
Abstract
Mutations in hemojuvelin (HJV) cause severe juvenile hemochromatosis, characterized by iron loading of the heart, liver, and pancreas. Knockout (KO) mice lacking HJV (Hjv−/−) spontaneously load with dietary iron and, therefore, present a model for hereditary hemochromatosis (HH). In HH, iron chelation may be considered in noncandidates for phlebotomy. We examined the effects of deferasirox, an oral chelator, in Hjv−/− mice. Hepatic, cardiac, splenic, and pancreatic iron were determined by measuring elemental iron and scoring histological sections. Heart and liver iron levels were also determined repeatedly by quantitative R2* magnetic resonance imaging (MRI). The time course of iron loading without intervention was followed from Week 8 of age (study start) to Week 20, when once-daily (5×/ week) deferasirox was administered, to Week 28. At 8 weeks, liver iron of KO mice was already markedly elevated versus wild-type mice (P < 0.001) and reached a plateau around Week 14. In contrast, Week 8 cardiac and pancreatic iron levels were similar in both KO and wild-type mice and, compared with the liver, showed a delayed but massive iron loading up to Week 20. Contrary to the liver, heart, and pancreas, the KO mice spleen had lower iron content versus wild-type mice. In Hjv−/− mice, liver and heart iron burden was effectively reduced with deferasirox 100 mg/kg (P < 0.05). Although deferasirox was less efficacious at this dose in the pancreas, over the observed time period, a clear trend toward reduced organ iron load was noted. There was no noticeable effect of deferasirox upon splenic iron in Hjv−/− mice. Quantitative R2* MRI demonstrated the ability to assess iron concentrations in the liver and myocardial muscle accurately and repetitively. Hepatic (R = 0.86; P = 3.2*10− 12) and delayed myocardial (R = 0.81; P = 2.9*10− 10) iron accumulation could be followed noninvasively with high agreement to invasive methods.
Introduction
Genetic analysis of hemochromatosis has led to the discovery of a number of genes whose mutations disrupt iron homeostasis and lead to iron overload. Hepcidin, which is synthesized in the liver, has been identified as a key hormone for iron regulation (1). The plasma level of hepcidin reflects an integrated response to body iron load, anemia, hypoxia, and inflammation (reviewed in (2)). How exactly these body states are translated into a particular hepcidin level is a matter of intense research. At the molecular level, several candidate effectors that potentially serve as sensors for these body states have been identified including: transferrin receptor (TFR) 1, TFR 2, HFE, hemojuvelin (HJV), and IL-6 receptor.
Recent studies have shown that hepcidin regulates iron absorption by binding to ferroportin, the iron exporter of intestinal epithelial cells and reticuloendothelial macrophages. The interaction of hepcidin and ferroportin causes internalization of ferroportin and, therefore, abrogation of iron flux into the blood circulation (3, 4).
Homozygous disruption of the HFE gene by the C282Y mutation (5) is the major cause of hereditary hemochromatosis (HH), producing an inappropriately low expression of hepcidin (6) and leading to late-onset (dietary) iron overload. Two phenotypically similar forms of juvenile hemochromatosis (JH; or type 2 hemochromatosis) exist: one is due to mutations of the HJV gene on chromosome 1, which encodes the protein HJV (type 2A hemochromatosis), and the other is due to the mutation of the HAMP gene, encoding hepcidin on chromosome 19 (type 2B hemochromatosis). Both forms of JH result in very low hepcidin levels and secondary increased uptake of dietary iron. An important step toward the elucidation of the signaling pathway that leads from HJV to the expression of hepcidin has been made by the identification of bone morphogenetic protein (BMP) as a co-receptor of HJV, and that a HJV mutant associated with JH has impaired BMP signaling activity (7). However, the detailed role of HJV in iron homeostasis is still controversial and has yet to be fully established (8, 9).
In humans, iron overload in hemochromatosis is generally treated by phlebotomy. For example, in HFE-type hemochromatosis (after initial diagnosis of high iron burden), phlebotomies may be performed weekly for 6–24 months until iron stores normalize, as assessed by serum ferritin levels. Many patients then undergo periodic phlebotomies (usually 3–4 per year) to maintain acceptable levels of iron (10). Despite this generally safe and effective mode of iron removal, not all patients are candidates for phlebotomy due to underlying anemia, heart disease, or poor venous access, and compliance with regular phlebotomy may be an issue (11). Iron chelators are logical candidates to potentially replace or support a phlebotomy regimen. Indeed, the first demonstration that deferoxamine (Desferal, DFO) has the ability to mobilize and excrete body iron was in a hemochromatosis patient in 1961 (12). Due to its lack of oral bioavailability and short plasma half-life, DFO has to be given by slow sc or iv infusion and is not a realistic alternative to phlebotomy. Recently, deferasirox (Exjade), an orally active and long-acting tridentate iron chelator (13), has been approved for use in transfusional iron overload. Due to its properties, particularly its ability to mobilize liver iron, a clinical study evaluating the safety and efficacy of deferasirox in patients with C282Y HH and iron overload has been initiated (14).
In support of these studies, the effects of deferasirox have been assessed in HJV knockout (KO) mice. These mice, which lack the HJV orthologe (Hjv−/−), have been described recently (15, 16); the resulting mouse phenotype bears great similarity to JH in humans. These mice rapidly accumulated excess iron in the liver, heart, and pancreas, confirming that loss of HJV protein leads to iron overload.
The primary objectives of our study in Hjv−/− mice were to a) investigate the time course of organ iron loading and b) assess the iron-unloading effects of deferasirox on key organs including the liver, heart, spleen and pancreas. Inductively-coupled plasma optical emission spectrometry (ICP-OES) was utilized to assess organ iron levels post mortem. In addition, organ-iron deposition patterns were assessed histologically. Finally, hepatic and cardiac iron levels were measured in vivo by magnetic resonance imaging (MRI).
Materials and Methods
Statement on Animal Welfare.
All aspects of these studies were approved by the Veterinäramt Basel-Stadt and documented in the Tierversuchsbewilligung Nr 2099.
Animals.
Generation of Hjv−/− mice has recently been described (15). Rederivation was performed by transplantation of mutant heterozygous mouse embryos into C57BL/6 mice to obtain seven heterozygous (Hjv+/−) male mice. Crossing of these males with C57BL/6 mice and selection for heterozygous animals allowed Hjv+/−× Hjv+/− crossing and production of sufficient age-matched Hjv−/− and Hjv+/+ males for the study.
Maintenance Conditions.
Animals were kept individually in standard rodent cages (Type II Macrolon), had free access to water (HCl-acidified tap water; pH 2.5–2.9), and received standard rodent feed (Kliba/Nafag 3893).
Study Outline.
Age-matched male Hjv−/− and Hjv+/+mice entered the study at 8 weeks of age. The study duration was up to 28 weeks of age. Up to 20 weeks, organ iron was investigated in groups of animals to observe the time course of (spontaneous) iron loading. Between 20 and 28 weeks of age, groups of animals were treated daily with deferasirox or vehicle to observe unloading of organ iron. Mice were divided into two main subpopulations: one for the iron determination of organs by ICP-OES and MRI, and the other for histological organ assessment. Within the subpopulations, mice were randomized to groups of n = 4–7 and sacrificed for organ iron (OI) determination by ICP-OES and/or histology (H) at the time points depicted in Figure 1.
Preparation of Suspension for Administration of Test Compound.
The test compound (deferasirox, Exjade) was suspended in 0.5% aqueous Klucel HF (hydroxypropylcellulose) by ultrasonication. Each animal received 0.1 ml 0.5% aqueous Klucel (vehicle-treated animals) or 0.1 ml suspension per 10 g body weight (= 10 ml/kg) by gavage. Suspensions were kept at 4°C and made freshly every 2 weeks.
Treatment of Animals with Test Compound.
Deferasirox was given by oral gavage using a 1-ml plastic syringe equipped with a curved luer-lock canula.
Anesthesia.
For MRI measurements, animals were treated by inhalation anesthesia using 3%–5% isoflurane in O2/N2O 1:2 (v/v) mixture applied with a face mask at a rate of 0.7 L/min.
Perfusion of Animals.
If organs were removed for determination of organ iron, animals were perfused. For this procedure, after initial anesthesia, animals were equipped with a face mask to continuously inhale 1%–3% (volume) isoflurane. The thorax was opened and the left ventricle of the heart was pierced with a stainless steel needle, through which a sterile 0.9% NaCl solution was pumped at a flow rate of 200 ml/hr. As soon as the right auricle started swelling, it was opened and perfusion was stopped when the fluid was clear.
Collection of Organs for Iron Determination by ICP-OES.
Organs were removed by standard procedures, placed individually in 20-ml plastic scintillation vials, and dried for 16 hrs at 105°C in an oven that allowed air circulation.
ICP-OES.
Iron determination by ICP-OES was performed according to standard procedures in full accordance with the Pharm. Eur. Method 2.2.22 “Atomic Emission Spectrometry, Method I – Direct Calibration.”
Collection and Processing of Organs for Histology and Microscopic Evaluation.
Animals were sacrificed using CO2 and bled by cutting the vena cava. Organs were sampled by standard procedures, weighed, and stored in 4% formalin. Liver, heart, pancreas, and spleen were trimmed, embedded in Paraplast®, sectioned and stained with hematoxylin and eosin and the Prussian blue method for iron, subjected to microscopic examination, and the amount of positive staining for iron was determined semi-quantitatively using the following grading scheme: − no staining, (+) traces, + minimal, ++ slight, +++ moderate, ++++ marked.
MRI.
All experiments were performed on a Biospec DBX 47/30 spectrometer (Bruker Biospin, Ettlingen, Germany) at 4.7 T equipped with a self-shielded 12-cm bore gradient system capable of switching 200 mT/m in 170 μsec, Paravision software for data acquisition, and a 1H quadrature resonator with a diameter of 35 mm (Rapid Biomedical GmbH, Rimpar, Germany).
ECG Recording.
ECG was recorded with Bruker Physiogard SM 785 NMR (Bruker Biospin) using three platinum subdermal needle electrodes (Grass Instruments, West Warwick, USA) inserted into the two front paws and one hind paw. Normal heart rates of mice were approximately 550–650 bpm.
R2* Assessment of the Myocardium.
ECG-gated double oblique cardiac short axis gradient echo (GE) images were recorded at eight time points of the cardiac cycle beginning at the end-diastolic time point and separated by 22 ms. The image acquisition parameters were as follows: Time of repetition (TR) was given by the heart rate of the animal, but minimally 300 ms. Echo time (TE) = 2.8 ms, field of view (FOV) = 35 ×35 mm2, slice thickness 1.2 mm (single slice), number of averages (NA) = 4, matrix = 256 × 128 pixel (zero filled to 2562 pixel). Multiple single-echo images of the same position with TEs of 2.8, 5.6, 6.5, 7.5, 9, 12, and 15 ms were then acquired serially.
R2* Assessment of the Liver.
One transversal imaging plane was set to the center of the liver. R2*-weighted GE images were acquired by multiple single GE non-ECG gated images. The image acquisition parameters were as for cardiac images with TR = 200 ms, TEs = 2.8, 3.5, 4, 5.6, 9, and 15 ms.
R2* Computation.
A region-of-interest (ROI) of the left-ventricular myocardium consisting of the interventricular septum was drawn for R2* computation. Care was taken to exclude myocardial regions with major susceptibility artifacts adjacent to the lung. The signal time course was fitted to a mono-exponential function:
where S(t) is the signal at echo times TE, and A is the signal amplitude at zero TE.
Liver R2* was computed accordingly, but with an offset C added to algorithm (1):
Statistical Analysis.
Two-way ANOVA determined the influence of genetic background and time on iron accumulation. The statistical differences of pairwise-comparisons were determined using the Holm-Sidak test. Two-way repeated measure ANOVA with post hoc Holm-Sidak test was used to analyze MRI data where the time course was assessed in individual animals. One-way ANOVA was used to determine the differences in tissue iron content between various groups (Hjv−/− mice treated with 0, 30, or 100 mg/kg deferasirox and Hjv+/+ mice) using Holm-Sidak or Dunn’s tests. Statistical analyses were performed using SigmaStat 3.11 (Systat Software Inc.) with a significance level of P < 0.05. When required (and possible), data were transformed to provide normally distributed groups of equal variances. Tests for unequal variances were routinely performed during ANOVA analyses, but were further evaluated by using Bartlett’s F test (Systat 11.0, Systat Software Inc.). Linear regression was performed using SigmaPlot 9.01 (Systat Software Inc.) and Prism 3.0 (GraphPad Software Inc.) and nonlinear regression was performed using SigmaPlot 9.1. The statistical significance of mortality rates was determined by Fisher’s exact test (Hjv−/− vs Hjv+/+; SigmaStat 3.11) or Cochrane’s test for a linear trend (effect of 0, 30, or 100 mg/kg deferasirox on Hjv−/− mice only; Systat 11.0). No statistical analyses were performed for the results of histological evaluations.
Results
Body Weight Development of Hjv−/− and Hjv+/+ Animals.
The body weights of Hjv−/− and Hjv+/+ mice during the passive iron loading phase and during the deferasirox treatment phase are represented in Figure 2. The weight gains of the mice were initially similar, but both the rate of body weight gain and the final body weights were slightly affected in the Hjv−/− mice compared with the Hjv+/+ mice. Two-way ANOVA indicated a significant (P < 0.05) influence of time and mutation for animals ≥14 weeks of age (Fig. 2A). Deferasirox treatment apparently had little effect on the body weight of Hjv−/− mice, as the treatment groups did not differ significantly (Fig. 2B).
Spontaneous Iron Loading of Organs and Effects of Treatment with Deferasirox.
Liver: Liver iron concentration (LIC) was significantly (P < 0.001) higher at 8 and 28 weeks in the Hjv−/− mice compared with the Hjv+/+ mice (Fig. 3A). In the Hjv−/− mice, LIC was also significantly (P < 0.01) higher in the 14-week group compared with the 8-week group. LIC in the 14-, 20-, and 28-week groups was not statistically different, indicating that a plateau in iron loading was reached at around 14 weeks. Maximal LIC at Week 28 (Hjv−/− vehicle group) was approximately 21-fold higher than in the corresponding Hjv+/+ group. Histologically, in 8-week-old animals, depositions of iron-positive granules in hepatocytes of Hjv−/− mice were mainly in centrilobular or peripherolobular regions. By the age of 20 weeks, iron deposits were increased and located mainly in hepatocytes of peripherolobular areas and also in Kupffer cells (Table 1).
Deferasirox treatment, which started at Week 20 and ended at Week 28, significantly (P < 0.05) reduced LIC at the 100-mg/kg dose compared with the vehicle-treated group (Fig. 3A). Mean reductions of iron concentration compared with vehicle were 8.4% and 63.1% for the 30-and 100-mg/kg doses, respectively. Compared with Hjv+/+ mice, treatment of Hjv−/− mice with 100 mg/kg deferasirox for 8 weeks reduced the differences in liver iron levels from 21-fold to 8-fold (Fig. 2A). Histology sections indicated a further slight increase of iron deposition in vehicle-treated animals but with a similar distribution as in 20-week-old animals. Treatment with deferasirox at 30 and 100 mg/kg resulted in a decrease in iron-positive granules (Table 1, Fig. 4). In some animals, at the end of the study, deposits could only be found in Kupffer cells or macrophages, but no longer in hepatocytes. Due to the inter-individual variability and limited number of animals, no clear dose-dependent effect was noted in the sections.
Heart:
Visual inspection of the iron concentration versus time curve (Fig. 3B) suggests delayed iron loading of the heart compared with the liver. Notably, at Week 8, the differences in heart iron concentrations between Hjv−/− and Hjv+/+ mice are small. Maximal mean cardiac iron concentration at Week 28 (Hjv−/− vehicle group) was approximately 5-fold higher than in the corresponding Hjv+/+ group. By histology, no relevant iron deposits were observed in 8-week-old Hjv−/− animals. At 20 weeks of age, mild iron deposition was found mainly in cardiomyocytes. In 28-week-old mice treated with vehicle, a further slight increase of iron-positive granules in cardiomyocytes was noted (Table 1).
Deferasirox treatment significantly (P < 0.05) reduced cardiac iron concentration at the 100-mg/kg dose compared with the vehicle-treated group. Mean reductions of iron concentration compared with vehicle were 10.6% and 41.9% for the 30- and 100-mg/kg doses, respectively. Compared with Hjv+/+ mice, treatment of Hjv−/− mice with 100 mg/kg deferasirox for 8 weeks reduced the differences in heart iron levels from 5-fold to 2.9-fold. Histologically, iron-positive granules were somewhat reduced, with deferasirox at 30 and 100 mg/kg showing a slight dose dependency (Table 1, Fig. 4).
Pancreas:
Similar to the heart, delayed iron loading of the pancreas compared with the liver was observed (Fig. 3C). Pancreatic iron concentrations in Hjv+/+ mice were low over the observed time period, but maximal mean iron concentration at Week 28 (Hjv−/− vehicle group) was approximately 34-fold higher than in the corresponding Hjv+/+ group. Histologically, minimal focal iron-positive granules were observed in the pancreas in individual 8-week-old Hjv−/− animals. By the age of 20 weeks, slight iron deposits were seen in all Hjv−/− animals, predominantly in acinar cells of the exocrine pancreas and, to a lesser extent, in interstitial macrophages. The endocrine parts (pancreatic islets) appeared not to be affected. In 28-week-old mice treated with vehicle, there was a further slight increase in deposits showing similar distribution to 20-week-old animals (Table 1).
Treatment of animals with deferasirox showed no significant effects on iron levels. Still, a clear trend to lower iron concentrations was seen with the 100-mg/kg dose. Compared with Hjv+/+ mice, treatment of Hjv−/− mice with 100 mg/kg deferasirox for 8 weeks reduced the mean differences in pancreatic iron levels from 34-fold to approximately 21-fold. Histologically, iron-positive granules were minimally decreased in animals treated with deferasirox at 30 and 100 mg/kg (Table 1, Fig. 4).
Spleen:
Contrary to the other organs, the concentration time course (Fig. 3D) indicates that splenic iron in Hjv−/− mice is lower than in Hjv+/+ mice, although at Week 28, the difference in mean organ iron concentration was not statistically significant between Hjv−/− and Hjv+/+ mice. In histology sections, no relevant iron deposits were observed in 8-week-old animals. By the age of 20 weeks, minimal iron deposits were found, mainly in macrophages of the red pulp and in the white pulp. In 28-week-old mice treated with vehicle, the amount and distribution were comparable to those in 20-week-old animals. Treatment of mice with deferasirox for 8 weeks showed no effect on splenic iron when compared with the vehicle control as assessed by ICP-OES or histology.
In vivo Assessment of Hepatic and Cardiac Iron Content by Quantitative R2* MRI.
Nine age-matched groups of animals, three wild-type Hjv+/+ and six Hjv−/− groups, were investigated at several time points from 8 to 28 weeks of age (Fig. 1). During Weeks 8–20, natural iron loading was observed in vivo in the liver and heart by assessing R2* relaxation rate. Thereafter, the Hjv −/− animals were treated with either vehicle or deferasirox and in vivo assessment of R2* was continued.
Hepatic Iron:
Figure 5 shows T2*-weighted images of an individual animal taken on Week 20 when the iron concentration was maximal, Week 24, and Week 28, the end of the study. The animal was treated with 100 mg/kg deferasirox, and the reduction of the iron concentration in the liver can be seen by increased brightening over time.
Assessed R2* values were correlated post hoc with the iron concentrations as measured with ICP-OES. Pearson correlation (Fig. 6) revealed a strong association with R = 0.86 (P = 3.2*10−12). Linear regression between the parameters was used to calibrate R2*, allowing the assessment of in vivo hepatic iron concentration.
The hepatic iron time courses of the various groups are shown in Figure 7. Hepatic iron content in Hjv−/− animals increased steadily to approximately 7 mg Fe/g dry weight (dw) during the loading phase. Thereafter, treatment with 100 mg/kg deferasirox resulted in a rapid decrease. The value at the last time point must be viewed with caution, since only one animal was assessed due to several fatalities during MRI evaluation at this time point. Treatment with 30 mg/kg deferasirox resulted in a trend of dropping iron concentration, whereas iron levels in vehicle-treated animals reached a plateau and had a tendency to fall at the end of the study. The iron concentration time course was comparable to iron content as measured with ICP-OES (Fig. 3A).
Myocardial Iron:
T2*-weighted MR images of the heart show brightening during 8 weeks of deferasirox treatment (Fig. 8), though to a lesser extent than in liver images. Post hoc Pearson correlation of R2* versus myocardial iron content as measured by ICP-OES revealed a strong association (R = 0.81; P = 2.9*10−10). The linear regression coefficients were applied to calibrate R2* for myocardial iron concentration. Myocardial iron content time course revealed a delayed onset of iron accumulation in the myocardium (Fig. 9). No difference between Hjv+/+ and Hjv−/− could be discerned until Week 14. During the treatment phase, only myocardial iron in the group with the highest dose of 100 mg/kg tended to decrease. Due to the high fatality rate, only one animal was measured from this group at the latest time point. As the mean iron concentrations of the three groups were different at treatment onset, the relative myocardial iron concentration time course is shown in Figure 9B. The values were normalized to the pretreatment values of each group; thus, it became more apparent that the in vivo derived result was in line with post mortem iron determination (Fig. 3B).
Discussion
Several types of HH have been identified involving mutations of the HFE, HAMP, HJV, TFR2 and FPR1 genes, a common feature of which is increased iron absorption leading to iron overload. Disruption of HJV leads to a particularly severe juvenile form of HH that, in humans, leads to massive iron overload within the first 2 decades of life. Phlebotomy is the conventional treatment for HH, although iron chelation may be an alternative in patients who are not candidates for, or intolerant to, phlebotomy. Combined phlebotomy/chelation may be beneficial for severe cases of iron overload.
The RGMc gene (Hjv) is the mouse equivalent to the human HJV gene; therefore, organs of Hjv−/− mice become spontaneously iron loaded (15). To obtain supportive evidence for a beneficial effect of deferasirox in HH, the time course of iron loading of Hjv−/− versus the wild-type mice and the outcome of deferasirox treatment were investigated.
Body weight development in Hjv−/− mice was only marginally affected and a difference from Hjv+/+ mice became significant only after Week 14. Treatment with vehicle, 30 mg/kg deferasirox, or 100 mg/kg deferasirox for 8 weeks did not affect body weight.
Iron loading in Hjv−/− mice plateaued later in the heart and pancreas than in the liver, indicating delayed iron deposition; this is consistent with the liver being the primary iron storage organ. R2* and myocardial iron concentration measured by ICP-OES were almost perfectly correlated and the various groups had distinct differences in iron overload at the beginning of deferasirox treatment. The concentration ranges for myocardial iron were similar for both techniques (0.4 mg Fe/g dw at Week 8 to 1.2 mg Fe/g dw at Week 20). Administration of 100 mg/kg deferasirox for 8 weeks significantly decreased liver and cardiac iron, confirming that deferasirox has access to both organs. However, the last cardiac MRI data point has to be interpreted with caution, since only one animal was assessed. Nevertheless, individual R2* assessments revealed a solid decrease in iron content already at Week 24 in the hearts of five out of seven animals. Of the other organs investigated, the pancreas of Hjv−/− mice became most severely iron loaded, compared with wild-type mice; deferasirox 100 mg/kg showed a trend toward decreasing iron burden in the pancreas.
No statistically significant effects of 30 mg/kg deferasirox were observed. Although this may seem contradictory to human data (17), it may be explained by the lower exposure of deferasirox in small animals and the fact that the animals were treated for a relatively short period of time.
Interestingly, the data suggested that iron metabolism in wild-type animals is more tightly controlled (as reflected by low variance) than in mutant groups, and that deferasirox does not overcome variability in iron load caused by the mutation.
A good correlation was found between iron determinations by ICP-OES and histological assessment. Apart from the iron deposits, no relevant morphological alterations were observed in the organs evaluated histopathologically in untreated Hjv−/− mice or in animals treated with vehicle or deferasirox. In young Hjv−/− mice, iron was first deposited in liver hepatocytes and, with increasing iron load, in Kupffer cells. Hepatocytes of peripherolobular areas were more consistently affected, which is in line with previously reported data (1, 16); deferasirox appears to reduce iron load in hepatocytes more readily than in Kupffer cells. Prominent iron deposits in iron-loaded mice were also seen in the exocrine pancreas, but not in the pancreatic islets, which also agrees with other published data (1, 18); deferasirox minimally reduced stainable iron deposits. Iron deposits in the heart were less pronounced than in the liver or pancreas, but were decreased by deferasirox. As for USF2 KO mice (1), iron deposition was very low in the spleen of Hjv−/− mice compared with wild-type animals, where an age-dependent increase in stainable iron deposits was observed. Low splenic iron in Hjv−/− mice is consistent with the notion of ‘uncontrolled’ iron release due to lack of hepcidin, which enhances ferroportin expression on the surface of splenic macrophages (16).
In humans, MRI enables non-invasive and repeated assessment of organ iron content over time, since R2* relaxation rate is directly correlated to tissue iron concentration (19–21). In the present study, hepatic R2* was significantly correlated with iron concentration. The variability observed (Fig. 7) may have various causes, including misregistration of the multiple echo images due to respiration, inclusion of large blood vessels, inhomogeneous liver iron distribution (R2* was measured from one slice whereas [Fe] was assessed in the entire organ), or variable iron storage crystals with varying paramagnetic properties. A strong linear correlation also exists between R2* and liver or myocardial iron in gerbils (19). After retrospectively applying the obtained correlation coefficients, the iron concentration of wild-type mice was in agreement with approximately 0.21 mg Fe/kg dw and 0.25 mg Fe/kg dw for MRI and ICP-OES, respectively. The overall liver iron time courses derived from MRI and ICP-OES are similar, though MRI had a larger dynamic range. It should be noted that the ICP-OES assessments were performed on animal groups sacrificed at the various time points, whereas MRI was performed on the same animal throughout the study. Furthermore, the relaxivity of hepatic iron may depend on the crystal type or size and the relaxivity difference of iron oxide–containing contrast agents with diverse iron core sizes between 4 and 11 nm (data not shown) indicated that measurement of small crystals underestimated the amount of iron present. This may partly explain the discrepancy in liver iron levels between MRI and ICP-OES at 8 weeks of age, as at that early time point the deposited iron could be clustered in smaller crystals.
In conclusion, this study confirmed that Hjv−/− mice serve as a model for hemochromatosis with severe iron overload. Iron loading of the heart and pancreas was delayed versus the liver, which already had high iron content in 8-week-old mice. Deferasirox effectively reduced liver and heart iron of Hjv−/− mice and showed a trend to reduction of iron in the pancreas. In vivo MRI R2* techniques have shown accurate iron concentration assessments in rat liver and heart myocardium.
Histology: Summary of Iron Deposits in Various Organs of Hjv−/− and Hjv+/+ Mice a

Study outline. Hjv−/− and Hjv+/+ mice were age-matched and were fed a regular diet throughout the whole study. Groups of animals were sacrificed for organ removal at the time points indicated by the tips of the arrows. For example, at 8 weeks of age, six Hjv−/− animals were sacrificed for organ iron (OI) determination and six animals for organ histology (H). At Week 20, three groups of Hjv−/− animals were treated 5 days a week either with vehicle, 30 mg/kg deferasirox, or 100 mg/kg deferasirox for 8 weeks. Hjv+/+ animals were not treated with deferasirox. The black diamonds indicate time points (8, 14, 20, 24, and 28 weeks) at which MRI studies were performed.
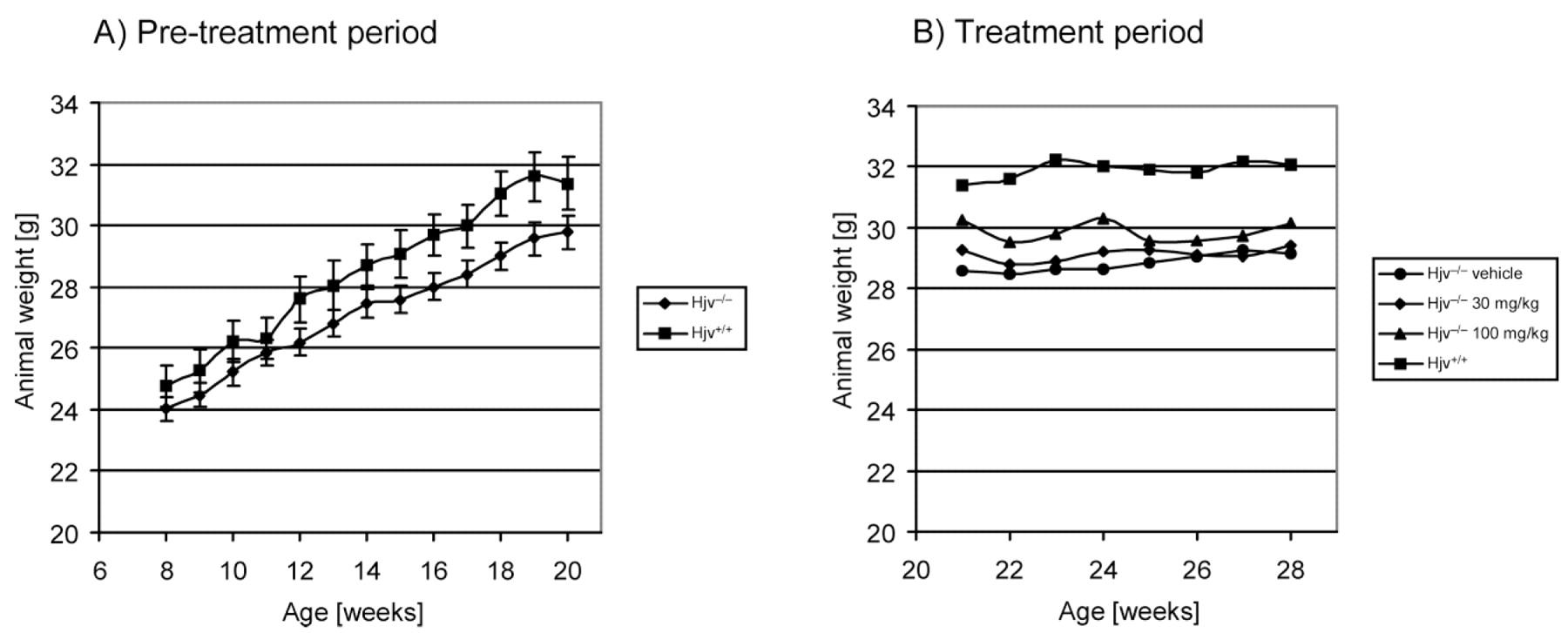
Body weights of Hjv−/− and Hjv+/+ mice were recorded periodically during the passive iron acquisition phase (Panel A) and the deferasirox treatment phase (Panel B). Two-way ANOVA was used to determine the effect of mutation and time on body weight gains in Panel A. Significant differences (P < 0.05) between the groups were calculated for animals aged 14 weeks and older. Body weights between the treatment groups were not statistically different (panel B).
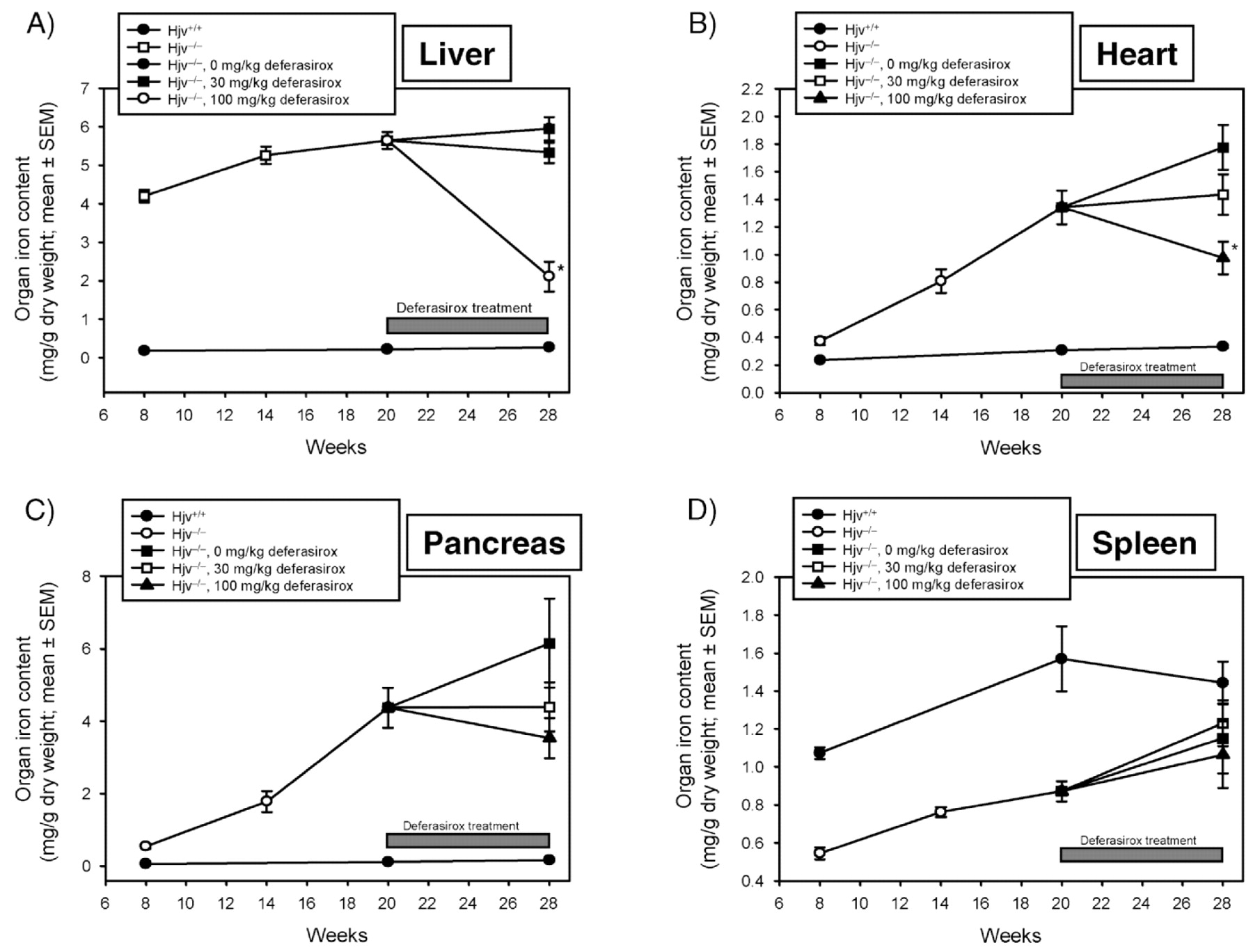
Organ iron as determined by ICP-OES.
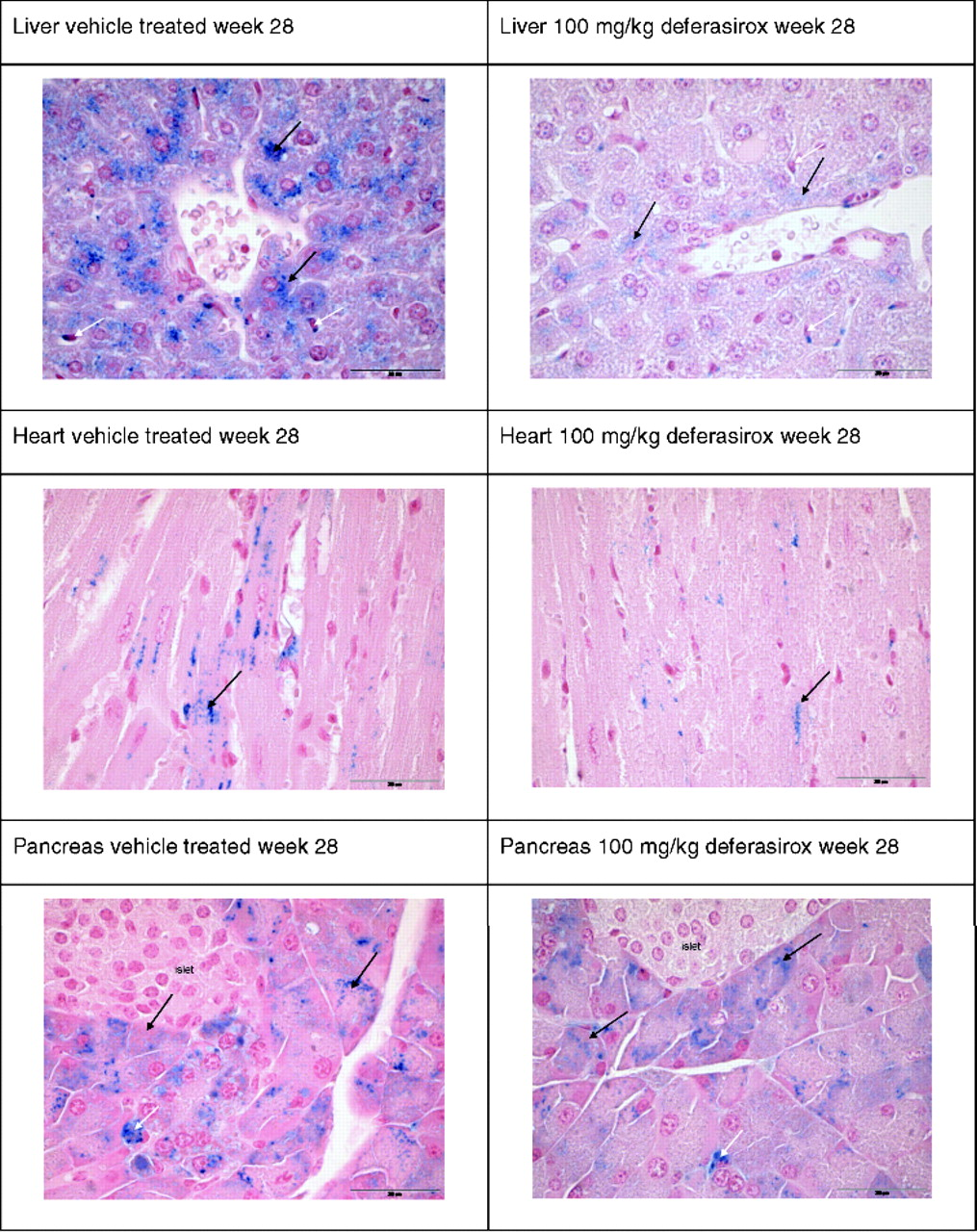
Histology of liver heart and pancreas at Week 28. Liver: vehicle treated, iron deposits in hepatocytes (black arrow), and Kupffer cells (white arrows); deferasirox treated, substantial reduction of iron deposits in hepatocytes (black arrows) and less pronounced in Kupffer cells (white arrows). Heart: Vehicle treated, iron deposits in cardiomyocytes (black arrows); deferasirox treated, reduction of iron deposits in cardiomyocytes (black arrow). Exocrine pancreas: vehicle treated, iron deposits in acinar cells (black arrows) and interstitial macrophages (white arrow); deferasirox treated, slight reduction of iron deposits in acinar cells (black arrows); interstitial macrophage (white arrow).

T2*-weighted liver MR images of a Hjv–/– animal at 20 (A), 24 (B), and 28 (C) weeks of age. The animal was treated with 100 mg/kg deferasirox po daily during this time period. Mean R2* values of the liver were 553.2 s−1, 244.2 s−1, and 43.6 s−1 at 20, 24, and 28 weeks, respectively.

Pearson product moment correlation of hepatic iron concentration with R2*. Linear regression was performed for calibration of R2* with the iron concentration.


T2*-weighted heart MR images of a Hjv−/− animal at 20 (A), 24 (B), and 28 (C) weeks of age. The animal was treated with 100 mg/kg deferasirox during this time period. R2* values of the myocardium were 164.0 s−1, 128.0 s−1, and 35.6 s−1 at 20, 24, and 28 weeks, respectively.

(A) Myocardial iron concentration time course (means ± SEM) as measured in vivo by MRI. Treatment with either vehicle or deferasirox was initiated at Week 20 and continued to the end of the study at Week 28. (B) Relative iron concentration changes of the three groups during the treatment period.
Footnotes
2
Current address: SAN GmbH, Rieden, Switzerland.
3
Current address: Institute of Zoology, University of Zürich, Switzerland.