
Abstract
Familial Mediterranean fever (FMF) is an autoinflammatory disease caused by mutations in the MEFV locus, which encodes the protein pyrin. While it is known that pyrin is expressed in myeloid cells and several fibroblastic cell types, the exact function of pyrin in these cells and the mechanism underlying the pathological effect of pyrin mutations have yet to be revealed. Here, we document that in migrating human monocytes, pyrin protein is dramatically polarized at the leading edge, where it co-localizes with polymerizing actin. ASC (Apoptosis-associated Speck protein with CARD domain), a known pyrin-interacting protein and a critical component of the inflamma-some, is also located at the leading edge in migrating monocytes. Similarly, both pyrin and ASC concentrate in dynamically polymerizing actin-rich tails generated by Listeria monocytogenes. Pyrin’s B-box and coiled-coil region is required for its association with Listeria tails. Pyrin also binds, with low affinity and via the same domains, to actin, VASP, and Arp3. Though disease-causing mutations in pyrin do not appear to alter its localization to the leading edge or to Listeria rocket tails, they could potentially have important functional consequences in the context of processes such as migration and cell synapse formation. The co-localization of pyrin and ASC together at such sites may provide an important link between cytoskeletal signaling and inflammasome function.
Introduction
The term “autoinflammatory” is used to describe a set of heritable and multi-factorial human diseases characterized by fever and inflammation without apparent T- or B-cell involvement (1). These diseases are believed to represent disorders of the innate immune system. FMF, caused by mutations in the MEFV locus, is characterized by sporadic attacks of fever with painful inflammation that is most often localized to synovial, pleural or peritoneal membranes. FMF is the most common of these auto-inflammatory syndromes, and is particularly prevalent in the Mediterranean region. In fact, the frequency of several of the known MEFV mutant alleles is so remarkably high in regions of Armenia, Turkey and Morocco that a possible heterozygote advantage has been postulated (2, 3). Indeed, positive selection appears to have acted on the pyrin protein during primate evolution (4). Interestingly, the known human mutations in the pyrin protein are predicted to be wild-type sequences in primate ancestors. These amino acid sites must therefore be sites that can change the functional character of the pyrin protein, without destroying it, in response to some unknown environmental pressure. However, nature of the functional changes caused by the mutations is not yet clear.
FMF attacks are characterized by a massive influx of neutrophils into the inflamed site. Pyrin, the protein product of the MEFV gene, is expressed in neutrophils as well as in monocytes/macrophages and dendritic cells (5, 6). Pyrin is also expressed, although at lower levels, in tissue fibroblasts found at the sites of inflammatory attacks, including skin, synovial and peritoneal fibroblasts (5–7). These localizations, combined with the evolutionary data indicating that pyrin is functionally evolving in response to some environmental pressure, suggest that pyrin may operate on the front line of the inflammatory response, either sensing or coordinating responses to pathogenic insults.
Recent work in several laboratories has documented the interaction of pyrin with five cellular proteins: PSTPIP1 (8), 14-3-3 (9), Caspase-1 (10), ASC (11), and Siva (12). The interaction with ASC (Apoptosis-associated Speck-like protein with CARD domain), is particularly interesting since multimers of ASC provide a central platform for the formation of several types of inflammasome, a multiprotein complex that efficiently activates Caspase-1, promoting the activation and release of IL-1β (13, 14). ASC also has a tendency to form large peri-nuclear aggregates called “specks,” structures that appear to presage the death of the cell (11, 15). Pyrin binds to ASC via N-terminal pyrin domains (PyD), encoded by exon 1 of both proteins. The interaction of pyrin with ASC modulates both the formation of ASC specks and the rate of apoptotic death following speck formation (11). Additional data suggest that pyrin also modulates ASC’s inflammasome function (16–18). Indeed, interactions between ASC and Caspase-1 (19), between pyrin and Caspase-1 (10) and between ASC and pyrin (11) have all been documented, and analysis of the binding domains responsible for these interactions indicate that tri-molecular interactions are possible. That is, Caspase-1 binds via its N-terminal CARD domain to the C-terminal CARD domain of ASC; pyrin binds to the N-terminal PyD of ASC; and the C-terminal rfp/B30.2/SPRY domain of pyrin interacts with Caspase-1. Thus, this complex of proteins could play a regulatory role in inflammation and perhaps apoptosis.
Pyrin also interacts with PSTPIP1, the protein that is mutated in PAPA syndrome (Pyogenic Arthritis, Pyoderma gangrenosum, and Acne), another debilitating autoinflam-matory disease. The direct link between pyrin and PSTPIP1 involves pyrin’s B-box and coiled coil domains; the B-box alone is necessary, but not sufficient for this interaction (8). PSTPIP1 is homologous to cdc15, an actin-associated protein important for cytokinesis in yeast. Overexpression of PSTPIP1 results in the generation of filopodia by 3T3 cells (20), suggesting that mammalian PSTPIP1 also affects actin organization.
Earlier studies of pyrin in transfected cells documented direct interactions between pyrin and elements of the cellular cytoskeleton (21). An interaction with tubulin was mapped to the N-terminal PyD of pyrin. A potential interaction with actin was also documented in that study, but not well characterized; in transfected cells, pyrin was observed to co-localize with actin in lamellar structures and in supranuclear ruffles, but not with stress fibers. The domain of pyrin responsible for this co-localization could not be determined (21). More recent work indicates that, through its interaction with Siva, a protein that functions in the apoptotic response to oxidative stress, pyrin is occasionally localized in actin stress fibers (12). These studies, and the interaction of pyrin with PSTPIP1, predict that pyrin plays a role in cytoskeletal signaling pathways. This is important because such pathways are regulators of cell migration as well as cell-cell signaling, critical processes that control the inflammatory response.
Here, we examine more closely the distribution of pyrin in native human monocytes. We document the co-localization of pyrin and ASC in membrane ruffles and show that, in migrating human monocytes, both ASC and pyrin are dramatically polarized to the leading edge, a site of active actin polymerization. We confirmed the association of both pyrin and ASC to polymerizing actin and further mapped the pyrin domains involved using the Listeria monocytogenes rocket tail assay (22). In addition, we identified low affinity interactions between pyrin and three cytoskeletal proteins (Arp3, VASP, and actin) that may be involved in the recruitment pyrin to sites of active actin polymerization in the cell. The additional co-localization of ASC to the same dynamic actin-rich regions could provide a previously unrecognized means to connect cytoskeletal function to inflammatory signaling.
Materials and Methods
Cell Culture and Transfection.
COS-7, HeLa, and 293T cells were cultured in Dulbecco’s Modified Eagle Medium (Gibco by Invitrogen, Carlsbad, CA) supplemented with 10% (vol/vol) fetal bovine serum. For experiments, cells were plated on sterile glass coverslips in 6-well culture plates. Cells were transfected using FUGENE-6 (Roche Applied Science, Indianapolis, IN) and incubated for 24–48 hrs before analysis.
Monocyte Isolation.
Human blood was collected from healthy volunteers (IRB# 1992-0480) and mixed with an anti-coagulation solution containing 0.14 M anhydrous citric acid, 0.20 M citric acid trisodium salt, and 0.22 M dextrose. Blood was then incubated at room temperature for 30–45 mins after the addition of a solution containing 6% dextran to facilitate sedimentation of red blood cells. The upper, leukocyte-rich layer was then removed and any remaining red blood cells were destroyed by hypotonic lysis in distilled water. Leukocytes were concentrated by centrifugation, then layered onto Ficoll/Paque (Pharmacia, Uppsala, Sweden) density gradient and centrifuged again. The monocyte/lymphocyte layer was removed, and monocytes were enriched by cell adhesion. Non-adherent lymphocytes were washed off 24 hrs later, and the adherent monocytes were fixed and stained.
Plasmids and Antibodies.
All myc- and FLAG-tagged constructs were cloned into pCMVTag3a and pCMVTag2b (Stratagene, La Jolla, CA) respectively. GFP tagged gelsolin, capping protein, Arp3, VASP, and α-actinin as well as YFP tagged actin constructs were obtained from Maria Diakonova, Ph.D. (University of Michigan, Ann Arbor, MI). HRP-conjugated α-myc primary antibody and goat α-mouse IgG secondary antibody were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Mouse α-FLAG (M2 monoclonal), rabbit α-myc (polyclonal), and mouse α-tubulin (monoclonal) were obtained from Sigma (St. Louis, MO). Mouse α-GFP (monoclonal) was acquired from Clontech (Mountain View, CA). Goat α-mouse AF488, goat α-mouse AF568, goat α-rabbit AF488, and goat α-rabbit AF568 secondary antibodies as well as Phalloidin AF488 were purchased from Molecular Probes (by Invitrogen, Eugene, OR). Rabbit α-human pyrin polyclonal antibody was generated against pyrin amino acids 1–374, and was generously provided by Dr. Daniel Kastner (National Institutes of Health, Bethesda, MD). Mouse α-ASC monoclonal antibody was a kind gift of Dr. Junya Masumoto (Shinshu University School of Medicine, Nagano, Japan).
Listeria Rocket Assay.
Single colonies of wild-type Listeria monocytogenes were picked from BHI agar plates and grown in BHI media at 25°C overnight. The bacteria were then sub-cultured in fresh BHI media at 37°C for 90 mins. Bacterial cells were pelleted by centrifugation, washed in PBS three times, and then diluted to an OD of 0.05 in PBS. One mL of this bacterial solution was then added to each well containing COS-7 cells, resulting in approximately 1 × 106 bacteria per well. The bacteria were allowed to infect cells for 2 hrs, then the media was replaced with DMEM + 10% FBS containing gentamicin at 50 μg/ mL. Cells were incubated for an additional 5 hrs, and then fixed with 4% paraformaldehyde in PBS and stained.
Cytochalasin D Treatment.
A 2 mM stock of cytochalasin D prepared in chloroform was diluted in cell culture medium to a working concentration of 10 μM. Cells were treated for 30 mins before fixation with 4% paraformaldehyde in PBS.
Immunofluorescence.
Cells were permeabilized using 0.2% Triton X-100 in PBS, and blocked with a solution containing 10% goat serum, 1% bovine serum albumin, and 0.1% Tween 20 in PBS. Primary and secondary antibody application was carried out, and nuclei were counterstained with DAPI. Coverslips were mounted on slides using a ProLong Antifade Kit (Molecular Probes by Invitrogen, Eugene, OR) and allowed to dry in the dark for several hours. Slides were visualized using a Nikon E800 microscope or an Olympus FV-500 confocal microscope.
Immunoprecipitation and Western Blotting.
293T cells were transfected as described, and were lysed 48 hrs later using a buffer containing 0.5% Triton-X. Protein lysates were tested for expression and proteins were precipitated using α-myc antibody and a mixture of protein A and protein G-coated sepharose beads (Invitrogen, Carlsbad, CA). The immunoprecipitation reactions were run out on gradient polyacrylamide gels (4–20%; purchased from BioRad Laboratories, Hercules, CA) and transferred to nitrocellulose membranes. Membranes were blocked in a buffered solution of nonfat milk and bovine serum albumin, and then treated with antibodies diluted in blocking solution. Membranes were washed extensively and treated with SuperSignal West Pico Chemiluminescent Substrate (Pierce Biotechnology, Rockford, IL), then exposed to X-ray film.
Results
Pyrin and ASC Colocalize with Actin in Membrane Ruffles in Human Monocytes and in Transfected Cells.
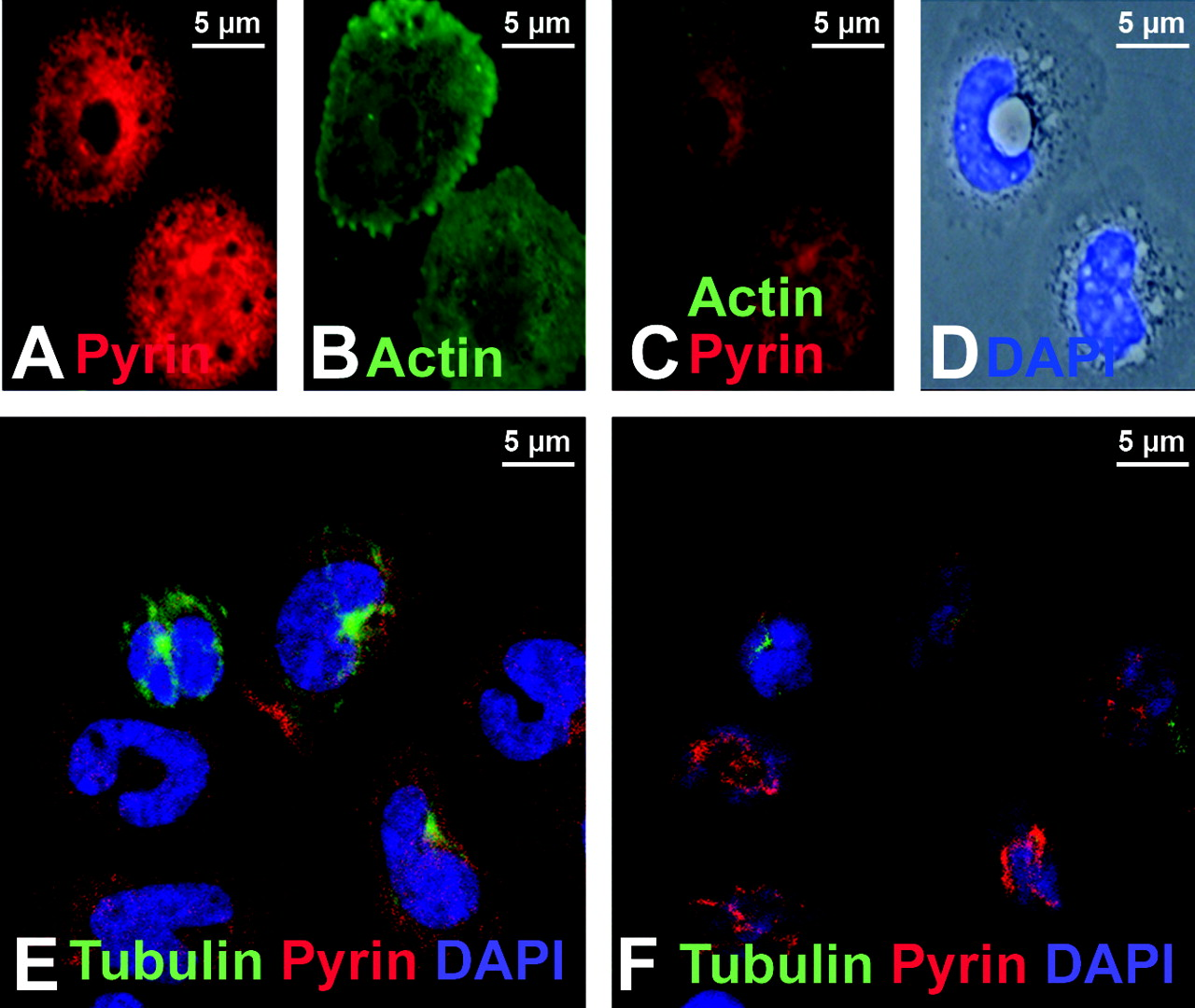
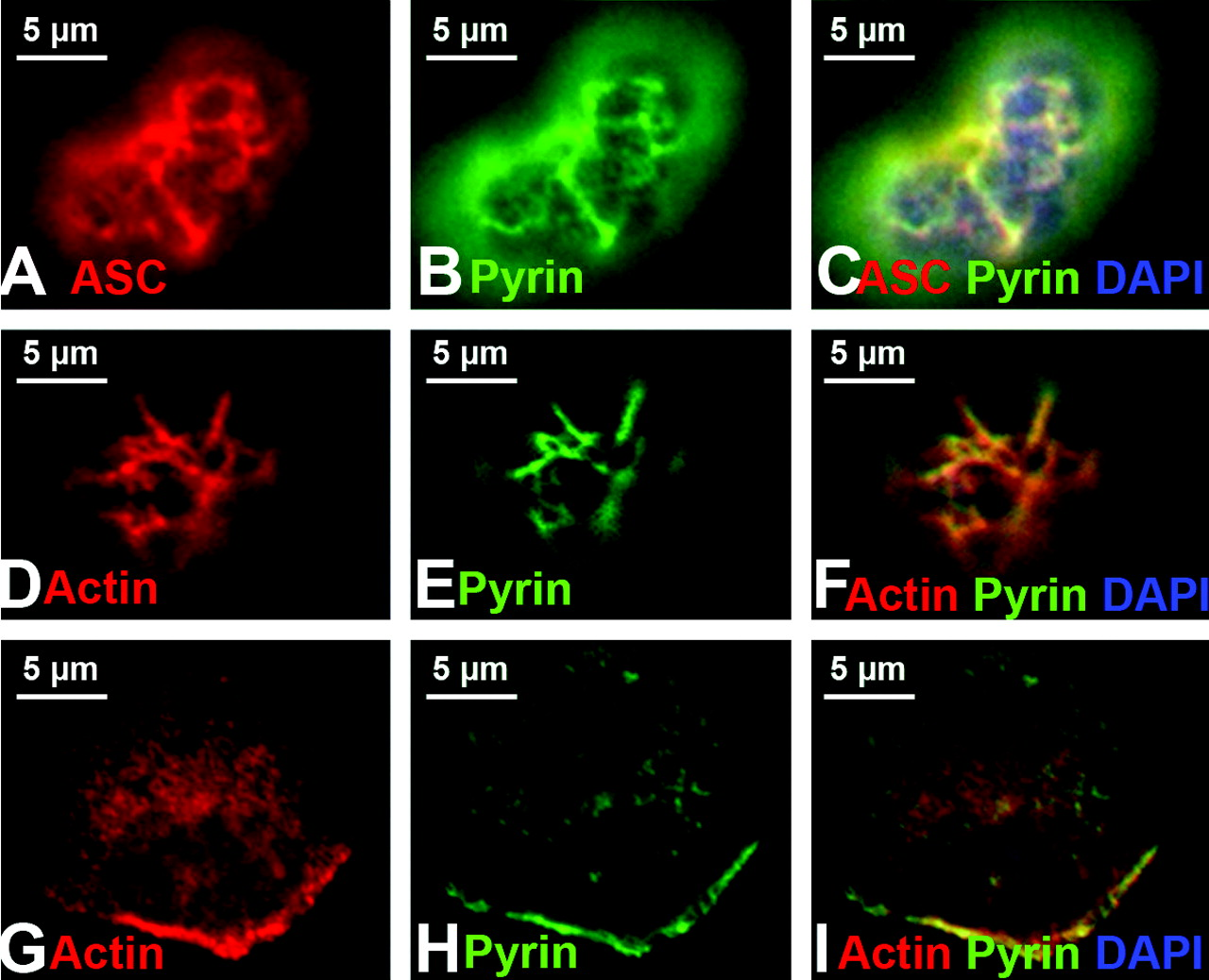
Freshly isolated human monocytes were examined after immunostaining with a polyclonal antibody previously documented to detect human pyrin (6). As shown in Figure 1A–C, pyrin was found in filamentous structures at the top of the cell, above the nucleus. The same cells were also stained with a monoclonal antibody that recognizes ASC and an identical pattern was observed (Fig. 1D–F). In cells co-stained for pyrin and filamentous actin (using phalloidin), the filamentous structures were revealed to be actin-rich membrane ruffles (Fig. 1G–I).
Transfection of HeLa cells with myc-tagged pyrin confirmed the localization of pyrin to supranuclear ruffles (Fig. 1J, arrow); pyrin was also observed in lamellapodial structures (Fig. 1K, arrow). Co-staining with phalloidin confirmed the presence of actin in both ruffles and lamellapodia (Fig. 1L). At higher magnification (Fig. 1M–O), pyrin could be found in lamellapodia (arrowheads), but not in actin stress fibers (arrow).
The Pattern of Pyrin Staining in Human Monocytes Requires Intact Actin Microfilaments, but Does Not Coincide with the Microtubular Network.
To test whether the pattern of pyrin localization in human monocytes depends on an intact actin network, we treated monocytes with cytochalasin D, which disrupts actin filaments. The reticular pattern of both pyrin and phalloidin staining was abolished in cytochalasin D-treated cells (Fig. 2A–D).
Pyrin was previously shown to co-localize with micro-tubules, but those findings were based only on the examination of pyrin in transfected cells (21). We examined the relationship between pyrin and the microtubule network in human monocytes using confocal microscopy. Figure 2E and 2F show two different Z-planes of a field of human monocytes. In Figure 2E, a section near the base of the cells reveals the microtubule organizing network (green) in several cells. A section close to the top of the cells (Fig. 2F) shows the supranuclear distribution of pyrin. In no case did we find evidence of co-localization between pyrin and the microtubular network in human monocytes.
Pyrin and ASC Concentrate in Areas of Actively Polymerizing Actin.
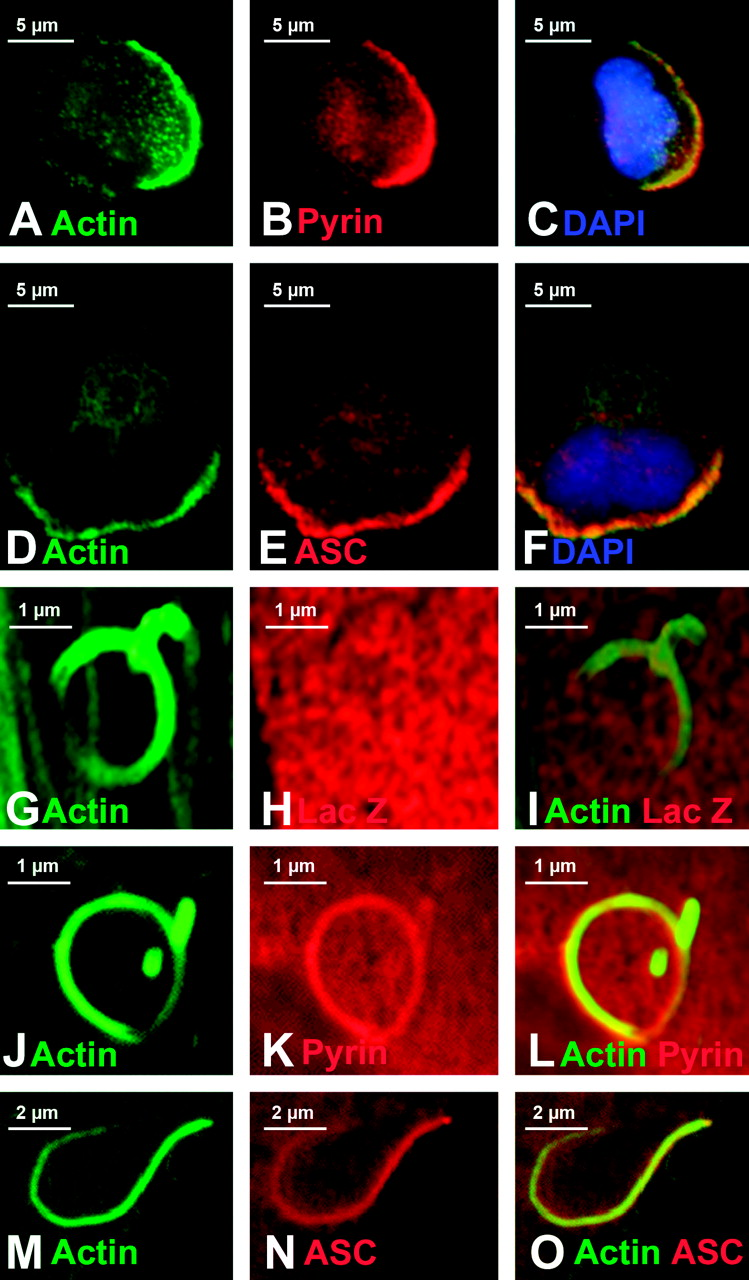
In some monocytes stained with both phalloidin and anti-pyrin, we noted a clear-cut polarization of actin staining (Fig. 3A). These cells exhibited the typical morphology of migrating monocytes (23): the leading edge was actin-rich, the nucleus was pulled towards the leading edge and the trailing edge contained numerous spiky cell projections (the uropod). Strikingly, in these migrating cells, pyrin staining was polarized to the actin-rich leading edge (Fig. 3B–C). In cells stained for actin and ASC, a similar polarized pattern of staining was observed (Fig. 3D–F).
The leading edge of migrating cells is a region that is rich in polymerizing actin. A convenient assay to detect and further dissect the association of proteins with the actin polymerizing machinery is the Listeria monocytogenes actin rocket tail assay (22, 24). Listeria monocytogenes is an infectious bacterium that uses the actin polymerizing machinery of the host cell to build a comet-shaped actin-based tail (a “rocket”) that propels the bacterium throughout the cytoplasm and into neighboring cells. These tails are distinct structures, easily identified by phalloidin staining (Fig. 3G).
We transfected myc-tagged pyrin into COS-7 cells and infected these cells with Listeria. COS-7 cells were chosen because they are large, flat, and easily transfectable, making for easy viewing of rocket tails. Additionally, COS-7 cells do not express pyrin or several known pyrin-binding proteins, allowing us to differentiate between a direct interaction of actin-associated proteins with pyrin and an indirect association of pyrin with actin-associated proteins. Rocket tails were allowed to develop, and then were visualized by staining with phalloidin. The negative control protein LacZ was not recruited to Listeria rocket tails (Fig. 3G–I). However, pyrin was consistently found in rocket tails (Fig. 3J–L); the pyrin staining intensity inside of the actin tails exceeded the staining intensity elsewhere in the cell, indicating that pyrin was being concentrated within the tail, in close association with the polymerizing actin. Likewise, ASC was drawn to Listeria actin tails (Fig. 3M–O).
The Association of Pyrin with Sites of Active Actin Polymerization Requires the B-Box and Coiled-Coil Domains.
To map the regions of pyrin that mediate its recruitment to Listeria tails, we used a series of deletion mutations, summarized in Figure 4 (see Supplemental Fig. 1). Exon 1, the region encoding the PyD of pyrin that is known to interact with ASC, was dispensable for pyrin localization to Listeria tails (Supplemental Fig. 1A–C), as was exon 2 (Supplemental Fig. 1D–F). However, exon 3 (Supplemental Fig. 1G–I) and exons 3–4 (data not shown) were weakly recruited. Importantly, the C-terminal Rfp/B30.2/SPRY domain encoded by exon 10, where the majority of pyrin mutations are found, was not required for recruitment of pyrin to Listeria rocket tails (Supplemental Fig. 1J–L). Two minimal regions of pyrin were found to strongly and consistently localize to Listeria tails: exons 3–5, encoding the B-box and part of the coiled-coil region of pyrin (Supplemental Fig. 1M–O), and the exon 6–10 region, which includes additional coiled-coil sequences as well as the Rfp domain (Supplemental Fig. 1P–R). Testing the exon 6–9 region alone was not feasible due to the apparent instability of this protein fragment. In addition, testing the PyD and CARD domains of ASC was impossible due to the fact that these domains, when expressed alone, form filaments in cells (11, 25).
Exclusion of the Known Pyrin-Interacting Protein PSTPIP1 as the Mediator of Pyrin’s Recruitment to Listeria Rocket Tails.
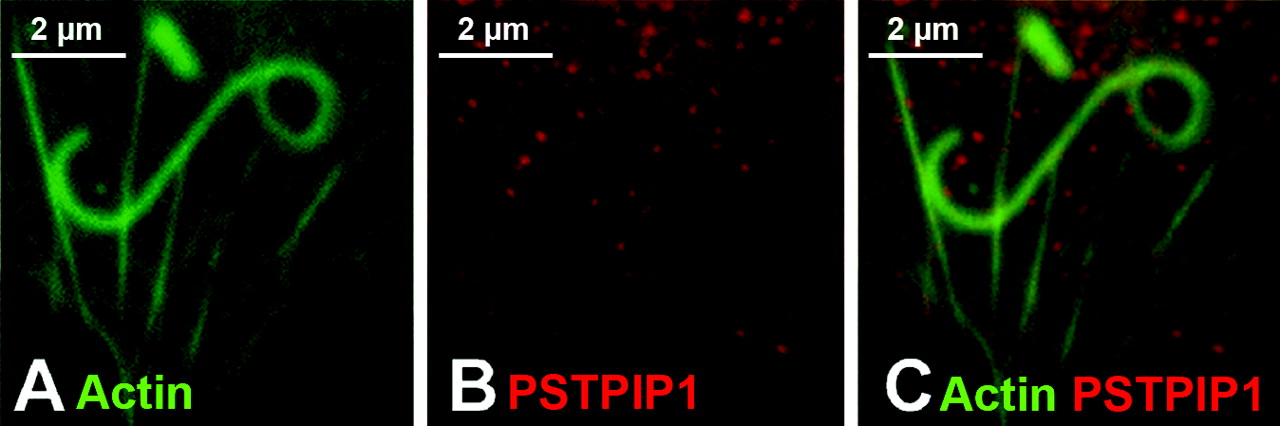
PSTPIP1 was previously isolated from a monocyte cDNA library using a yeast two hybrid screen with pyrin as the bait (8). PSTPIP1 is the homolog of yeast cdc15, a yeast actin-binding protein (26); overexpression of murine PSTPIP1 in 3T3 fibroblasts causes actin reorganization and extension of filopodia (20). More recent work has shown that PSTPIP1 is recruited to the immunological synapse by WASP, a protein that is important for the initiation of actin polymerization (27). Thus, PSTPIP1 is an actin-associated protein that has the potential to modulate PEST phosphatase activity in actin-based cell functions. We hypothesized that since pyrin is known to interact with PSTPIP1, and since PSTPIP1 and pyrin are both found in monocytes, PSTPIP1 may recruit pyrin to the leading edge. We therefore tested this possibility in the Listeria rocket assay. We found that when PSTPIP1 is transfected into COS-7 cells, PSTPIP1 forms delicate filaments throughout the cytoplasm (data not shown). When these cells are further infected with Listeria, the filaments are often disrupted, but PSTPIP1 does not localize to Listeria actin tails in the absence of pyrin (Fig. 5). Thus pyrin’s recruitment to sites of active actin polymerization likely does not involve PSTPIP1.
Identification of Cellular Proteins that Could Recruit Pyrin to Listeria Rocket Tails.
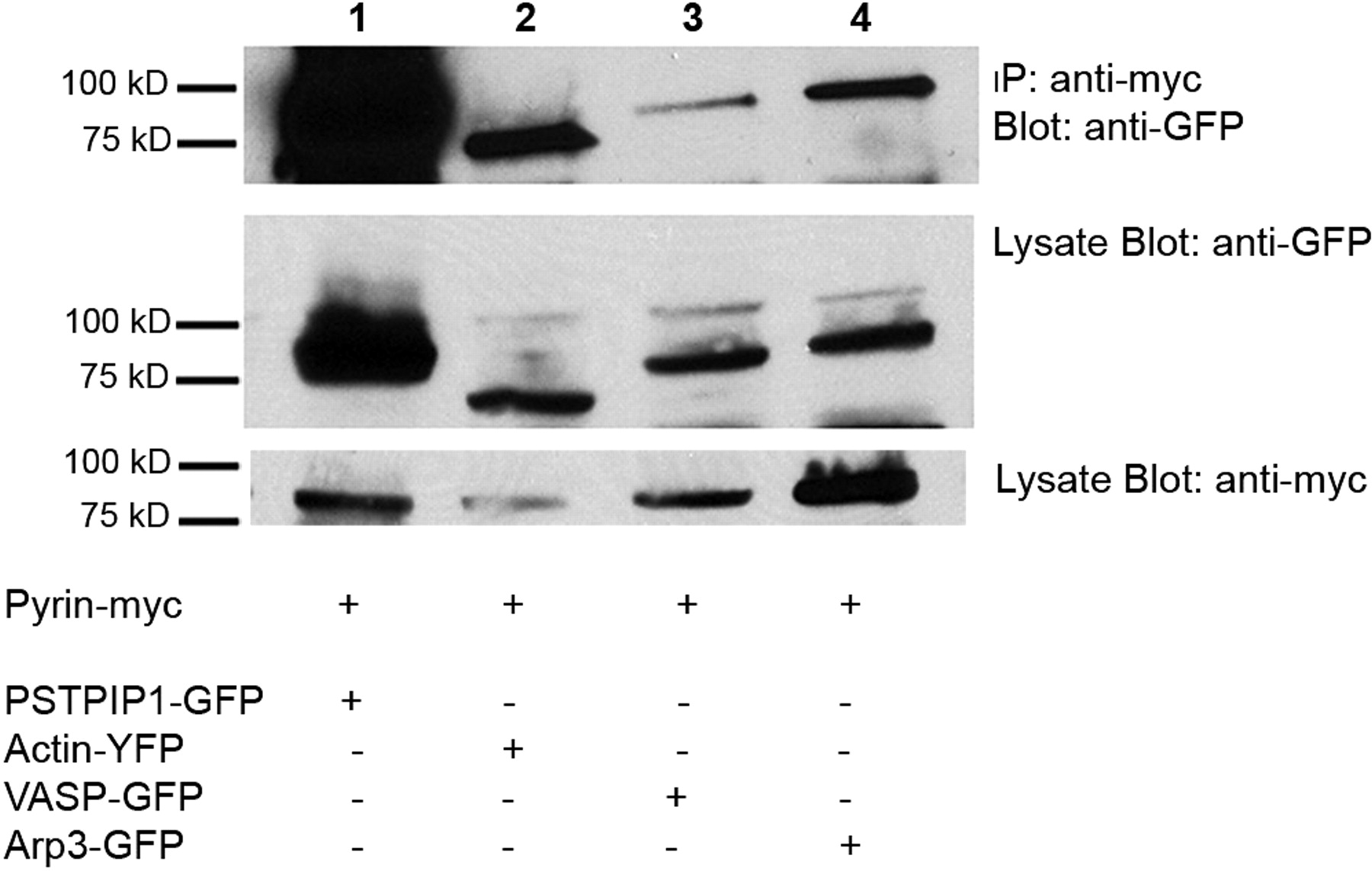
Seven cellular proteins are minimally required for the formation of Listeria rocket tails in vitro: VASP, gelsolin, CapZ, α-actinin, Arp3, profilin, and actin (reviewed in ref (28)). We tested, using immunoprecipitation assays, whether GFP-labeled forms of each of these proteins could bind to exons 3–5 or exons 6–10 of pyrin (regions sufficient for recruitment to rocket tails). Our initial experiments showed a co-precipitation of pyrin exons 3–5 with actin, VASP, and Arp3, while only actin and VASP were precipitated with exons 6–10 (data not shown). These interactions were rather weak, as immunoprecipitation was inefficient, but could be confirmed using full-length pyrin (Fig. 6). Additionally, these experiments confirmed the stability of our proteins in vitro as shown in Figure 6 in the panels beneath the immunoprecipitation. These weak interactions were therefore subsequently confirmed using other strategies (see ASC speck binding assay, below).
Since ASC is also drawn to Listeria actin tails and to leading edge actin, we tested whether any of the seven GFP-tagged Listeria tail proteins described above interact directly with ASC. Previous studies had indicated that immunoprecipitations using ASC can be problematic due to its extreme insolubility (11). We therefore examined whether any of the seven proteins are recruited to ASC specks in transfected COS-7 cells. Cells co-transfected with ASC and the individual GFP-tagged proteins were examined 24 hrs later, at a time when specks are clearly visible in transfected cells. We were not able to detect the direct binding of any of these seven proteins to ASC specks (data not shown). Thus, the nature of the cellular protein that is responsible for the association between ASC and Listeria rocket tails is unclear.
Confirmation of Pyrin Binding Studies Using an ASC-Speck Binding Assay.
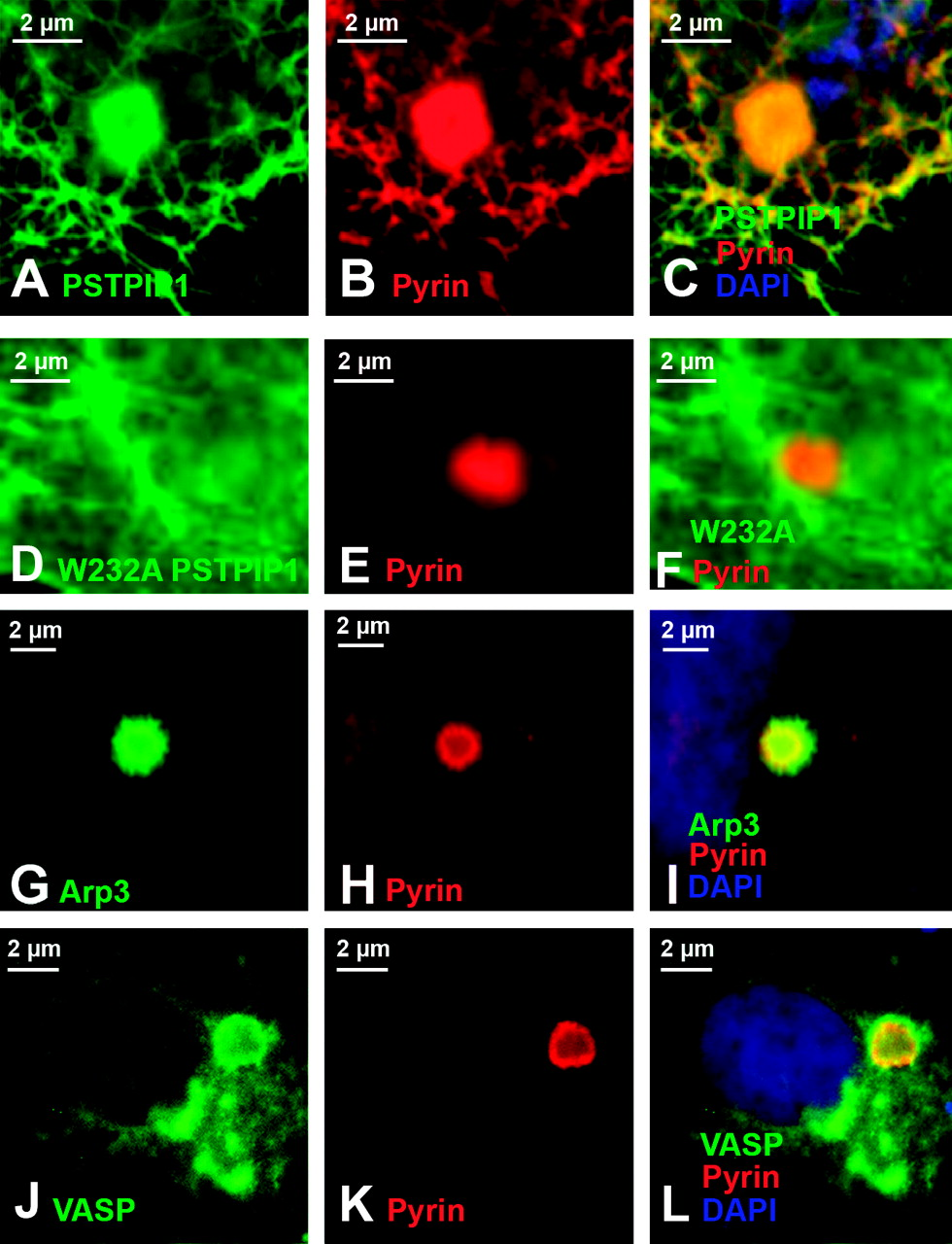
Because the pyrin immunoprecipitation results suggested rather weak interactions, we wished to confirm the binding using a different approach. Thus, we took advantage of a visual protein:protein interaction assay that we previously devised to examine pyrin-interacting proteins (12). Pyrin is known to co-localize with ASC specks, and the binding of pyrin to ASC requires the PyD of both proteins (the PyD of pyrin is encoded by exon 1, a region that is dispensable for pyrin’s recruitment to Listeria rocket tails, see Supplemental Fig. 1 and Fig. 4). Since the studies above indicate that ASC alone does not recruit the seven Listeria actin tail proteins to ASC specks, we used cells transfected with three proteins (ASC, pyrin, GFP-tagged test protein) to examine whether pyrin can recruit the other seven candidate binding proteins to ASC specks. Figure 7A–F validates the assay, showing that pyrin can recruit wild-type PSTPIP1 to the ASC speck (Fig. 7A–C), but is unable to recruit the W232A mutant form of PSTPIP1 that does not bind to pyrin (Fig. 7D–F). As shown in Figure 7, both Arp3 (panels G–I) and VASP (panels J–L) were carried to ASC specks by pyrin, while all other proteins tested were not, further validating the immunoprecipitation results.
Mutations in Pyrin Do Not Appear to Alter Its Association with Polymerizing Actin in Human Monocytes or in the Listeria Rocket Assay.
In monocytes from a patient with FMF (homozygous for the M694V mutation), pyrin and ASC were seen in membrane ruffles in a pattern indistinguishable from that seen in monocytes of unaffected individuals (Fig. 8A–C). Co-staining with phalloidin to identify polymerized actin and for pyrin revealed that pyrin co-localized with actin in membrane ruffles in these monocytes as well (Fig. 8D–F). In addition, migrating monocytes in which a leading edge could be identified also displayed a polarized distribution in which mutant pyrin was dramatically localized to the leading edge (Fig. 8G–I).
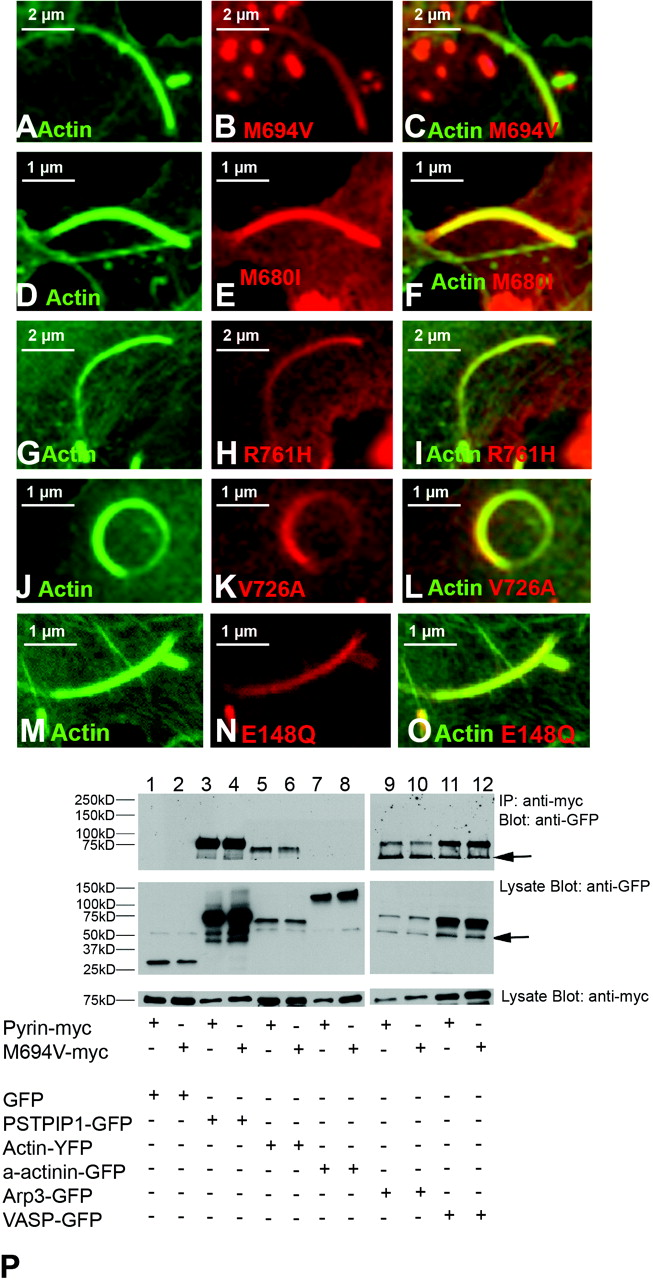
To further test the effect of FMF-associated pyrin mutations on the ability of pyrin to interact with polymerizing actin, we tested whether these mutations altered the ability of pyrin to concentrate in Listeria rocket tails. Several disease causing mutations, including M694V (Fig. 9A–C), M680I (Fig. 9D–F), R761H (Fig. 9G–I), V726A (Fig. 9J–L), E148Q (Fig. 9M–O), and P369S (data not shown) were tested. All mutations retained the ability to localize to Listeria tails. Next, we tested whether the M694V mutant form of pyrin was also able to bind to actin-associated proteins. We found that all forms of mutant pyrin showed binding to Arp3, VASP, and actin at levels comparable to wild-type pyrin (see Fig. 9P for M694V results). Additionally, we found that neither wild-type nor mutant pyrin bind to the actin-associated protein α-actinin.
Discussion
In this report, we demonstrate a clear link between pyrin, its interacting partner ASC, and a specialized portion of the cellular actin cytoskeleton. While in freshly isolated monocytes, pyrin and ASC are both associated with membrane ruffles throughout the cell, they undergo a dramatic polarization to the leading edge of migrating cells, where they co-localize with the actively polymerizing actin network. The re-localization appears nearly complete; little staining for either protein is detectable in other regions of the cell. This polarization suggests that both pyrin and ASC may play important roles in cytoskeletal signaling at the cell’s leading edge.
Our data indicate that pyrin’s affinity for the leading edge may be at least in part explained by the fact that it weakly interacts with actin, as well as with two proteins that play important roles in actin polymerization at the leading edge, VASP and Arp3. It is not possible to conclude at this time whether these interactions are co-dependent. That is, the association of pyrin with cellular actin could potentially depend partially on stabilization provided by its weak interactions with VASP and/or Arp3, and vis versa.
In human monocytes, the co-localization of pyrin and ASC at these specialized sites may be further reinforced by the previously recognized interaction between these two proteins. In contrast, in the Listeria assay in COS-7 cells, which do not express either ASC or pyrin, it can be concluded that the association of ASC with polymerizing actin does not depend on its association with pyrin, and likewise, the association of pyrin with these regions does not depend on ASC. Since ASC (at least in the conformation that it assumes in specks) does not appear to interact with actin, VASP and Arp3, independent mechanisms must stabilize the two proteins in the actin compartment of Listeria tails.
One of the domains of pyrin important for its localization to sites of active actin polymerization (the B-box and coiled coil) is identical to the domain required for pyrin’s interaction with PSTPIP1 (8). Since PSTPIP1 is homologous to cdc15, an actin-associated protein in yeast (29), we initially speculated that PSTPIP1 might be responsible for the recruitment of pyrin to the leading edge of migrating cells. However, infection of cells with Listeria does not result in the recruitment of PSTPIP1 to Listeria tails. When pyrin and PSTPIP1 are co-transfected into cells, pyrin co-localizes with PSTPIP1 filaments (data not shown). Yet, when these co-transfected cells are infected with Listeria, PSTPIP1 filaments are disrupted, but only pyrin is localized to tails (see Supplemental Fig. 2A–C). Thus, under these conditions, pyrin appears to prefer to associate with polymerizing actin rather than with PSTPIP1. It is possible that Listeria infection affects the phosphorylation status of PSTPIP1, as its binding to pyrin is highly dependent upon the level of PSTPIP1 phosphorylation (8).
Together, these data suggest that the association of pyrin with actively polymerizing actin is independent of pyrin’s association with ASC and PSTPIP1 and that PSTPIP1 does not spontaneously associate with these actin structures. However, PSTPIP1 can be recruited to the immunological synapse in T-cells, another site of active actin polymerization; this recruitment depends on WASP, a known substrate of PEST phosphatases, and a binding partner of PSTPIP1. Thus, under some regulated conditions, wild-type PSTPIP1 (independently of pyrin) is localized to sites of polymerizing actin. Our results are in agreement with a recent study of PSTPIP1 localization in neutrophils (30); in these cells, PSTPIP1 is localized to the trailing edge of the cell, the uropod. In this region, PSTPIP1 is associated with an actin compartment that is stable, rather than the dynamically polymerizing actin pool that is seen at the cell’s leading edge.
Mutations in PSTPIP1 cause PAPA syndrome, a debilitating inflammatory disease that begins in childhood with a sterile arthritis and progresses after puberty to massive granulomas and severe cystic acne (31). Previous work from the Kastner laboratory revealed that mutations in PSTPIP1 increase its affinity for pyrin, suggesting a molecular link in the disease etiology of FMF and PAPA syndrome (8). In accord with these data, we have seen that mutant forms of PSTPIP1 are drawn to actin tails by wild-type pyrin (Supplemental Fig. 2D–F); this recruitment depends on pyrin, as mutant PSTPIP1 by itself is not drawn to Listeria tails (Supplemental Fig. 2G–I). Since PSTPIP1 functions as an adaptor that links PEST phosphatases to their substrates, alterations in PSTPIP1 compartmentalization mediated by its increased binding to pyrin could potentially have important consequences for the phosphor-ylation status of important regulatory molecules that are known to bind to PSTPIP1 (e.g., WASP, c-Abl).
It is striking that of the more than 70 mutations that have now been identified in MEFV, the highest concentration can be found in the most C-terminal domain encoded by exon 10, a domain variously known as the Rfp or SPRY or B30.2 domain (21, 32). Though one of the regions of pyrin that is sufficient for its association with polymerizing actin is encoded by exons 6–10, our binding studies indicate that exon 10 is not necessary for this interaction. Indeed, mutations in exon 10 do not prevent recruitment of pyrin to Listeria rocket tails, nor do they cause a detectible reduction in mutant pyrin’s ability to bind both Arp3 and VASP. However, since binding to both Arp3 and VASP is quite weak, and may even be co-dependent, it is possible that there is an alteration in binding affinity that is simply too slight to be detected by immunoprecipitation. But, precisely because of this low affinity, minimal alterations have the potential to have major consequences for function.
The binding of pyrin to VASP and Arp3 explains pyrin’s localization in regions of polymerizing actin, and suggests a role in actin polymerization. Both Arp3 and VASP are components of the leading edge and Listeria rocket tails. Arp3 works in conjunction with Arp2 to nucleate actin filaments at the minus end, allowing rapid growth at the plus end. Additionally, the Arp2/3 complex can nucleate a branch from an existing actin filament, allowing the formation of a web of actin filaments. The Arp2/3 complex is consistently overexpressed in invasive colorectal cancers and contributes greatly to the cell’s increased motility and thus its ability to metastasize (33). VASP protects the growing ends of actin filaments from capping protein, which would stop their growth. It is associated with focal adhesions and is thought to be critical for cell motility. It has recently been proposed that an increase in the expression of VASP is involved in the progression and invasion of lung adenocarcinomas, and that this is likely due to an increase in cell motility (34). Additionally, cardiac fibroblasts from VASP null mice display increased cell spreading, but impaired migration and reorientation ability in a wounding assay (35). Thus, an attractive hypothesis, currently being tested, is that pyrin might modulate cell migration through its interaction with these proteins. In any case, the studies described here provide evidence that the connection between pyrin, ASC and the active actin cytoskeleton is more extensive than previously appreciated.
Pyrin, ASC, and actin expression in native and transfected cells. In monocytes (panels A–I), pyrin and ASC localized to supranuclear ruffles (A–F). Staining with phalloidin revealed that these ruffles were actin-based (G–I). In panels J–O, myc-tagged pyrin has been overexpressed in HeLa cells. Panels J and K represent different confocal planes of the same HeLa cell, and indicate that pyrin can simultaneously localize to both membrane ruffles (white arrow, J) and the edge of cells (white arrow, K). Co-staining for pyrin and actin reveals a similar pyrin and actin distribution in supranuclear ruffles (L). At increased magnification, it is possible to see that while pyrin co-localizes with actin at the edge of cells (white arrowheads), it does not co-localize with actin stress fibers (white arrow, M–O). All original images were captured at ×600 magnification. Pyrin distribution in monocytes. Treating cells with cytochalasin D to disrupt actin distribution also resulted in alteration of pyrin distribution (A–D). Confocal images of monocytes co-stained for pyrin and tubulin reveal that pyrin is located in supranuclear ruffles and does not co-localize with microtubules (E–F). All original images were captured at ×600 magnification. Pyrin co-localizes with regions of polymerizing actin. In migrating monocytes, distribution of pyrin (A–C) and ASC (D–F) is polarized. Both proteins are concentrated in the actin-rich leading edge of migrating cells. In the Listeria rocket assay, COS-7 cells were infected with Listeria monocytogenes and their actin-based tails were used to recruit actin polymerizing machinery. Listeria tails do not recruit the negative control protein myc-LacZ (G–I), but do recruit both myc-tagged pyrin (J–L) and myc-tagged ASC (M–O). All original images were captured at × 600 magnification. Regions of pyrin required for recruitment to Listeria rocket tails. Several myc-tagged regions of pyrin were tested for their ability to localize to Listeria rocket tails. Two minimal regions of pyrin were found to be sufficient for this recruitment: exons 3–5 and exons 6–10. Since these regions encompass the B-box and coiled-coil domains of pyrin, we determined that these domains were critical for pyrin’s recruitment to regions of polymerizing actin. PSTPIP1 is not recruited to Listeria rocket tails. PSTPIP1 is a known actin-associated protein that also binds to pyrin. Despite this known interaction, FLAG-tagged PSTPIP1 is not recruited to Listeria rocket tails. All original images were captured at ×600 magnification. Pyrin interacts with actin, VASP, and Arp3 by immunoprecipitation. Immunoprecipitation experiments revealed that full-length myc-tagged pyrin precipitates YFP-tagged actin, as well as GFP-tagged VASP and Arp3. These interactions were weak as compared with the positive control protein PSTPIP1-GFP, but were reproducible. Pyrin recruits VASP and Arp3 to ASC specks. The ASC speck assay is a recently described protein:protein interaction assay. The validity of this assay is demonstrated by myc-pyrin’s ability to recruit wild-type FLAG-PSTPIP1, which is normally not seen in specks, to ASC specks (A–C). However, a mutated version of PSTPIP1, FLAG-W232A, that does not interact with pyrin is not recruited to specks (D–F). Myc-pyrin recruits GFP-tagged versions of both Arp3 (G–I) and VASP (J–L) to specks, verifying the interaction between these proteins. All original images were captured at ×600 magnification. Effect of pyrin mutations on pyrin distribution. Monocytes isolated from an FMF patient homozygous for the M694V mutation were examined for pyrin, ASC, and actin distribution. Both ASC and pyrin were found in supranuclear ruffles (A–C) in a pattern indistinguishable from that of wild-type monocytes. Co-staining for actin (using phalloidin) and pyrin revealed that the two proteins co-localize in these ruffles (D–F). In migrating monocytes, pyrin was again seen concentrated in the leading edge (G–I). All original images were captured at ×600 magnification. Effect of pyrin mutations on protein:protein interactions. FMF-causing mutant versions of myc-tagged pyrin were tested in the Listeria rocket assay. Here we show five of the mutations tested: M694V (A–C), M680I (D–F), R761H (G–I), V726A (J–L), and E148Q (M–O). In all cases, mutant pyrin was recruited to Listeria rocket tails. The ability of M694V mutant pyrin to bind to actin-associated proteins was then tested using immunoprecipitation. M694V bound to GFP-tagged PSTPIP1, actin (YFP-tagged), VASP, and Arp3 at levels comparable to wild-type pyrin (P). All original images were captured at ×600 magnification.

Footnotes
This work was supported by (NIH R01-AI053262). P.S. was supported by the Organogenesis Training Program (NIH/NICHD T32-HL07505).
Acknowledgements
The authors thank Dan Kastner for providing the α-human pyrin antibody; Maria Diakonova for providing the GFP and YFP tagged constructs; Kristen Verhey and Joel Swanson for valuable advice; the University of Michigan Organogenesis Morphology Core for microscopy services.