
Abstract
Glucocorticoids and mineralocorticoids modulate Na+ transport via epithelial Na+ channels (ENaC). The rat submandibular epithelial cell line, SMG-C6, expresses α-ENaC mRNA and protein and exhibits amiloride-sensitive Na+ transport when grown in low-serum (2.5%) defined medium, therefore, we examined the effects of altering the composition of the SMG-C6 cell growth medium on ENaC expression and function. No differences in basal or amiloride-sensitive short-circuit current (Isc) were measured across SMG-C6 monolayers grown in the absence of thyroid hormone, insulin, transferrin, or EGF. In the absence of hydrocortisone, basal and amiloride-sensitive Isc significantly decreased. Similarly, monolayers grown in 10% serum-supplemented medium had lower basal Isc and no response to amiloride. Adding hydrocortisone (1.1 μM) to either the low or 10% serum medium increased basal and amiloride-sensitive Isc, which was blocked by RU486, the glucocorticoid and progesterone receptor antagonist. Aldosterone also induced an increase in α-ENaC expression and Na+ transport, which was also blocked by RU486 but not by the mineralocorticoid receptor antagonist spironolactone. Thus, in the SMG-C6 cell line, hydrocortisone and aldosterone increased ENaC expression and basal epithelial Na+ transport. The absence of endogenous ENaC expression in culture conditions devoid of steroids makes the properties of this cell line an excellent model for investigating pathways regulating ENaC expression and Na+ transport.
Introduction
Epithelial Na+ channels (ENaC) regulate the transport of Na+ across tight epithelia lining several organ systems (1). Identified in the apical epithelium of many tissues including lung, kidney, distal colon, sweat and salivary ducts (2–4), ENaC activity comprises the rate-limiting step for Na+ entry into the cell. Na+ is then extruded into the interstitium via Na+-K+-ATPase located in the basolateral membrane (3). ENaC is a multimeric protein complex consisting of three homologous subunits (α, β, and γ) (5). When the subunits are independently expressed in Xenopus laevis oocytes or rat thyroid epithelial cells, only the α-subunit is capable of actively transporting Na+ (6, 7). Neither the β- nor γ-subunits form functional Na+ channels when expressed alone; however, coexpression with α-subunits greatly enhances Na+ current (6, 7).
A variety of hormones and growth factors; including glucocorticoids, mineralocorticoids, catecholamines, insulin, and epidermal growth factor; as well as various in vitro culture conditions such as oxygen concentration, alter the expression or activity of the amiloride-sensitive Na+ channel in a cell-specific manner (8–11). Glucocorticoids effectively upregulate ENaC activity in renal collecting duct cells (12). Glucocorticoids also upregulate all three ENaC subunits and Na+ uptake in bronchial epithelium (13) and stimulate amiloride-sensitive current in cultured lung explants across pulmonary epithelial monolayers (14, 15). Insulin upregulates ENaC expression and function in renal cells (16, 17); however, EGF increases amiloride-insensitive Na+ transport and decreases ENaC subunit expression in alveolar cells (18). Mineralocorticoids, such as aldosterone, also play an important role in the upregulation of ENaC activity in renal tissues and segments of the gastrointestinal tract (3). In the apical membrane of the A6 cell line, aldosterone increases the expression of all three ENaC subunits (19). Aldosterone has been shown to stimulate Na+ channels in adult rat alveolar type II epithelial cells (20).
The SMG-C6 immortalized rat submandibular epithelial cell line is derived from the acinar gland and appears to exhibit properties of both acinar and ductal salivary epithelial cells (2, 21). This cell line expresses ENaC mRNA and protein, and its monolayers form tight junctions with a predominantly amiloride-sensitive Na+ current (22). The cell line is grown in a low-serum (FBS 2.5%) defined growth medium supplemented with the following: transferrin, hydrocortisone, retinoic acid, triiodothyroxine, insulin, and epidermal growth factor. In the present study, we altered the composition of the SMG-C6–defined culture medium and measured whether ENaC expression and activity would be altered. Since both glucocorticoids and mineralocorticoids affect Na+ transport, we also examined the effects of aldosterone on Na+ transport in SMG-C6 epithelial cells.
Materials and Methods
Materials.
Dulbecco’s modified Eagles Medium (DMEM), Ham F-12, fetal bovine serum (FBS), and trace element mix were obtained from Gibco (Grand Island, NY). Transferrin and epidermal growth factor (EGF) were obtained from JRH Biosciences (Lenexa, KS). Insulin was obtained from Novo Nordisk Pharmaceuticals (Princeton, NJ). Retinoic acid, triiodothyroxine, hydrocortisone, aldosterone, spironolactone, and amiloride were purchased from Sigma Chemical Co. (St. Louis, MO). RU486 was obtained from BioMol (Plymouth Meeting, PA). The anti-rat α-, β-, and γ-ENaC antibodies were obtained from Affinity BioReagents (Golden, CO). All other antibodies used in this study were purchased from Santa Cruz Biotechnology (Santa Cruz, CA).
Cell Culture.
SMG-C6 cells passages 28–44 were grown in one of the following defined DMEM/F12 1:1 growth media: (1) 2.5 % FBS (low serum) supplemented with 5 μg/ml transferrin, 1.1 μM hydrocortisone, 0.1 μM retinoic acid (RA), 2.0 nM triiodothyroxine (T3), 5 μg/ml insulin, 80 ng/ml epidermal growth factor (EGF), a trace element mix (5 ml/500 ml), 100 U/ml penicillin and 100 μg/ml streptomycin, (2–7) the same low serum medium described above in the absence of either transferrin, hydrocortisone, retinoic acid, T3, insulin, or EGF, (8, 9) 10% FBS (high serum) with or without 1.1 μM hydro-cortisone, (10–12) the low serum defined medium described in (1) in the absence of hydrocortisone plus either 1 μM aldosterone alone or in combination with either 10 μM RU486 or 10 μM spironolactone. All cells were grown in 25 cm2 flasks and cultured at 37°C with 5% CO2 in an incubator. Once cells reached confluence in the flasks, cells were washed with Hanks buffered salt solution, and 0.25% trypsin was added to remove cells. The cells were counted with a Coulter® Multisizer II and seeded at 50–100,000 cells/cm2 on Snapwell filters (diameter 12 mm, pore size 0.4 μm) coated with human collagen type-1 (Becton Dickinson, Franklin Lakes, NJ). Cells seeded on the permeable supports were maintained in their designated culture medium.
Transepithelial Transport Studies.
The transepithelial resistance (Rt) of the SMG-C6 monolayers was measured daily by an EVOM-G (World Precision Instruments, WPI, Sarasota, FL). Filters exhibiting Rt greater than 800 Ω·cm2 were placed in a modified Ussing chamber equipped with Ag/AgCl electrodes and bathed with Krebs-Ringer bicarbonate (KRB) medium containing 120 mM NaCl, 2.5 mM K2HPO4, 0.6 mM KH2PO4, 1.2 mM CaCl2, 1.2 mM MgCl2, 20 mM NaHCO3, and 10 mM glucose at a pH of 7.4. In the chamber KRB medium was kept at 37°C and airlifted with 5% CO2/21% O2. Using a DVC-1000 Voltage/Current Clamp (WPI) with automatic fluid resistance compensation, stable baseline potential difference (PD) was measured. After allowing equilibration, the PD was clamped to 0 mV, and the short circuit current (Isc) was continuously recorded. Transepithelial resistance was calculated using Ohm’s law (Rt = ΔPD/Δ Isc) by clamping the monolayer PD to 1 mV for 0.5 seconds every 30 seconds and measuring the change in current. The PD is measured in the apical fluid and is negative relative to the basolateral side. The measured Isc reflects the active transport of positive ions from the apical to basolateral side. In this cell line, the predominant current is from Na+ movement from the apical side, and the administration of amiloride (10 μM) virtually eliminates the basal current likely by blocking Na+ entry into the cell (22). After Isc reached steady state, baseline values were recorded, and the amiloride-sensitive Isc was measured as the difference in current in the presence and absence of amiloride. Amiloride was dissolved in DMSO, with the final dilution in the Ussing chamber fluid being 1:1000. This amount of DMSO did not alter bioelectric properties, and previous studies established this concentration of DMSO in amiloride does not alter Rt, indicating that changes in transepithelial Isc was a direct effect of the specific agent studied. Neither basal bioelectric properties nor the response to specific agents varied following each cell passage.
To measure the basolateral Na+-K+-ATPase pump activity, SMG-C6 monolayers were first treated with apical amiloride (10 μM) followed by nystatin (50 μM) to permeabilize the apical membrane. Treatment with nystatin increases apical Na+ conductance, and the resultant Na+ transport via the Na+-K+-ATPase pump becomes the rate-limiting step. Ouabain (1 mm) was added to the basolateral bath, and the change in the current, ouabain-sensitive current (Iouabain), was measured as an indicator of Na+-K+-ATPase activity (23, 24).
Western Blot Analysis.
After culturing SMG-C6 cells in the various media for 24 hours, the cells were rinsed and then homogenized in lysis buffer (Tris-HCL 50 mM, pH 7.4, TBS) containing 5 mM EDTA, 5 mM EGTA, leupeptin (1 μM), pepstatin A (1 μM), aprotinin (1 μM), and phenylmethylsulphonyl fluoride (1 μM). Protein concentrations were determined using the BioRad DC protein assay (Hercules, CA), with BSA used as the standard. Fifty to sixty micrograms of total protein was subjected to SDS-PAGE (7.5% gel) using the buffer system of Laemmli (25). The separated proteins were transferred electrophoretically to PVDF membranes (Millipore, Bedford, MA) using a semi-dry transfer blot system. The proteins were blocked in TBS containing 5% non-fat dried milk powder for 1 h and then incubated with primary antibody in TBS containing 0.01% Tween-20 and 5% non-fat dried milk powder for 24 h. The blots were then incubated with secondary antibody conjugated to horseradish peroxidase appropriately diluted in the same buffer for 1 h. Peroxidase-labeled proteins were visualized using an enhanced chemiluminescence (ECL) assay kit (Amersham Pharmacia Biotech, England). The relative intensities of the bands were quantified by densitometry using Image J.
Statistical Analysis.
Data are expressed as the mean with the corresponding standard error of the mean (SEM). Comparisons of amiloride sensitive Isc as well as densitometric analysis of Western blots following each treatment were made by the unpaired Student’s t test or ANOVA where applicable. Significance was accepted at a P value < 0.05.
Results
Effect of Growth Factors and Hormones on Na+ Transport and ENaC Expression.
SMG-C6 cells were seeded on permeable filters and grown in the low-serum defined media in the presence or absence of specific supplements. In all the various growth media, there were no differences in the time it took the filters to reach confluence (as reflected in the daily Rt measurements, and there were no differences in the Rt values once mounted in the Ussing chambers. The basal PD and Isc measured across the monolayers before and after the administration of 10 μM amiloride to the apical side are listed in Table 1. In the low-serum defined medium with all the supplements, amiloride inhibited approximately 90% of basal Isc (from 15.6 ± 1.7 to 1.3 ± 0.2 μA/cm2). Our laboratory has previously demonstrated that basal Isc across SMG-C6 monolayers is primarily transepithelial Na+ current and is inhibited by submicromolar concentrations of amiloride (K0.5 ~392 nM) (22). In the low-serum defined medium without supplemental hydrocortisone, the spontaneous and amiloride-sensitive Isc were significantly lower (1.6 ± 0.2 μA/cm2 and 1.1 ± 0.2 μA/cm2, respectively, Fig. 1A). In contrast, if the SMG-C6 monolayers were grown in the low-serum defined medium with hydrocortisone but without transferrin, T3, insulin, retinoic acid, or EGF, the amiloride-sensitive Isc was not significantly altered. Whole cell lysates derived from cells grown in a medium devoid of individual supplements were probed using a rabbit polyclonal anti-α-ENaC antibody for α-ENaC protein. α-ENaC protein was significantly decreased only when cells were grown in the absence of hydrocortisone (Fig. 1B and Fig. 2). In contrast the expression of the β-ENaC and γ-ENaC protein levels remained unaltered by the addition of hydrocortisone to the culture medium (Fig. 2).
Effect of Serum on Electrogenic Na+ Transport and ENaC Expression.
If SMG-C6 monolayers were grown in a medium (Ham F-12/DMEM) supplemented with only 10% FBS, the amiloride-sensitive Isc was 0.6 ± 0.3 μA/cm2 (Fig. 3A). Although the concentration of hydrocortisone in commercially available FBS can vary, the calculated concentration in the 10% medium in our laboratory was 0.06 μM. When hydrocortisone (1.1 μM) was added to the 10% FBS medium, the amiloride-sensitive Isc after 24 hours increased to 18.7 ± 5.2 μA/cm2. Similarly treated cells were subjected to Western blot analysis. α-ENaC protein was increased after the addition of hydrocortisone for 24 hours (Fig. 3B).
Effect of Glucocorticoid and Mineralocorticoid Inhibitors.
To further characterize the glucocorticoid response, monolayers were treated with both hydrocortisone and RU486, a potent glucocorticoid and progesterone receptor antagonist, for 24 hours. As shown in Figure 4A, the amiloride sensitive current is significantly decreased in the presence of RU486 (from 14.3 ± 1.7 to 0.1 ± 0.01 μA/cm2). Whole cell lysates exposed to hydrocortisone or hydrocortisone plus RU486 were probed using α-, β- and γ-ENaC antibody (Fig. 4B). Neither hydrocortisone in the presence or absence of RU486 induced a change in β- or γ-ENaC protein; however α-ENaC expression was decreased in the presence of RU486. Since aldosterone upregulates electrogenic Na+ transport in several epithelial tissues, SMG-C6 monolayers were grown in 2.5% FBS medium supplemented with aldosterone (1 μM) without hydrocortisone. In the presence of aldosterone, the amiloride-sensitive Isc (17.6 ± 3.1 μA/cm2) was similar to hydrocortisone-treated monolayers. When the mineralocorticoid receptor antagonist, spironolactone (10 μM), was added to the aldosterone-supplemented medium, the resulting amiloride-sensitive Isc was not altered, but co-treatment with RU486 did significantly decrease electrogenic Na+ transport (Fig. 5A). The expression of α-ENaC protein was also decreased in cells treated with aldosterone and RU486 but not in cells treated with aldosterone and spironolactone (Fig. 5B). These findings indicate that the effects of aldosterone in this cell line are likely to be mediated through the glucocorticoid receptor. Finally, Figure 5C demonstrates that despite the aldosterone increase in α-ENaC protein expression with supplemental aldosterone, there was no effect on the β- or γ-subunits.
Effect of Glucocorticoid or Mineralocorticoid on Na+-K+-ATPase Activity.
Both glucocorticoids and mineralocorticoids have been shown to upregulate Na+-K+-ATPase activity in various cell types, generating the Na+ gradient necessary to support Na+ entry (14, 24, 26). To quantify the effects of hydrocortisone or aldosterone on Na+ pump activity, nystatin was added to the apical bathing solution to increase the permeability of the amiloride-treated apical membrane (23). Following equilibration of the resultant rise in current, ouabain (1 mM) was added to the basolateral solution, and the change in current (IOuabain)) was measured. IOuabain across monolayers grown in the presence or absence of hydrocortisone or aldosterone were similar (Fig. 6A). These data suggest that in this cell line glucocorticoid and mineralocorticoid supplementation increased transepithelial Na+ movement primarily by increasing amiloride-sensitive Na+ entry rather than by stimulating basolateral Na+-K+-ATPase activity. Western blot analysis also demonstrated that in the SMG-C6 cell line, Na+-K+-ATPase-α1 protein expression was unchanged in the presence or absence of hydrocortisone or aldosterone (Fig. 6B).
Discussion
ENaC expression and function in various epithelia are regulated by glucocorticoids, mineralocorticoids, and various growth factors. Dexamethasone, prednisone, and hydrocortisone have all been found to increase amiloride-sensitive Isc in bovine mammary epithelial cells; however, dexamethasone increased β- and γ-ENaC mRNA without increasing α-ENaC mRNA (27). In contrast, in primary cultures from fetal rat distal lung epithelia and cultured fetal lung explants, dexamethasone increased α-ENaC mRNA levels (10, 28, 29). Dexamethasone also upregulated α-, β- and γ-ENaC mRNA expression and α-ENaC function in a bronchiolar cell line (H441) and in adult rat alveolar epithelial cells (13, 23, 30). Mustafa et al showed that dexamethasone induced α-ENaC formation in preterm rabbit lungs (31). Furthermore, dexamethasone increased α-ENaC mRNA in mouse fetal lung kidney explants (32) and in rat kidney cells (33).
Stimulation of ENaC expression and activity by mineralocorticoids was evident in cells from kidneys (12) and the colon (34, 35). The induction of ENaC subunits by aldosterone appeared to be tissue specific. In bovine mammary epithelial cells, aldosterone had no effect on the basal or amiloride sensitive Isc (27). In colon epithelia and renal cells (A6), aldosterone enhanced the transcription of β- and γ-ENaC subunits, while α-ENaC was constitutively expressed (36, 37). In contrast, aldosterone evoked an increase in α-, but not β- or γ-ENaC mRNA and protein in the rat kidney (36, 38). Besides affecting ENaC subunit expression, aldosterone also increased ENaC trafficking and subunit incorporation into the apical membrane (16, 17).
In adult rat alveolar type II cells, aldosterone increased amiloride-sensitive alveolar fluid clearance, a response inhibited by spironolactone (20), indicating that aldosterone effects were mediated by mineralocorticoid receptors. In contrast, our findings in the SMG-C6 cell line demonstrate that the aldosterone-induced increase in ENaC expression and function were not altered by spironolactone, and instead the effects were mediated via the glucocorticoid receptor. This finding is similar to that in bovine mammary epithelial cells where RU486, but not spironolactone, decreased the amiloride sensitive Isc produced by corticosteroids (27). Mineralocorticoid receptors are not present until late in gestation in many mammals. In the human fetus there is a maturational increase in glucocorticoid receptor mRNA expression between 8–17 weeks gestations, with much lower levels of mineralocorticoid receptor mRNA concentrated primarily in the bronchial columnar epithelial cells and not in alveolar cells (39).
Besides glucocorticoids and mineralocorticoids, other hormones and growth factors have been shown to influence ENaC activity. Vasopressin and insulin also increase Na+ transport by increasing the membrane number of ENaC subunits (16, 17). Although T3 alone did not have a direct effect, in lung explants it potentiated the effect of hydrocortisone on amiloride-sensitive current (15). In fetal lung explants, keratinocyte growth factor (KGF) decreased α-ENaC expression (40). Chronic exposure to EGF reduced transepithelial Na+ absorption by inhibiting amiloride-sensitive Na+ entry into renal collecting duct cells (24). EGF was also shown to increase Na+ conductance; however, this reflected an increase in amiloride-insensitive Na+ transport and decrease in ENaC subunit expression (18). In the SMG-C6 cell line, transferrin, T3, insulin, retinoic acid, or EGF did not alter amiloride-sensitive Isc or α-ENaC expression. Therefore, the effects of these growth factors appear to be cell or tissue specific.
The SMG-C6 cell line derived from rat submandibular acinar glands expresses specific acinar cell proteins and retains receptor-stimulated mobilization of intracellular signaling elements (Ca2+ and cAMP) comparable to intact salivary glands or freshly isolated cells (21, 41). This cell line is grown in a defined medium with low (2.5%) serum supplemented with hormones and growth factors vital for the maintenance and growth of salivary acinar cells (42). When grown on permeable supports, allowing for exposure to media on both sides of the cells, the monolayers displayed clear evidence of polarization with stable transepithelial resistance, facilitating bioelectric characterization in Ussing chambers. As would be expected from salivary acinar cells, agonist-stimulated Cl− secretory conductance is present. However, the majority of basal current was found to be sensitive to apical amiloride (22). Consistent with the vectorial Na+ transport measured under Isc conditions, SMG-C6 cells were found to express ENaC subunits. ENaC is not present in the acinar portion of the salivary gland but is expressed in the apical membrane of the striated and interlobular ducts (43). Patch-clamp technique supports a role for ENaC in regulating the volume and composition of secondary saliva in the salivary duct (2). The rat parotid acinar cell line immortalized from the same laboratory has also been found to express glucocorticoid-induced α-ENaC transcript (44, 45).
Changes in α-ENaC protein levels correlated with the functional changes seen in basal and amiloride-sensitive Isc. Our data are also consistent with a model that correlates changes in ENaC activity with α-ENaC upregulation since no increases in β- or γ-ENaC subunits were seen. When independently expressed in Xenopus laevis oocyte or rat thyroid cells, the α-subunit was found to be essential for Na+ transport (6, 7). In contrast, upregulation or insertion of the β-ENaC subunit into the A6 cell apical membrane altered sodium influx (37), whereas corticosteroids increased β- and γ-, but not α-ENaC gene expression in intestinal epithelium (33). In the SMG-C6 cells, endogenous levels of α-ENaC mRNA were virtually absent when grown without hydrocortisone (46, 47). Therefore, the regulation of ENaC subunits appears to be species and tissue specific.
Since RU486, and not spironolactone, inhibited aldosterone-induced stimulation of ENaC activity in the SMG-C6 cells, the response to aldosterone was dependent on binding with the glucocorticoid receptor. In renal cells, stimulation of ENaC channel activity by aldosterone has been shown to occur in two phases, an initial increase in Na+ conductance (within 1 hour), thought to involve activation or insertion of silent or inactive channels into the apical membrane, and a late phase (18–24 hours), which is dependent on genomic activation and synthesis of ENaC protein subunits (19, 36). Similar to cells from the distal colon, the SMG-C6 Na+ transport likely requires channel de novo synthesis (35). Both glucocorticoids and mineralocorticoids regulate the transcript expression of the regulatory protein sgk1 (12, 48). sgk1 has been shown to mediate the early response by aldosterone responsive cells and to also phosphorylate Nedd4–2 (49). The binding of Nedd4 family decreases Na+ current by reducing ENaC expression at the cell surface. In contrast, phosphorylated Nedd4–2 is unable to bind with the PY motif of ENaC subunits, thus maintaining ENaC function at the cell membrane (49). Alternatively, the data could also be explained in part by the reduced retrieval and degradation of ENaC by hydrocortisone and aldosterone. Further work is in progress to investigate the effects of glucocorticoids and the interactions of sgk1 and Nedd4–2 on this cell line.
Finally, in some tissues or cells, activation of Na+-K+-ATPase may increase Na+ transport by increasing the electrochemical driving force for Na+ entry. Glucocorticoids and mineralocorticoids as well as other hormones and growth factors have been shown to alter Na+-K+-ATPase function (26). Similarly, the activation of pre-existing transport proteins, independent of Na+ cell entry, increased Na+ transport (50). In this cell line, the ouabain-sensitive current (Iouabain) was unaltered by hydrocortisone or aldosterone, indicating that Na+ pump activity was not the primary force for increased Na+ current.
In summary, α-ENaC protein expression and Na+ transport were significantly reduced in SMG-C6 cells when cultured in medium devoid of glucocorticoids or mineralocorticoids. We speculate that the absence of electrogenic Na+ transport is due to the reduced expression of Na+ channels rather than increased degradation. As there is very little ENaC expression under these culture conditions, this cell line serves as an excellent model for investigating the mechanistic/signaling pathways by which specific agonists upregulate ENaC expression and Na+ transport.
a
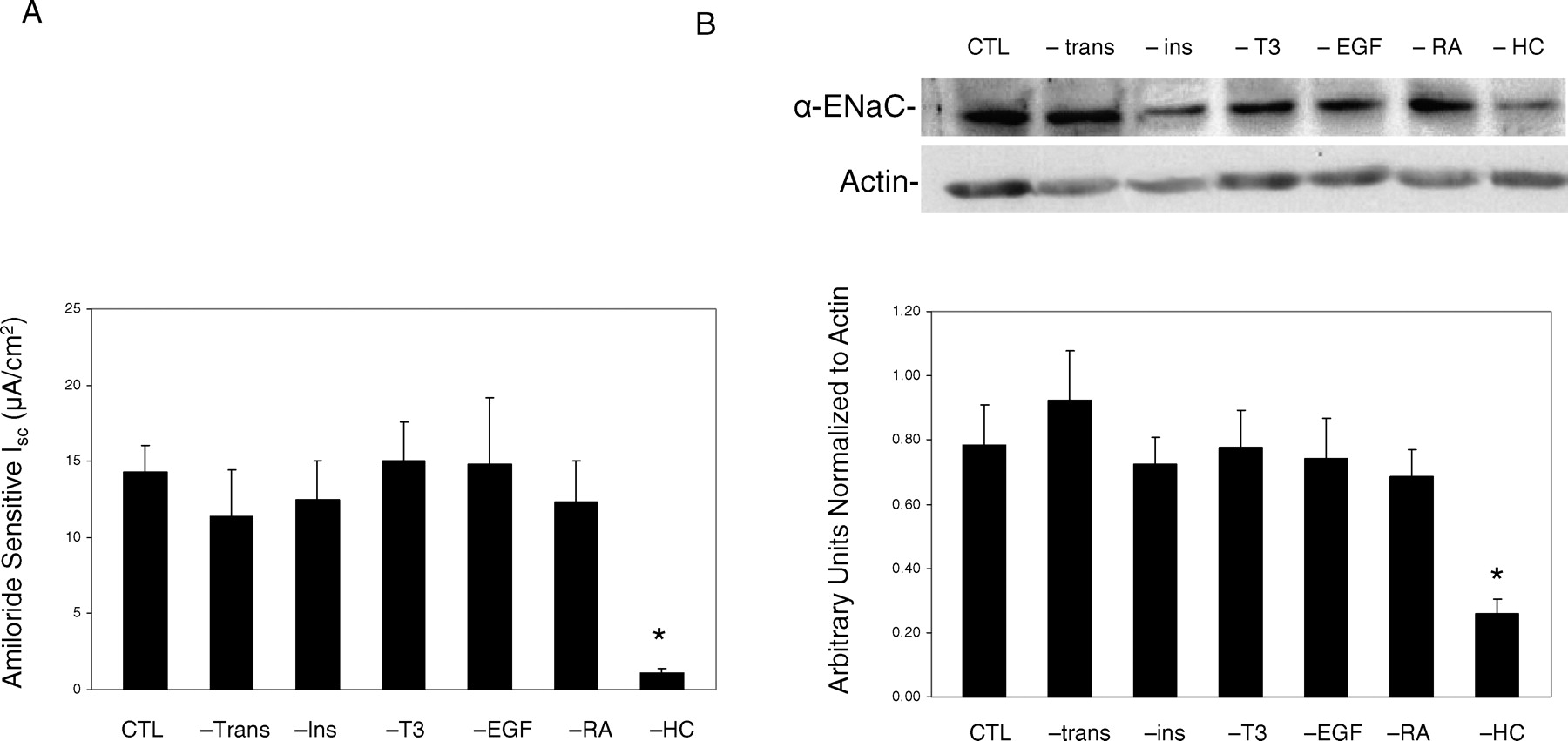
A) Effect of removing specific hormone or growth factor from low-serum (2.5%) defined growth media on amiloride-sensitive Isc across SMG-C6 monolayers. Monolayers were grown in defined or control (CTL) media in the absence of transferrin (trans), T3, insulin (Ins), EGF, retinoic acid (RA), or hydrocortisone (HC). +HC (CTL) and –HC were used as controls (n = 16) for each supplement removal (n = 4–7). B) Protein expression via a representative Western blot of cells grown in control media or in media devoid of specific growth factors or hormones. Graph depicts densitometric analysis normalized to actin. n = 6 for each experiment. *P < 0.05 vs CTL media.
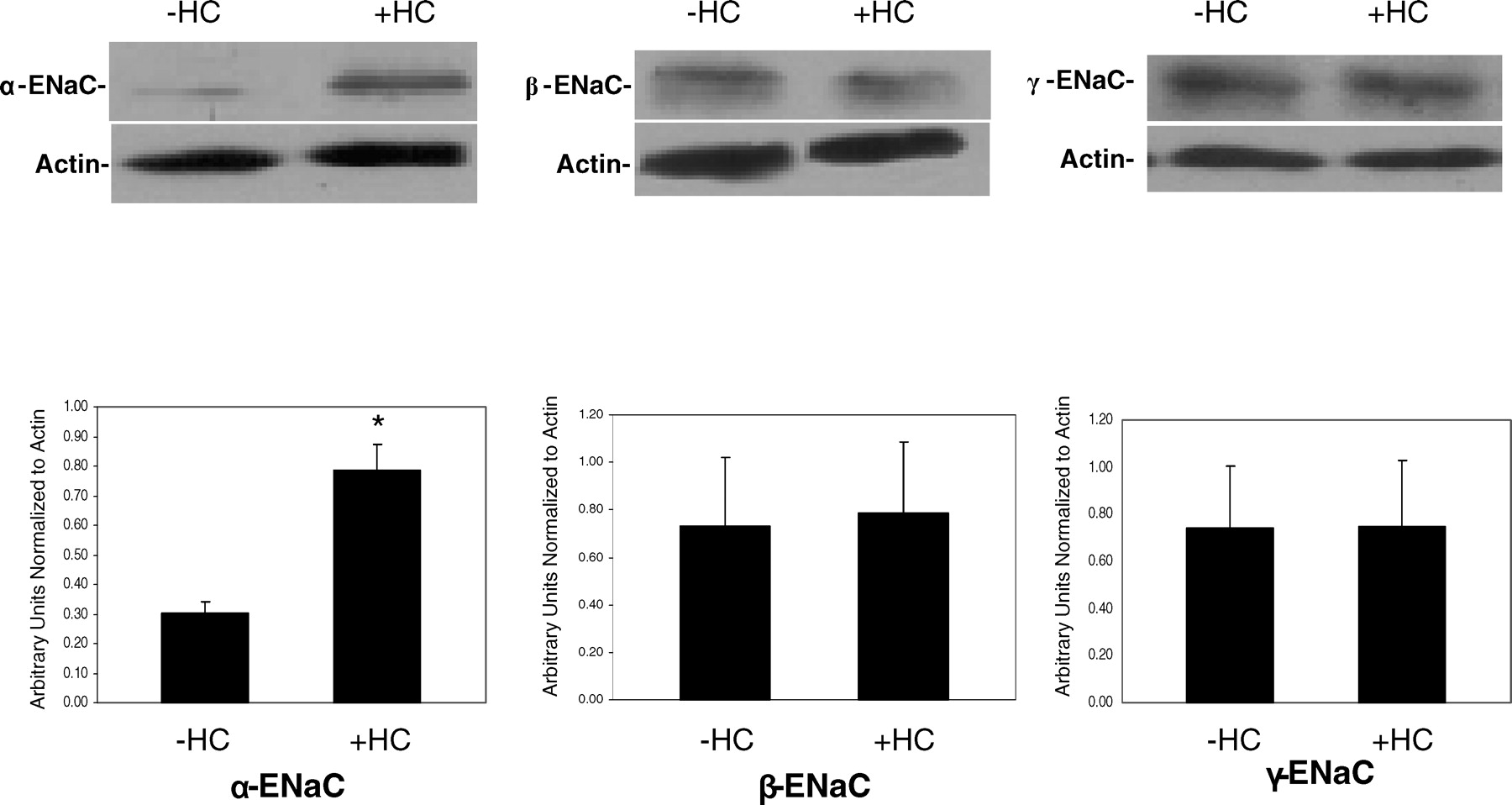
Representative Western blot and densitometric analysis for cells grown in the presence and absence of hydrocortisone (HC). Whole cell lysates were subjected to Western blot analysis as described in the methods section using α-, β-, and γ-ENaC subunit antibodies. n = 8 for α-ENaC and n = 6 for β- and γ-ENaC.
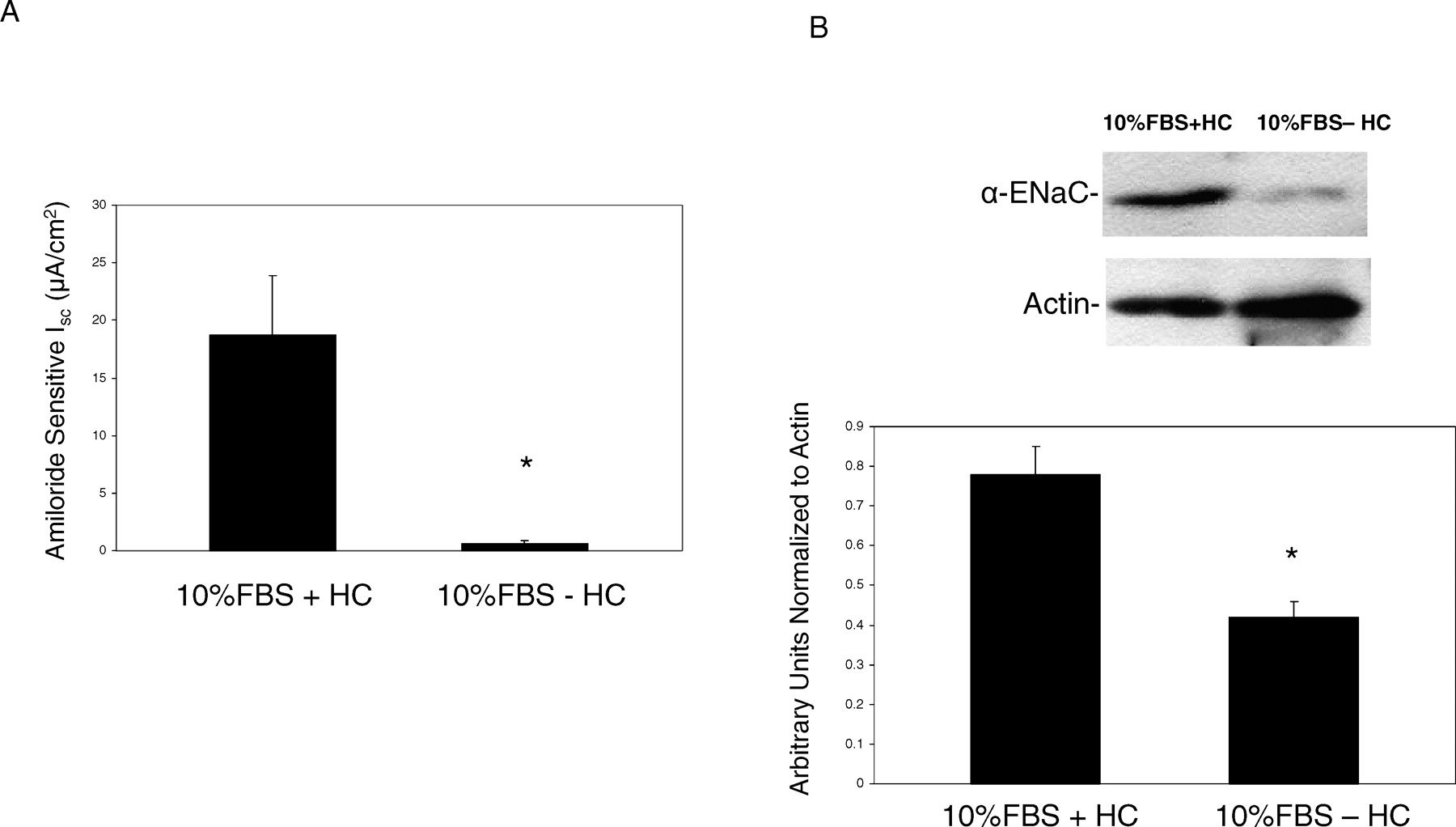
A) Amiloride-sensitive Isc across SMG-C6 monolayers grown in DMEM/F-12 with 10% FBS supplemented media with and without HC (n = 5). *P < 0.05 vs 10% FBS + HC. B) Representative Western blot and densitometric analysis of cells grown in 10% FBS either with or without HC (n = 3). *P < 0.05 vs 10% FBS + HC.
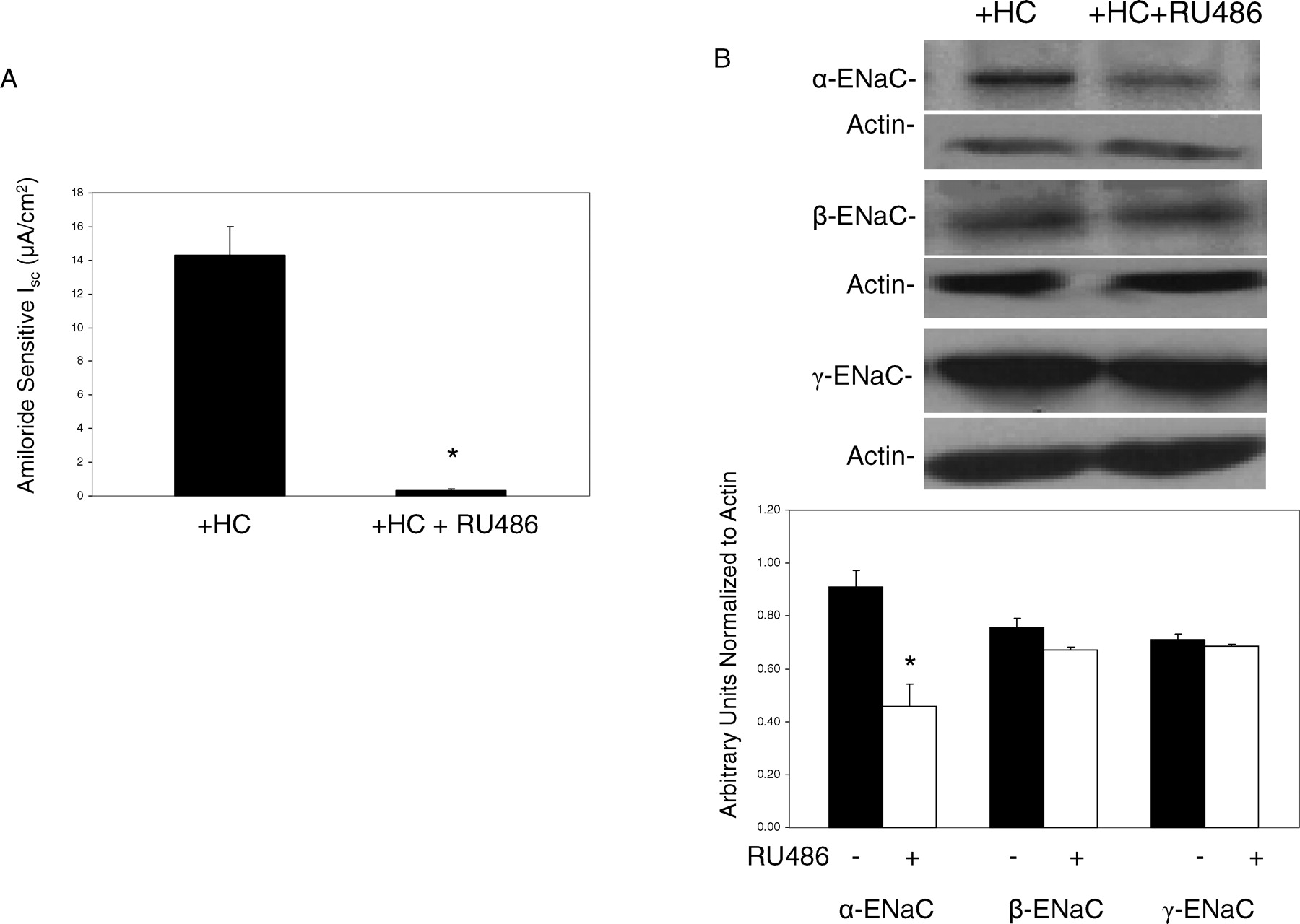
A) Effect of the glucocorticoid and progesterone receptor antagonist, RU486 (RU), on amiloride-sensitive Isc in SMG-C6 monolayers grown in the presence of hydrocortisone (n = 4). B) Representative Western blots and densitometric analysis normalized to actin demonstrating the effects of RU486on α-, β-, and γ-ENaC subunits (n = 6 for each experiment). *P < 0.05 vs hydrocortisone (HC) without RU486.

A) Effect of 48h incubation with aldosterone (Aldo, 1 μM) plus spironolactone (Sp, 10 μM) or RU486 (RU, 10 μM) on amiloride-sensitive Isc across SMG-C6 monolayers (n = 4–5). *P < 0.05 vs Aldo alone. B) Western blot representation of α-ENaC protein in cells treated with aldosterone plus spironolactone or RU486 (n = 6). *P < 0.05 vs Aldo alone. C) Western blot and densitometric analysis of the effect of aldosterone on α-, β-, and γ-ENaC protein (n = 5–6 for each experiment). *P < 0.05 compared with no hydrocortisone (−HC).
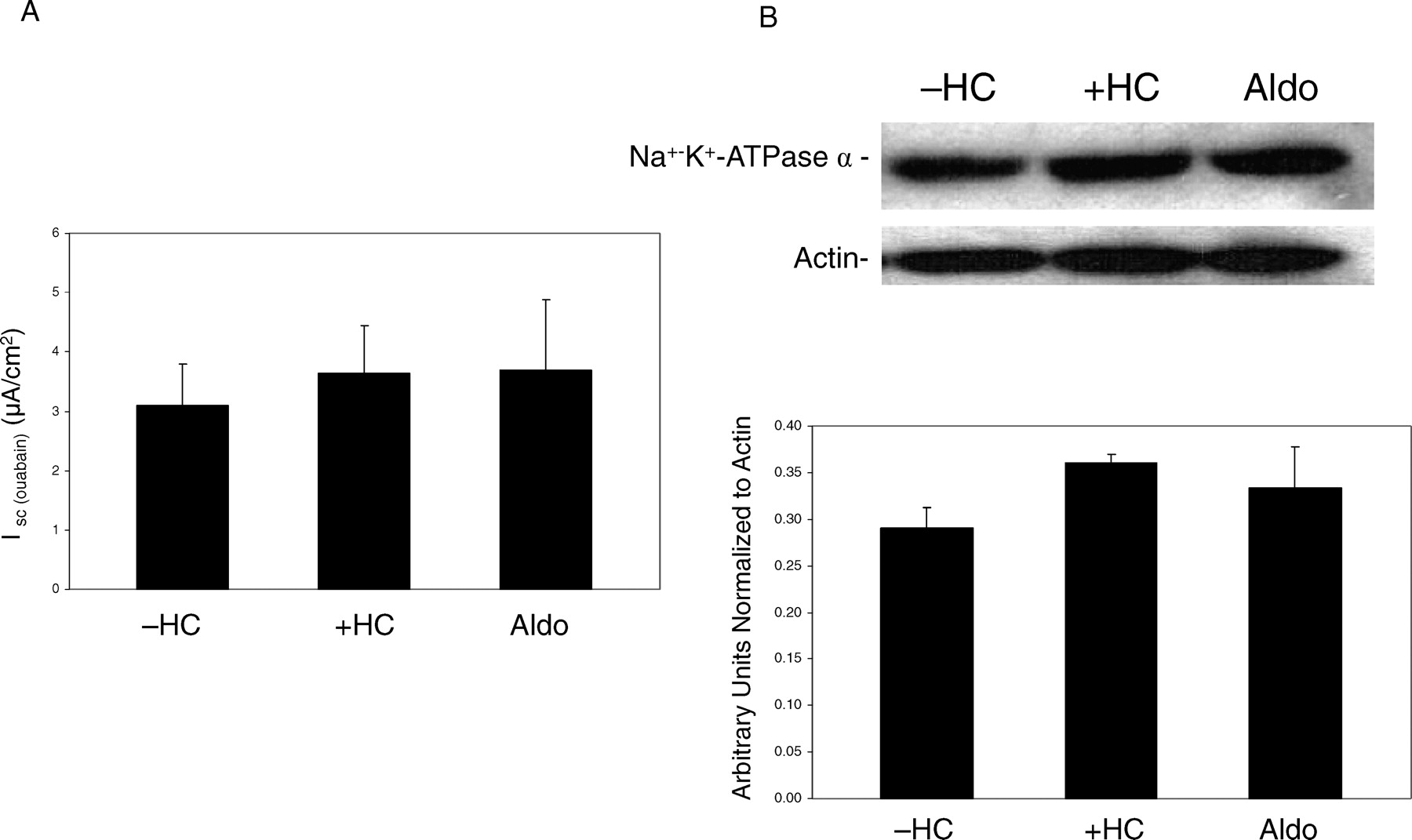
A) Effect of hydrocortisone (+HC) or aldosterone (+Aldo) on Na+-K+-ATPase-mediated current in permeabilized SMG-C6 monolayers. Values are means ± SE; n=6. B) Representative Western blot analysis of hydrocortisone (HC) or aldosterone (Aldo) effects on Na+-K+-ATPase protein in SMG-C6 cells (n = 5 for each treatment).
Footnotes
This work was supported by the Department of Pediatrics and the South Texas Health Research Center, University of Texas Health Science Center, San Antonio.
Acknowledgements
The authors acknowledge the technical assistance of Ms. Barbara Henson.