill the desired shape, may be augmented with relevant growth factors, and does not require open surgery for placement (27). ASC mixed with an injectable scaffold, composed of biphasic calcium phosphate granules in suspension with a self setting hydrogel consisting of Si-hydroxy-methylcellulose (HIBS), and injected subcutaneously was able to produce woven bone in 4 weeks (28). Hydrogels that mimic native tissue such as cartilage in viscoelasticity and water retention have been widely used. The hydrogel polyethylene glycol (PEG) in combination with chondroitin sulphate has been injected with BSC for chondrogenic differentiation (29).
Polyesters of naturally occurring alpha-hydroxy acids, such as poly glycolic acid (PGA), polylactic acid (PLA), polyethylene glycol diacrylate (PEDGA), and poly(lactic-co-glycolic acid) (PLGA), are widely used in tissue engineering. These polymers can be coated with apatite to enhance MSC attachment and osteogenic differentiation (30). The degradation rate of these polymers can be varied from weeks to years by altering crystallinity, molecular weight and copolymer ratio. Although largely considered safe in humans, the ester bonds gradually hydrolyze over time potentially forming harmful acidic by-products which may damage tissue (31). Nondegradable scaffolds, such as fluoropolymers, overcome problems of degradation but are typically inert materials that do not favor cell attachment. ASC bound to fluoropolymers were shown to bind better to polymers with a larger pore size, although overall cell binding was low (32).
Current in-depth molecular knowledge of how ASC interact with various scaffolds is limited. Cell adhesion to the ECM is mediated by integrins that form a direct physical linkage between the ECM and the cytoskeleton. Knowing which integrins are commonly expressed by individual ASC allows predictions of preferred scaffolds. For example, type-I collagen receptors have been shown to act in adherence of BSC to PLGA scaffolds (33).
The Use of ASC in Orthopaedics
Chondrogenic Differentiation
In Vitro
The chondrogenic potential of human ASC grown in 3D culture has been evaluated by seeding into high density micromass (34, 35), agarose, alginate or gelatin 3D scaffolds (36–38), or by growth factor exposure. Culturing conditions commonly use TGFβ, dexamethasone and ascorbate supplemented media. FGF-2 has been shown to promote proliferation and chondrogenesis of ASC in micromass culture (39). Successful differentiation is marked by expression of typical gene and chondrogenic extracellular matrix proteins such as sox-9, collagen type II, aggrecan, biglycan, decorin, chondroitin sulphate, and cartilage oligomeric matrix protein.
TGFβ and the BMPs are involved in initiation and maintenance of the chondrogenic and osteogenic phenotype through induction of osteogenic or chondrogenic differentiation. This induction is mediated through receptor binding. TGFβ and bone morphogenic proteins (BMPs) belong to the TGFβ superfamily that signals through activation of the Smad proteins and major mitogen-activated protein kinase (MAPK) cascades. The final differentiation state is dependant on the BMP isoform, concentration of the BMP, and origin of the stem cells (40). BMP-2 induces chondrogenesis in micromass culture of ASC, as characterized by collagen II expression and GAG production (41). BMP2 induced both osteogenic and chondrogenic phenotypes when added separately to ASC cultured in alginate beads (42). However, when TGFβ and BMP-2 were combined, the osteogenic phenotype was inhibited. A possible explanation for this cell behavior is that BMP2 induces the transcription factor Nkx3.2 which downregulates the osteogenesis gene Runx2; MSCs then undergo chondrogenic, not osteogenic, differentiation (42).
BMP-7 and BMP-6 induced a chondrogenic phenotype with ASC in micromass or alginate bead culture respectively (23, 40), while BMP-2 induced an osteogenic phenotype (40). Another study showed that BMP-6 induced TGFβ-receptor-1 expression in micromass cultures, and combined application of TGFβ and BMP-6 increased the chondrogenic potential of ASC (22). Material properties of a micromass culture system using ASC showed dynamic and static elastic modulus results similar to those of chondrocytes in an early chondrogenic differentiation process (43).
Several investigators have compared chondrogenesis efficacy between ASC and BSC (41). It has been suggested that ASC may have a lower chondrogenic differentiation potential than BSC (35), or that there may be little differences (44). Other studies show advantages of ASC. For example, compared with bone marrow aspiration, ASC are easy to expand in culture, and a large number of cells are available from a small amount of readily obtained tissue (45). Furthermore, ASC show a low senescence rate even when harvested from an adult, and do not cause an immune rejection response (46, 47).
In Vivo
Use of ASC in the healing of cartilage defects has recently been investigated. ASC were grown in high density culture in a growth medium containing FGF-2, growth hormone and TGFβ and placed in a fibrin glue scaffold inserted into the defects. Histologic analysis supported formation of a hyaline cartilage phenotype (45).
Osteogenic Differentiation
In Vitro
Osteogenic induction of ASC in culture occurs after 3–4 weeks in an osteogenic media containing dexamethasone, b-glycerophosphate, L-glutamine and ascorbate (16). Induction produces calcium phosphate mineralized extracellular matrix, osteocalcin, alkaline phosphatase, osteopontin, collagen type I, and expression of osteogenic transcription factor genes such as Runx2 and OSX (Osterix) (24, 25, 48). Stains such as alizarin red and von Kossa will indicate positive production of mineralized matrix in vitro (Fig. 1C). Induction of osteogenesis by MSC is enhanced by adding FGF2 to the culture media before exposure to osteogenic or chondrogenic inducing conditions (49). Once stem cells become differentiated, osteoblasts produce the extracellular matrix proteins indicative of the osteoblast phenotype.
Osteogenesis can be stimulated by members of the TGFβ superfamily, such as the bone morphogenic proteins BMPs. Transcriptional mechanisms of osteogenesis involve Runx2 which acts as a central mediator executing signals from the BMP and Wnt pathways to promote phenotype commitment and osteogenesis (50). Binding of BMP-2, −4, and −6, results in the activation of R-Smad with cell-specific transcription factors such as Runx2 with the formation of Runx2-Smad complexes that regulate target genes (42). Runx2 is a DNA-binding protein that acts as a scaffold for regulatory factors involved in skeletal gene expression essential for osteoblastic differentiation and skeletal morphogenesis.
In Vivo
Various ASC bone tissue engineering strategies have been reported. Initial studies involved subcutaneous in vivo formation of osteoid or bone tissue from ASC. Cells are typically seeded into scaffolds with or without prior in vitro osteoinduction, implanted subcutaneously for several weeks, followed by characterization of cell differentiation and bone matrix formation. In an early study using hydroxyapatite/tricalcium phosphate scaffolds, osteoid formation was present in 80% of SCID mice subcutaneous implants loaded with ASC, but absent in cell-free implants (51). In similar studies, both BSC and ASC loaded into atelocollagen scaffolds (52), and a tricalcium phosphate scaffold (53), were shown to differentiate into osteoblasts in vivo. Osteoinduced ASC mixed with the injectable scaffold induced the formation of vascularized and mineralized woven bone with osteoblasts, osteocytes and osteoclasts (28).
Recent work has shown the future utility of ASC in spine fusion. Recently successful spine fusion took place when ASC transduced with a BMP-2 expressing vector and loaded into a collagen type-I scaffold was implanted in rats. In contrast, when ASC was pre-treated with osteoinductive media containing BMP-2, spinal fusion was minimal (54).
In more complex animal models involving repair of bone defects, early work shows great promise in the repair of critical-sized defects where enough bone is lost to preclude normal biological healing. Some debate exists over whether in vitro osteoinduction of ASC is necessary prior to in vivo implantation of the ASC. In vitro stimulation is time consuming and expensive, while on the other hand unstimulated ASC placed directly in vivo may exhibit a slower or absent osteoinductive response.
Osteoinduced ASC gave favorable results when compared with uninduced ASC in several studies. Osteoinduced ASC loaded onto gelatin foam scaffolds exhibited greater rabbit calvarial defect healing than did non-induced ASC (55). Femoral bone defects were healed by ASC genetically modified to overexpress BMP-2 and loaded onto a collagen-ceramic scaffold. The defects were also healed by ASC loaded onto a scaffold impregnated with rhBMP-2; uninduced ASC, however, did not heal the defects (47). Similarly, ASC genetically modified by BMP-2 produced a significant increase of newly formed bone in a canine ulna bone defect using porous tricalcium phosphate ceramic granules as scaffold (56). Osteoinduced ASC layered over PLGA were implanted into rat calvarial defects. More robust bone tissue was obtained from the osteoinduced ASC than in controls (57). However, uninduced ASC or BSC grown on an apatite-coated PLGA scaffold were also effective in repair of critical-sized mouse calvarial defects. Significant intramembranous bone formation was present by two weeks (30). A “best of both worlds” approach involving rapid in vitro osteoinduction prior to in vivo implantation of ASC has been suggested (58). Recently, ASC mixed with platelet-rich plasma has been shown to induce periodontal ligament-like and alveolar bone regeneration in rats (59).
Intervertebrate Disc Differentiation
In Vitro.
Repair of degenerated intervertebrate disc (IVD) using MSC is of great interest due to the high incidence of low back pain. The IVD phenotype is not as well characterized as either cartilage or bone, and consequently less information exists about differentiation into the disc phenotype. Potential exists for harvested autologous ASC, either with or without growth factor pre-stimulation or matrix attachment, injection into IVD tissue for repair (60). When ASC and nucleus pulposus (NP) cells were grown in micromass under co-culture conditions where the micromasses were separated by a permeable membrane allowing soluble factor diffusion, aggrecan and collagen II expression were upregulated (61) Culture of ASC in type II collagen hydrogels, co-cultured with micromass nucleus pulposus (NP) cells, caused upregulation of collagen type II and aggrecan gene expression. This suggests that soluble factors released by NP cells could direct ASC to an NP-type lineage (62). The feasibility of a stem cell therapy for disc repair was suggested by a recent parallel in vitro in vivo study. The in vitro study showing increased ECM production after BSC and disc cells were co-cultured, while in vivo data showed persistence of BSC in rabbit disc for at least 24 weeks after implantation (63).
In Vivo
While no papers using ASC show disc repair, several recent papers using BSC report exciting findings. When BSC combined with a hyaluronic acid-derived scaffold were injected into pig IVD, discs had a central NP-like region similar to the normal structure (64). BSC, transplanted to degenerative discs in rabbits, proliferated and differentiated into cells expressing some of the major extracellular components of disc (65).
Tendons and Ligaments
ASC use in tendon and ligament repair is limited to date. Treatment of acute collagenase-induced tendon lesions in horses with autologous SF resulted in improved tendon morphology. Increased gene expression of cartilage oligomeric matrix protein (COMP) was seen. COMP binds to multiple collagen fibrils, possibly improving collagen fibril formation and organization (66).
Plasmids containing BMP-8 and an active variant of Smad8 have been transfected into a mesenchymal stem cell line and used to repair a rat tendon defect; the resulting tissue showed orientated collagen bundles (67). BSC, both with and without transfection with platelet derived growth factor, enhanced cellular infiltration and collagen deposition into an anterior cruciate ligament repair in rabbits (68).
Currently, no papers using ASC involve ligament repair. BSC was used to fabricate anterior cruciate ligament constructs in a collagen type I hydrogel attached to bone cylinders. After 10 days the construct was exposed to cyclic stretching. Elongated fibroblastic cells were produced along with collagen types I and III, fibronectin and elastin (69). These, and other MSC studies, suggest the potential of ASC in tendon and ligament repair.
Conclusions
Enormous advances in stem cell biology are anticipated to yield great benefit for the future treatment of orthopaedic tissue repair. Although a relative newcomer to the field, ASC have proven to be a useful source of stem cells in bone and cartilage repair. Exciting potential exists for IVD, tendon and ligament repair achieved by use of ASC. Additional research is needed to find cell markers specific for ASC characterization, and also for identification of markers that characterize each stage of differentiation. Optimization of cell growth, in vitro differentiation and appropriate choice of scaffolds will offer reproducible and successful surgical outcomes for orthopaedic tissue repair.
Use of ASC for Orthopaedic Tissue Repair a
CD Marker List of ASC, BSC Mouse ASC or BSC and SVF a
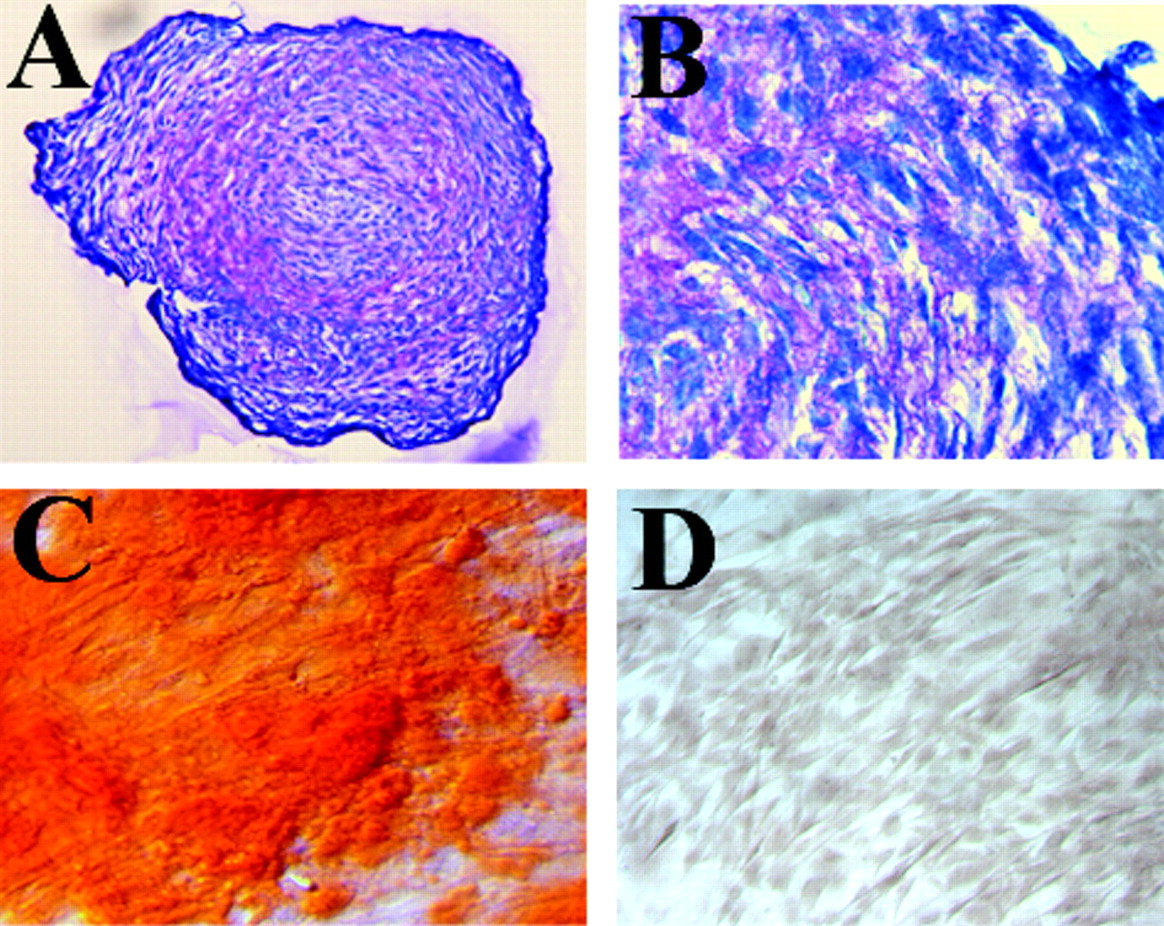
Osteogenic and chondrogenic differentiation of human ASC. A and B: High density cultures showed formation of a chondrogenic phenotype when cultured in micromass; pink ECM staining marks proteoglycans (A, ×95; B, ×240). C: ASC treated with osteogenic media for three weeks stained with Alizarin red. Red staining marks mineralized matrix produced by osteoblasts. D: Monolayer ASC in culture with non-differentiating media show no bone formation (C and D, ×105).

Lipogenic, chondrogenic and osteogenic differentiation of human ASC and BSC. Histochemical staining of adipocytes (oil red O), chondrocytes (Alcian Blue), and osteocytes (Alizarin red). BSCs and ASCs were induced into adipogenesis for 14 days, chondrogenesis under pellet culture for 21 days, and osteogenesis for 14 days. Reproduced by permission from
Footnotes
This work was supported by the Brook Center for Back Pain Research and the Charlotte-Mecklenburg Education and Research Foundation.