
Abstract
Several microbial species, including probiotic lactic acid bacteria, have the ability to irreversibly bind a large variety of polyphenols (flavonoids) and anthocyanidins found in many colored fruits and vegetables and to enhance their total oxidant-scavenging capacities (TOSC). The binding of flavonoids to microbial surfaces was further increased by the cationic polyelectrolytes ligands poly-L-histidine, chlorhexidine and Copaxone®. This phenomenon was confirmed visually, by the FRAP, DPPH, cyclic voltammetry, Folin-Ciocalteu as well as by luminol-dependent chemiluminescence techniques employed to assay TOSC. The possibility is considered that clinically, microbial cells in the oral cavity and in the gastro intestinal tract, complexed with antioxidant polyphenols from nutrients and with cationic ligands, might increase the protection of mammalian cells against damage induced by excessive generation of reactive oxygen species during infections and inflammation.
Introduction
Oxidative stress plays a central role in the pathophysiology of many clinical disorders (1–9). Several epidemiological studies conducted recently have indicated that dietary consumption of nutrients and beverages, rich in antioxidant bioflavonoids found in large amounts in colored fruits and vegetables, in red wine, green tea and in coffee, might decrease the risks of various disorders characterized by oxidative stress (10, 11). It has also been argued that irrespective of the amounts of low molecular weight antioxidants (LMWA) as well as of flavonoids consumed, the total oxidant-scavenging capacities (TOSC) of human body fluids, organs and of several cells in culture (12), could not be easily altered, due probably to strict regulatory mechanisms. However, the possibility that the TOSC of bacterial cells, present in large numbers in the oral cavity and the gastro intestinal tract, might be enhanced by attaching upon their surfaces flavonoids and also flavonoids complexed with cationic peptides, which act as opsonic ligands (13), has not been investigated. These issues are the basis for the current presentation, which will also discuss the possible beneficial effects of bacteria-polyphenol-polycation complexes as potential protectors against oxidative stress.
Materials and Methods
Polyphenols and Extracts from Plants.
Unless otherwise indicated, all reagents and materials used in this work were obtained from Sigma-Aldrich (St. Louis, MO, USA). Quercetin, catechin, gallic acid, tannic acid, morin, rutin, caffeic acid, epigallocatechin gallate (EGCG), curcumin, dopamine and nor-dihydroguiaretic acid (NDGA) were first dissolved in dimethyl sulfoxide (DMSO), at 100 mM and then further diluted by absolute ethanol/water to prepare diluted stock solutions at 1–10 mM. Due to the low solubility of quercetin in aqueous solutions, quercetin at 10 mM dissolved in DMSO was prepared in boiling water. One hour later, the insoluble quercetin was removed by high speed centrifugation and the completely soluble quercetin solution remaining was then passed through 0.45 μm filter (Schleicher & Schull). The amount of quercetin remaining soluble was approximately 500 μm as determined by high performance liquid chromatography using U.V. detector at 254 nm. Red wine, cranberry juice and green tea leaves (supernatant fluids derived from mixing 100 mg leaves with 10 ml boiling water) were purchased from a local supermarket.
Microbial Cells.
One hundred mg of a lyophilized preparation of Micrococcus lysodeikticus (M.L.) was suspended in 1 ml of Hanks’ balanced salt solution (HBSS) (Biological Industries, Kibbutz Beit Haemek, Israel) and adjusted to an optical density of ~30 at 550 nm (approx. 1 × 109 cells).
A commercial lyophilized mixture of 8 microbial probiotic species (VSL#3) was obtained from Seaford Pharmaceuticals Inc., USA. It contained a mixture of four species of Lactobacilli: 3.1% (w/w) L. plantarum, 7.3% L. acidophilus, 16% L. casei and 8.4% of L. delbrueckii subsp. Bulgaricus; three species of Bifidobacteria: B. infantis, B. breve and B. longum representing 17.7% of the mixture; and 47.5% of Streptococcus salivarius subsp. thermofilus. For all experiments, 100 mg of the probiotic formula powder were washed twice in a final volume of 20 ml of normal saline to remove excipients, suspended in 1 ml of normal saline (0.9% NaCl) and adjusted to an absorbancy of approx. 30OD550 (~108–109 cfu/ml).
Clinical isolates of non-MRSA (Methicillin-resistant Staphylococcus aureus) and Escherichia coli were obtained from the Clinical Microbiology Department, Hadassah Hospital, Jerusalem, Israel. The microbial cells were grown at 37°C for 14 hours on surfaces of brain heart infusion agar plates, and the colonies were scraped off, washed 3 times in a final volume of 20 ml of normal saline and adjusted to a density of 30OD550 (~108–109 cfu/ml).
Anaerobic microbial flora from deep gingival pockets from 6 patients with periodontal disease, obtained by consent, was cultivated at 37°C separately for 18 hours on blood agar plates in anaerobic jars. The colonies were scraped off, pulled together and washed three times in a final volume of 30 ml of normal saline. Bacterial suspensions were adjusted spectrophotometrically to a density of 30OD550 (~108–109 cfu/ml).
All the cell suspensions were kept on crushed ice and used to form complexes with polyphenols, fruit extracts and with cationic peptides (see below).
Coating Bacterial Cells by Polyphenols and by Cationic Peptides.
Bacterial cells (200 μL) were added to 0.8 ml of normal saline. The cells were then mixed either with 0.05–1 mM of various polyphenols or with 100 μL/ml amounts of red wine, cranberry juice, and green tea beverages. Bacterial suspensions were also treated simultaneously with polyphenols, plant extracts and with the cationic agents, poly L-histidine (M.W. 13000) (10–50 μM) and with 100–250 μg/ml of Copaxone® (glatiramer acetate), a polymer comprised of L-glutamic acid, L-lysine, L-alanine and L-tyrosine (Teva Pharmaceutical Industries, Ltd, Petah Tiqva, Israel). The reaction mixtures were kept for 10 minutes at room temperature, washed 3 times with a final volume of 30 ml of normal saline to remove unbound material, and finally re-suspended in 800 μL of HBSS. Various amounts of the complexes formed, were then tested for their TOSC, employing 4 different methods (see below).
Since most of the flavonoids employed in this study were first dissolved in DMSO or ethanol, and then diluted in HBSS, the amounts of solvents in the test samples (0.01%) did not interfere either with the assays employed or with the coating of bacteria by flavonoids and by the cationic agents.
Measurements of Polyphenols Absorption by Bacterial Cells.
The total polyphenolic content absorbed upon VSL#3 preparation was determined according to the modified Folin-Ciocalteu method and expressed as gallic acid equivalents (GAE) (14). Briefly, to 1 ml normal saline, were added 100 μL washed bacterial suspensions, complexed with polyphenol, followed by the addition of 50 μL of Folin reagent and immediately by 150 μL of a saturated sodium carbonate solution (25% w/v). After 10 minutes at room temperature the O.D. was determined at 760 nm.
Determinations of TOSC by Various Methods.
Luminol-Dependent Chemiluminescence (LDCL).
A glucose oxidase (GO) cocktail (15, 16) was used as a generator of reactive oxygen species. It is comprised of combinations among, 0.9 mL HBSS containing 1 mg/ml D-glucose, luminol (10 μM), glucose oxidase (GO) (2.3 units), sodium selenite (IV) (2 mM) and of CoCl2·4H2O (10 μM). This cocktail generates an intense and steady peak of luminescence due to the formation of hydrogen peroxide (H2O2) and hydroxyl radical (17). LDCL was measured by a Lumac 2500 M Biocounter (Landgraaf, The Netherlands) and expressed as counts per minute (cpm). The TOSC of the various agents tested is expressed as their ability to quench the luminescence generated by the cocktail due probably to both their antioxidant properties and their chelating activities for cobalt (II) (see Results).
Ferric Ion Reducing Antioxidant Power (FRAP) Assay.
The FRAP assay which uses antioxidants as reductants in a redox-linked colorimetric test was employed according to the modified method of Benzie and Strain (18). Briefly, a 300 mM acetate buffer at pH 3.6, 10 mM 2,4,6-tri-(2-pyridyl)-1,3,5-triazine, 98% (TPTZ; Sigma-Aldrich) (3.1 mg/ml in 40 mM HCl) and 20 mM FeCl3·6H2O (5.4 mg/ml in distilled H2O) were mixed together in a ratio of 10:1:1, respectively, to yield the working FRAP reagent. 10–40 μL aliquots of different agents were added to 300 μL of FRAP reagent. A total of 150 μL of the mixtures was applied to a 96-well plate. After 10 minutes of incubation, absorbance at a wavelength of 595 nm was measured with Biotek PowerWave 340 micro plate scanning spectrophotometer (Biotek Instruments, Winooski, VT). Calculations were made on the basis of standard curves obtained for a gallic acid solution and presented as gallic acid equivalent (GAE). All measurements were taken at room temperature with samples protected from direct light.
DPPH Assay.
The DPPH assay (19) measures hydrogen atom (or one electron) donating activity and hence provides an evaluation of antioxidant activity due to free radical scavenging. 2,2-Diphenyl-picrylhydrazyl radical (DPPH), a purple-colored stable free radical, is reduced to a yellow-colored diphenylpicryl hydrazine. A modified spectrophotometric assay was carried out by the following procedure: 50 μL of a sample solution containing the tested compound, e.g. bacterial cells or polyphenols-bacteria complexes were added to 800 μL of absolute methanol solution. Ten μL of DPPH (from 1 mM stock) were then added. Following 10 minutes of incubation at room temperature and protected from light, the absorbance at 515 nm was determined using a Cecil CE 1011 spectrophotometer (Cecil, London, UK). Calculations were made on the basis of standard curves obtained for a gallic acid solution and presented as gallic acid equivalents (GAE).
Cyclic Voltammetry (CV) and Differential Pulse Voltammetry (DPV) Methods.
Two voltammetry methods were used to measure the reducing power of bacteria coated by various polyphenols. The rationale for using the voltammetry methods for evaluating total polyphenols binding to bacteria was that all the low molecular weight antioxidants (LMWA) (scavengers) are reducing equivalents and thus all contribute to the total reducing power of the biological sample tested (20). A BAS model CV-1B cyclic voltammeter (West Lafayette, IN, USA) was used. CV tracings were recorded at a range of −0.3 V to 1.0 V and a rate of 100 mV/s vs. an Ag/AgCl reference electrode. The sensitivity of the cyclic voltammeter was set to 1e−5 (A/V). A three electrode system was used throughout the study. The working electrode was a glassy carbon disk (BAS MF-2012) of 3.2 mm diameter. A platinum wire served as the counter-auxiliary electrode. The working electrode was polished prior to each measurement with polishing kit (BAS-PK-1). CV tracings were analyzed to determine peak potential (E1/2) and anodic current (Ia). This potential is typical for specific tissues or compounds, and represents the ability of the samples reducing ability equivalents, representing the anodic wave capacity to donate electrons to the working electrode. The Ia correlated with the concentration of the reducing equivalents. While by using DPV, the current is measured at different times of the duration of one pulse, CV measures the potential, which is increased linearly to maximum and then decreases with the same gradient. On the other hand, in DPV measurements the effect of the charging current is minimized, so higher sensitivity can be achieved than in CV measurements.
Statistical Analysis.
Statistical analysis was performed using GraphPad Prism (GraphPad Software, San Diego, CA). Results are presented as the mean ± SD. Unpaired Student’s t test was performed as specified in figure/table legends (differences were considered significant when P < 0.05).
Results
Use of the GO Cocktail to Measure TOSC of Polyphenols.
Figure 1 shows that randomly selected polyphenols at 10 μM (equivalent to 4–12 μM gallic acid tested by the Folin-Ciocalteu phenol reagent), wine and a cranberry beverages at 10 μL (12–18 μM gallic acid equivalents), expressed different luminescence inhibitory patterns. The strongest inhibition of luminescence in a descending order was by wine, cranberry beverage, quercetin, gallic acid, curcumin, catechin, caffeic acid, tannic acid, dopamine and nor-dihydroguiaretic acid (NDGA). The data indicate that different polyphenols possess different antioxidant capabilities due either to their antioxidant properties or to their capacities to chelate cobalt (see the discussion section).
Polyphenols Enhance TOSC of a Probiotic Mixture (VSL#3).
The possibility that polyphenols bound to bacterial surfaces could exhibit enhanced TOSC was analyzed. Bacterial cells, washed in normal saline, were incubated with a variety of polyphenols to form complexes, washed to remove unbound materials and then tested for their oxidant scavenging capacities using the chemiluminescence, DPPH, FRAP and the cyclic voltammetry assays. By comparison, the antioxidant capacities of complexes formed between bacteria and red wine/various beverages, rich in polyphenols content, were also analyzed.
Use of the Glucose Oxidase Chemiluminescence Assay.
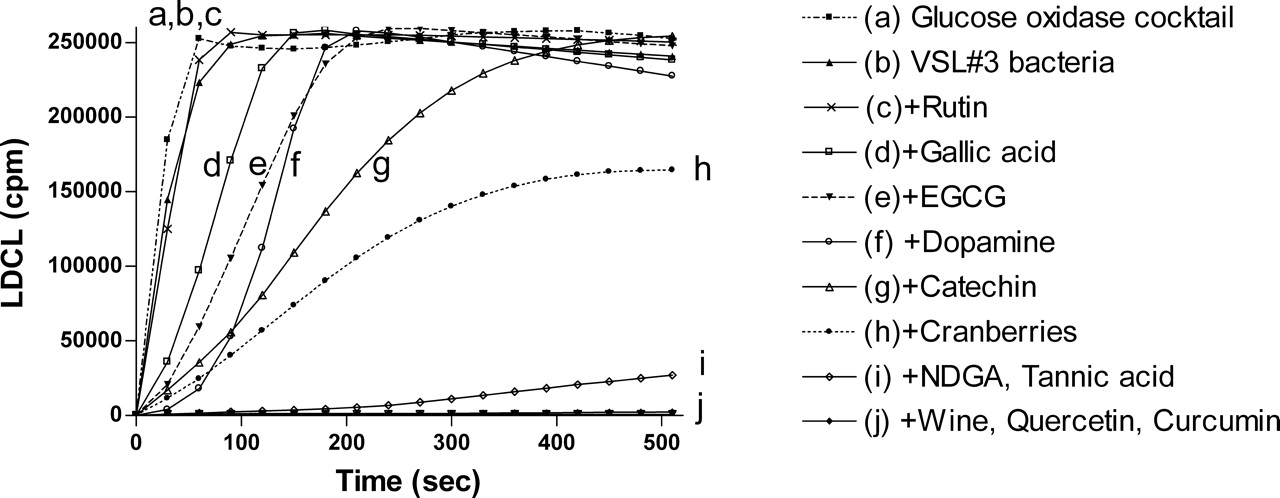
Figure 2 shows the LDCL inhibition patterns induced by VSL#3 pre-coated by 1 mM amounts of a variety of polyphenols and with 100 μL amounts of red wine and a cranberry beverage. While at the density used, bacteria alone had only very low antioxidative effects, stronger effects were induced by bacteria complexed with curcumin, quercetin, red wine, NDGA and by tannic acid. Cranberries and catechin had intermediate effects, and VSL#3 cells pre-coated by the other polyphenols had lesser antioxidant effects. The data in Figure 2 suggest that using the luminescence method not all polyphenols used at 1 mM to coat the bacterial cells, had equal binding affinities and antioxidant capacities.
The DPPH Assay.
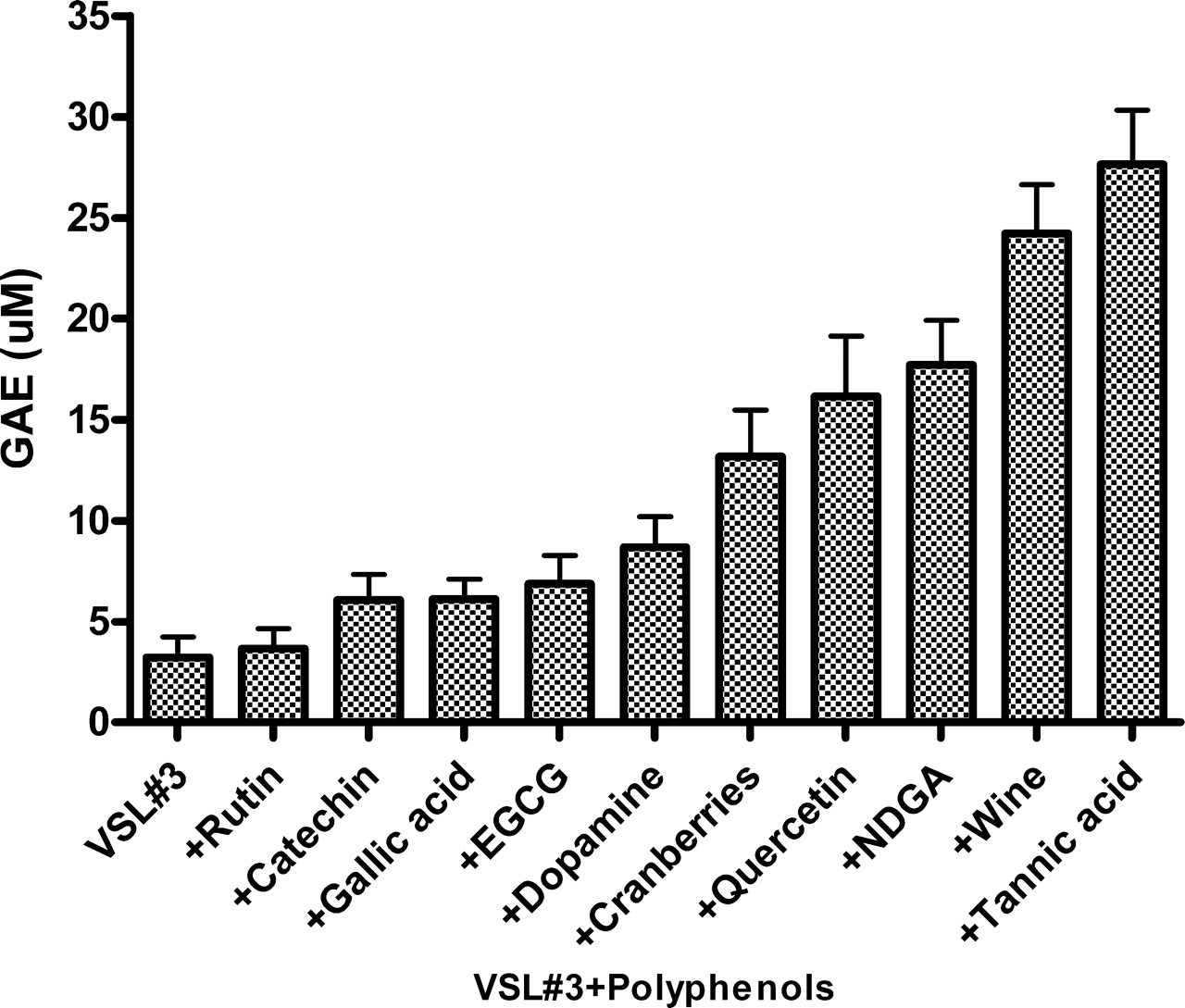
The DPPH assay measures hydrogen atom (or one electron) donating activity and hence provides means to evaluate antioxidant activity due to free radical scavenging. Expressed as gallic acid equivalents (GAE), Figure 3 shows that using this assay, while VSL#3 bacterial mixture, at the density used, showed only minimal antioxidant activity, cells pre-coated by 1 mM amounts of tannic acid (~55 μM GAE), had the strongest oxidant scavenging capacities. This was followed in descending order by the other agent used to pre-coat the bacteria.
The FRAP Assay.
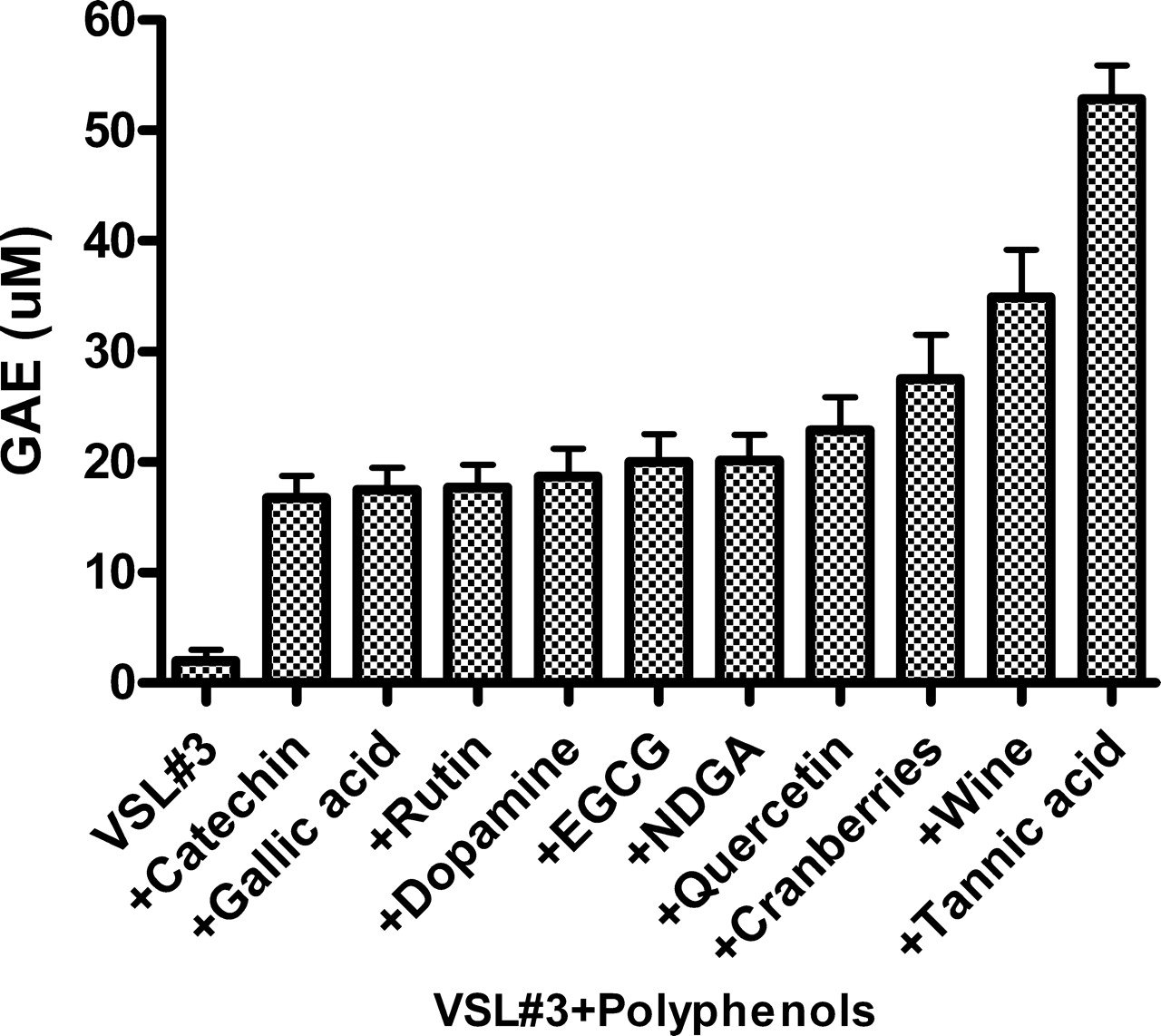
The FRAP assay uses antioxidants as reductants in a redox-linked colorimetric test. Expressed as gallic acid equivalents (GAE), Figure 4 shows that similar patterns of inhibition were obtained by the same polyphenols (see Fig. 3). While VSL#3 coated by 1 mM tannic acid possessed the most potent effect (~28 μM GAE), red wine at 100 μL was nearly as effective as tannic acid at this concentration (~25 μM GAE). The data in Figures 2–4 suggest that binding of certain polyphenols to bacterial cells significantly increased their TOSC.
The Folin-Ciocalteu’s Polyphenol Assay.
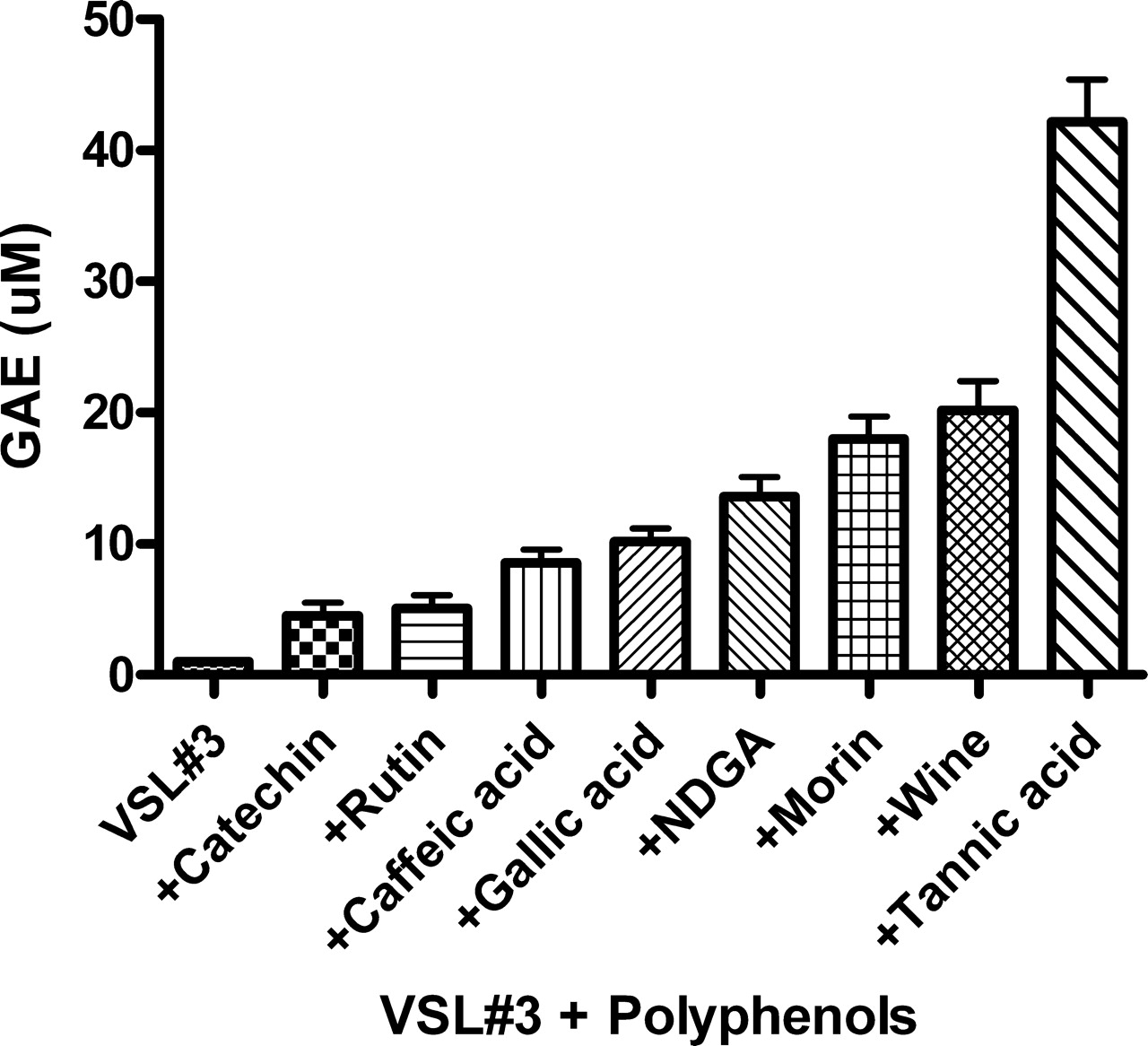
To investigate the extent of binding of various flavonoids to the VSL#3 bacterial surfaces, we used the Folin-Ciocalteu’s phenol reagent. Bacterial cells were first incubated with 500 μM of the various polyphenols, washed to remove unbound material and then interacted with the Folin reagent (see Materials and Methods). Figure 5 shows that tannic acid displayed the most potent gallic acid equivalents followed in a descending order by the other flavonoids tested. As can be seen, different polyphenols have different abilities to bind to the bacterial surfaces as expressed by gallic acid equivalents (GAE). This absorption of flavonoids to bacterial surfaces was dose-dependent (data not shown) and was also demonstrated using tannic acid as a representative flavonoid by cyclic voltammetry assay (see Fig. 6 below).
The Cyclic Voltammetry Assay.
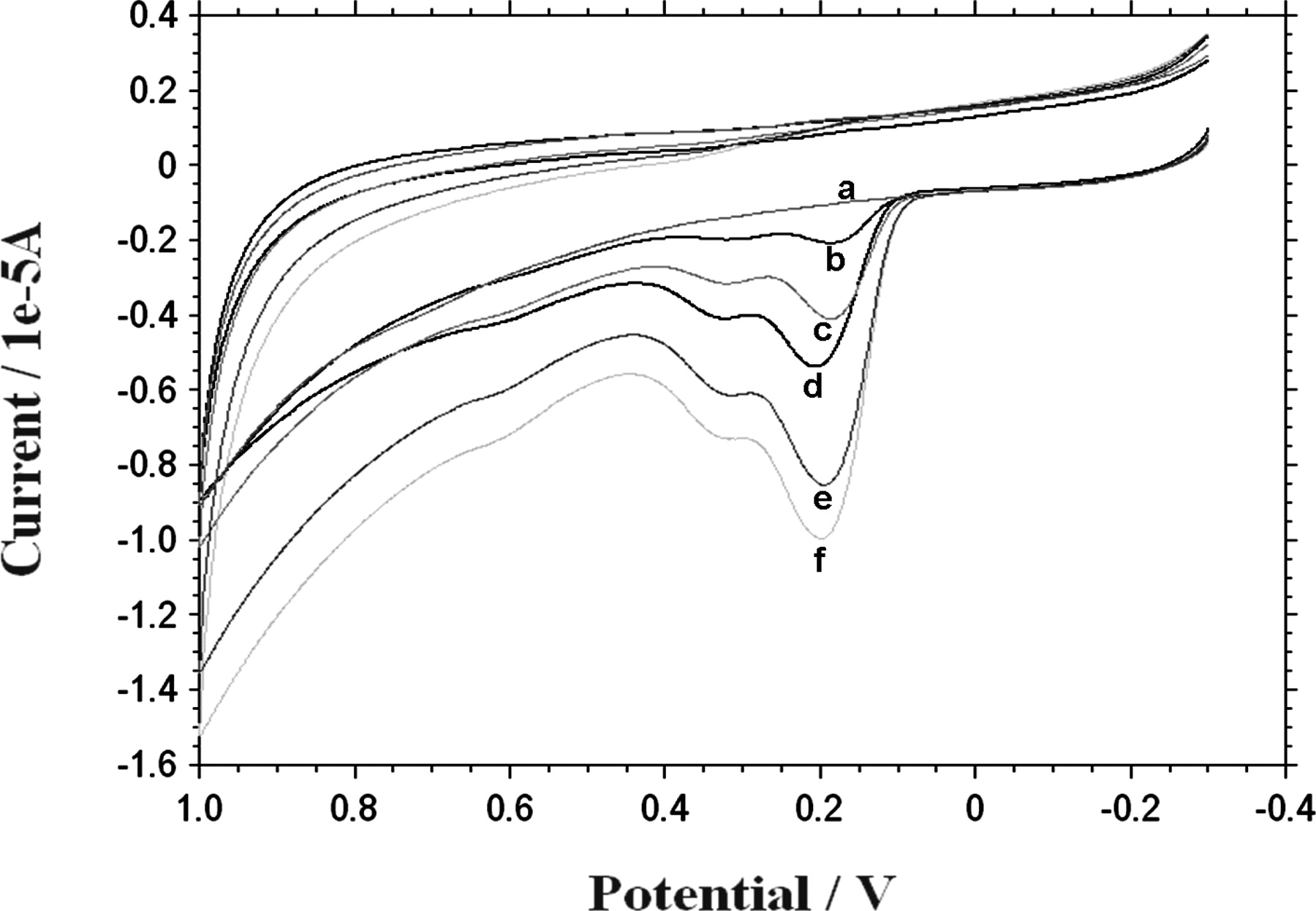
Polyphenolic compounds contain hydroxyl groups at the ring structures that can be electrochemically oxidized. Both flavonoids and phenolic acids exhibit one or two oxidation peaks (anodic waves), at about 0.2–0.3 V and 0.5–0.7 V, respectively. Oxidation peaks are associated with one or two oxidation centers present in polyphenolic molecules (21). Cyclic voltammetry (CV) experiments with VSL#3 were performed to test their ability to present a typical tannic acid voltammogram peaks following incubation with this polyphenol. VSL#3 preparation suspended in 1 ml electrolytes-rich HBSS were incubated for 10 minutes with tannic acid (0.1–1 mM) and washed 3 times to remove unbound polyphenol. Figure 6 shows that while under the experimental conditions, VSL#3 alone had no anodic wave (upper curve), tannic acid alone at 100 μM demonstrated a typical voltammogram (lowermost curve), showing 3 major oxidation anodic waves (at 0.22, 0.35 and 0.63 Volts). When the 0.22 V anodic wave was analyzed, it was also found that VSL#3 complexed with tannic acid at 0.1–1 mM showed dose-dependent voltammogram patterns, indicating that tannic acid was indeed bound to the bacterial cells. We verified the same patterns also with additional polyphenols and with other bacterial strains (See Fig. 9).
VSL#3 Bacterial Preparation Stained by Colored Polyphenols and Fruit Beverages.
An additional proof that polyphenols interact strongly and irreversibly with the VSL#3 preparation is presented in Figure 7. As can be seen (upper line test tubes), bacteria treated by the colored polyphenols morin and catechin, and by red wine, tea and polyphenon-60 (a 60% catechin preparation), retained typical colors. This phenomenon is even better demonstrated in the lower line of tubes where the bacterial suspensions were sedimented following centrifugation. It is also shown that VSL#3 complexed with the bile salts bilirubin and biliverdin (a bilirubin precursor) also retained a characteristic yellow/green color. It was also found (not shown) that a large variety of aerobic Gram positive and Gram negative bacteria as well as anaerobic bacteria obtained from a deep gingival crevice, also had the ability to strongly bind colored polyphenols to their surfaces.
Polyphenols Enhance TOSC of Other Bacterial Strains.
The ability of polyphenols to modify TOSC of a variety of additional bacterial species is shown in Figures 8 and 9 using the FRAP and the voltammetry methods, respectively (see Materials and Methods). Employing the FRAP assay, Figure 8 shows the enhancement of TOSC of Staphylococcus aureus and Micrococcus lysodeikticus (M.L.) induced by the polyphenols EGCG, tannic acid, curcumin (at 1 mM) and by red wine (at 100 μL). It indicates that for these bacterial strains tested, the highest antioxidant capacities can be endowed by wine, curcumin and by tannic acid.
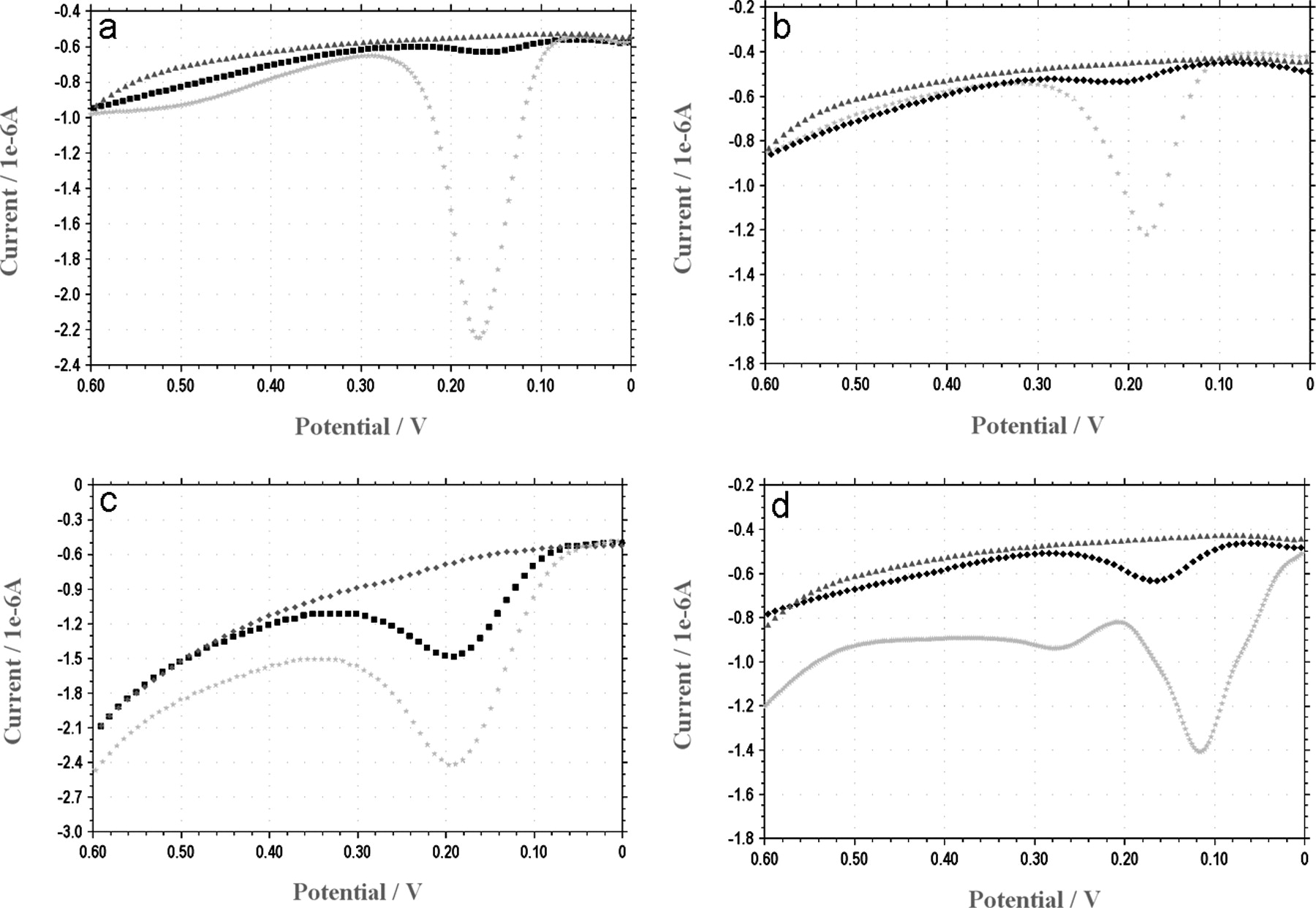
The voltammetry method was used to quantify electrochemical reducing capacities of E. coli coated by catechin, morin, quercetin and red wine (Fig. 9). E. coli were incubated with 1 mM catechin, morin and quercetin (a–c) or with 100 μL of red wine (d). Following 10 minutes of incubation followed by excessive wash in final volume of 20 ml saline, samples were electrochemically analyzed. As can be seen, all four agents used were attached to the bacterial cells and possessed the specific anodic wave of the attached agent (same potential represents the same agent) compared to control bacteria.
Similar patterns of improved antioxidant capacities were also observed when these various bacteria-polyphenols complexes were tested by the DPPH and by the GO chemiluminescence methods (data not shown).
Polycations Enhance Polyphenols Binding to Bacteria.
Previous studies (13) have shown that the cationic polyelectrolytes poly-L-histidine (PHSTD), poly-L-arginine and histone could opsonize bacteria for phagocytosis by both professional and non-professional phagocytic cells. This led us to test whether cationic agents might act as charged ligands also capable of enhancing the binding of polyphenols to bacterial surfaces and to endow upon them even higher TOSC properties. As a model, we have used anaerobic bacteria cultivated from deep gingival crevice pockets, obtained from patients suffering of periodontal disease. After an overnight growth on blood agar, under anaerobic conditions, the bacterial colonies were scraped off, washed in saline and treated for 10 minutes at room temperature by combinations among the polyphenols tannic acid, morin and catechin and the cationic agents poly-L-histidine (PHSTD) (100 μM) and Copaxone® (CPX) (200 μM). The latter is a cationic tetra peptide used in the treatment of multiple sclerosis. Figure 10 shows that, recorded after 6 minutes and expressed as area under the curve (AUC), the anaerobic bacteria had no inhibitory effects on LDCL and PHSTD/CPX inhibited LDCL by 13% and 3%, respectively. However, under the same conditions, the polyphenols morin, catechin and tannic acid reduced the AUC values by 8%, 47% and 52%, respectively, indicating the initiation of an improved TOSC. The figure also shows that while bacteria combined with morin and with PHSTD lowered AUC value by 75%, catechin and tannic acid complexed with CPX induced 69% and 77% of inhibition of AUC, respectively. It is also of note (data not shown) that CPX was also able to enhance TOSC of other micro-organisms complexed with dopamine (also a hydroxyl groups-containing molecule).
Discussion
The present communication shows that several polyphenols, known to be present in red wine as well as in many vegetables and fruit beverages, have the ability to irreversibly bind to surfaces of several Gram positive and Gram negative microorganisms and to significantly enhance their total oxidant-scavenging capacities (TOSC). Furthermore, the highly charged cationic peptides poly-L-histidine and Copaxone®, acting as ligands, could significantly further enhance the binding of polyphenols to microbial surfaces and to enhance their TOSC.
In the current study, we have employed a glucose oxidase (GO) luminol-dependent chemiluminescence method (15, 16) to investigate the TOSC patterns displayed by a series of single polyphenols (Fig. 1). In addition, we used the GO cocktail (Fig. 2), DPPH (Fig. 3) and FRAP (Figs. 4 and 8) assays to measure the ability of polyphenols to enhance TOSC of various bacterial cells. A visual proof for the binding of certain colored flavonoids to VSL#3 bacterial surfaces was also displayed (Fig. 7). The ability of flavonoids to bind to microbial surfaces was also verified by using the Folin-Ciocalteu reagent (Fig. 5) and also by the voltammetry techniques (Figs. 6 and 9).
The techniques used in this work have the ability to detect by different mechanisms total oxidant-scavenging capacities (TOSC). While the FRAP method determines ferric reducing capacities, the DPPH method uses this agent as a single radical, which can be scavenged by antioxidants. On the other hand, the Folin-Ciocalteu’s phenol reagent assay can actually detect and quantify only the number of hydroxyl groups but has no absolute correlation with their antioxidant properties. The GO luminescence cocktail used, which simultaneously generates H2O2, superoxide and cobalt-catalyzed OH· (15, 16) can measure in a dose-dependent manner a large variety of reducing agents and agents (flavonoids) possessing chelating properties for cobalt. The voltammetry methods measure reducing electrochemical potential of various chemical and biological samples. Due to the different mechanisms tested, we cannot expect a complete comparison among the oxidant-scavenging capacities of bacteria coated by the various flavonoids, as measured by different methods. For example, while bacteria coated by rutin did not possess a significant TOSC as measured by the LDCL (Fig. 2) and by the FRAP (Fig. 4) methods, rutin did possess a significant TOSC when measured by the DPPH (Fig. 3) and by the Folin (Fig. 5) assays. Also, gallic acid alone (Fig. 1), which showed a higher inhibitory effect on luminescence compared to tannic acid, had a lesser TOSC when complexed with bacteria (Figs. 2–5 ). This suggests that there is not always a direct correlation between the antioxidant capacities of the various flavonoids alone and their binding affinities to microbial surfaces as expressed by the luminescence method (Figs. 1 and 2). In addition, when bacteria-flavonoid complexes were assayed by the different methods (LDCL, FRAP, DPPH), no absolute correlation was found since, as mentioned above, these measure antioxidant capacities by different mechanisms.
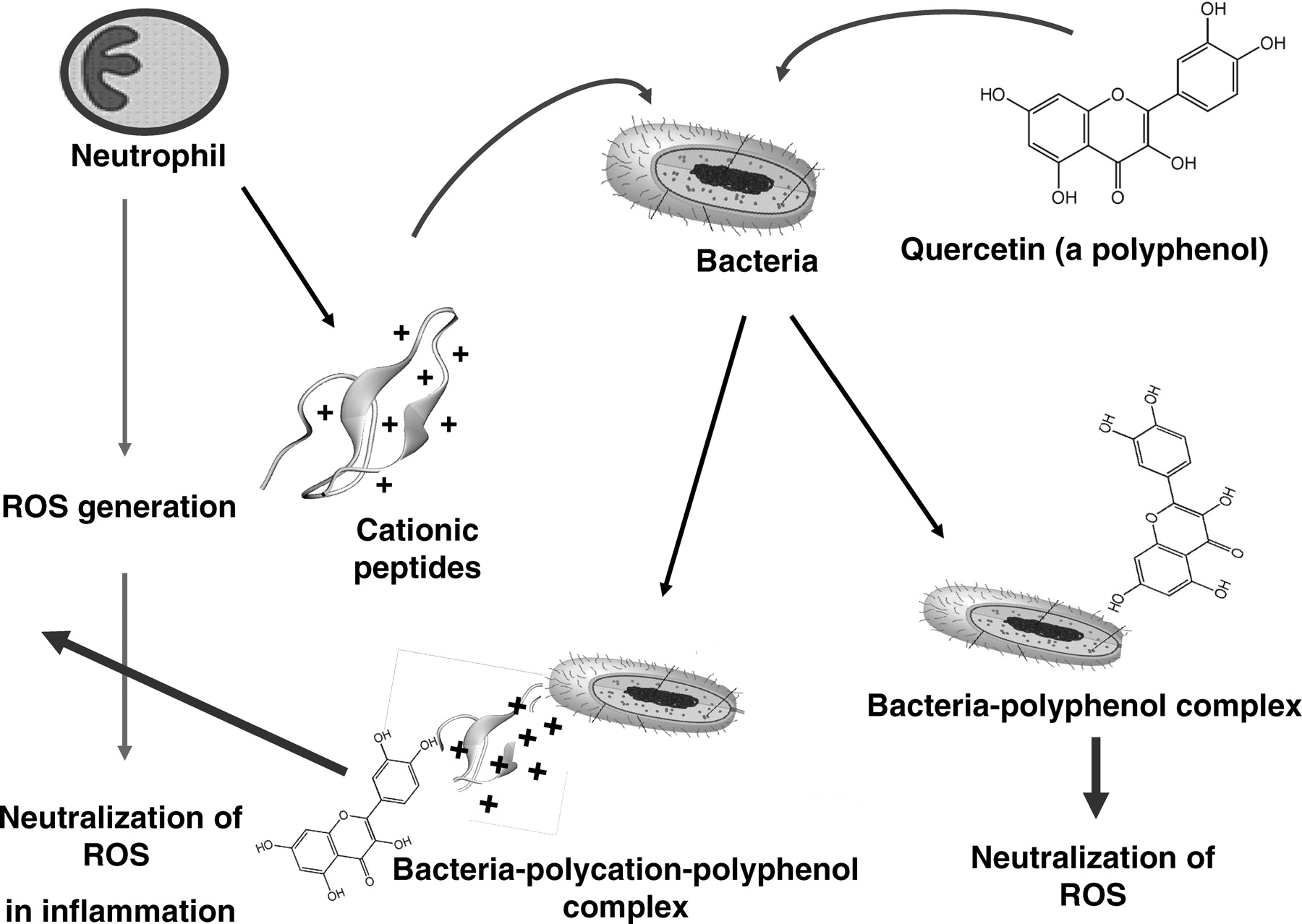
Using all these analytical methods, we confirmed that polyphenols have the ability to bind to and to enhance TOSC of several bacterial species, which could be further enhanced by cationic ligands such as poly-L-histidine and Copaxone® (Fig. 10). Similar results were also obtained by using the cationic ligands nuclear histone, lysozyme and chlorhexidine (not shown). Figure 11 is a schematic representation of a proposed possible mechanisms by which polyphenol-polycation-bacteria complexes might exert their biological and antioxidant activities in inflammation.
Although the nature of the binding of polyphenols to bacterial surfaces is still not fully clear, it is highly suggestive that they do so via hydroxyl groups, hydrophobic interactions, hydrogen bonding, their polymerization degree (22), or also by the presence of a methoxy group in the flavonoid C-ring (23). However, there might be major differences in the binding mechanisms and affinities of polyphenols to Gram positive and Gram negative bacteria. While Gram negatives possess an outer membrane, a possible target for polyphenol binding, the membrane of Gram positive bacteria is shielded from the outside by a thick cell wall (peptidoglycan) as well as by surface polysaccharides which might act both as binding sites for polyphenols (24). In this respect, it was shown that since purified peptidoglycan, but not lipopolysaccharide (endotoxin of Gram negatives), blocked the binding of epigallocatechin gallate (EGCG) from tea to Gram positive bacteria, the rigid cell wall, but not the internal membrane might be the target for polyphenol action. Polyphenols are also known to form complexes with proteins and polysaccharides (25), and it is also probable that the highly polymerized polyphenol tannic acid binds to bacterial surface components rather than to intracellular targets (26).
One example for differences in the binding affinities of polyphenols to bacterial cells is shown in Figure 8. While EGCG strongly enhanced the TOSC of Micrococcus lysodeikticus possessing a “simple peptidoglycan”, it was less effective if bound to Staphylococcus aureus, which possesses a more complex cell wall (27). This might perhaps also explain why while M. lysodeikticus can be readily lysed by lysozyme (a lytic enzyme), S. aureus is more resistant to lysozyme action. This might also indicate that the presence of certain shielding components on S. aureus surfaces might interfere with the binding of polyphenols.
It is also of interest that in addition to its bactericidal activity (26, 28) and its ability to significantly enhance TOSC of bacteria (Figs. 2–6 ), the polyphenol tannic acid, present in large amounts in vegetables, red wine and in tea, was found to protect the Gram-positive Micrococcus lysodeikticus against the direct lytic effect of lysozyme (data not shown). However, lysozyme can also induce bacteriolysis in Staphylococcus aureus due to its ability to function as a cationic peptide and to activate endogenous autolytic wall enzymes resulting in cell lysis (29). The ability of certain polyphenols to protect against bacteriolysis (not shown), might have important clinical implications since they might lower the massive release by bacteriolysis of the highly phlogistic cell wall components lipoteichoic acid, lipopolysaccharide and peptidoglycan, known to trigger inflammatory and post-inflammatory manifestations (30). However, further studies on the biological and structural properties of polyphenols binding sites and their possible role in microbial physiology and pathogenicity are warranted.
A possible relevance of our finings to evaluate the complexity of host-bacteria–flavonoids interactions in vivo is presented in our study. It suggests that following intake of polyphenols antioxidants, these might first interact with the oral epithelium, dentures, periodontal tissue and also with the rich microbial flora present in dental plaques and also in the gingival crevice (colonized mostly by anaerobic bacteria). Later on and downstream, antioxidant polyphenols might also interact with the microbial flora of the large intestines also to exert bactericidal effects (28, 31). Micro-organisms such as Lactobacillus (32–35), Staphylococcus aureus (36), Escherichia coli (37), Candida albicans (38), Pseudomonas aeruginosa (39) and additional microbial species, all possess low-grade antioxidant capacities as protectors against oxidant-dependent phagocytic killing. This suggests a possible additional role for flavonoids where formation of polyphenol complexes with bacteria might be crucial in host-pathogen interrelationships (40). The findings that the VSL#3 bacterial mixtures could also irreversibly bind the highly antioxidative bile-salt bilirubin (Fig. 7) might also be of significance since bile salts co-exist with polyphenols in the gastrointestinal (GI) tract and might also act synergistically to increase TOSC of bacterial cells.
While many polyphenols are highly bactericidal (28), it is paradoxical perhaps that even non-viable bacteria might still be of value. As shown, bacterial surfaces have the ability to bind large amounts of polyphenols and polycations which endows upon them enhanced TOSC against toxic reactive oxygen species (ROS). Also, since microbial flora of the dental plaque in the oral cavity mainly form biofilms, it will be of interest to explore whether polyphenols and polycations could readily penetrate and localize within biofilms to alter their biological activities (41).
What might be the clinical relevance of bacteria-polyphenol-polycation complexes formations in inflammatory processes and in homeostasis in the oral cavity and in the digestive tract? Polyphenols were shown to be effective in the treatment of a large variety of inflammatory conditions in the GI tract (42). Therefore, it might be speculated that inflammatory bowel diseases (IBD), which are characterized by a considerable infiltration of inflammatory cells in the gut mucosa (43), might be ameliorated by polyphenols treatment. In practice, Lactobacilli probiotics are currently used as supplements in the treatment of various GI tract disorders with promising results as they might improve intestinal microbial balance and immune homeostasis (44, 45). The rational selection of probiotic bacteria is based on evidences from animal and human studies (46, 47). Selective deficiencies in intestinal Lactobacillus and Bifidobacterium have been observed in patients with Crohn’s disease (48). Administration of Lactobacillus rhamnosus to children with Crohn’s disease resulted in significant reduction in disease activity index 4 weeks after initiation of therapy (49). Probiotic formulations were also found effective for the prevention (45) or treatment (50) of IBD-related pouchitis, and the therapeutic potential of different probiotics was also evaluated in a rat model of colitis (51).
Of interest are the observations that L-dopa (not shown) and dopamine, both containing hydroxyl groups and possessing antioxidant properties (Fig. 1) could also bind to bacterial cells (Fig. 2). Copaxone®, a cationic peptide, widely used in the treatment of certain types of multiple sclerosis patients (52), acts similarly to other polycations, capable of enhancing the binding of polyphenols to bacterial cells (Fig. 10). Therefore, the cationic agent Copaxone® may perhaps also serve as a ligand, capable of binding L-dopa and flavonoids to neural cells to increase their protection against free radicals. Furthermore, polycations, especially those released from activated neutrophiles, might have additional regulatory functions in inflammatory sites by acting as opsonizing agents, activators of the respiratory burst in neutrophils, and as binders of immune complexes to target cells (13). Polycations can also cationize superoxide dismutase and catalase, making them more protective against oxidative stress (53).
Since we found that polyphenols enhanced TOSC of a variety of microbial species, including lactic acid bacteria species (Figs. 2–7 ), Staphylococcus aureus and Micrococcus lysodeikticus (Fig. 8) and Escherichia coli (Fig. 9), it is suggested that stable bacteria-polyphenols-polycations complexes (Figs. 10, 11) might be effective also in clinical uses. To circumvent the metabolism and degradation of polyphenols to less active metabolites if administered per os, it might be speculated that supplementation of probiotic Lactobacilli, pre-coated by mixtures of polyphenol antioxidants and cationic agents might act as “slow release devices” to ameliorate GI disorders if administered either per os or via enemas directly into the caecum. Since additional microbial species have also been found to strongly bind antioxidant polyphenols and to significantly increase their TOSC, it appears that this is a universal phenomenon that can have advantages also for use in other therapeutic regimens.
Taken together, although polyphenols and cationic peptides can readily kill bacteria (28, 29), it is paradoxical perhaps that there might still be some beneficial values to the “life after death” phenomenon, whereby non-viable bacteria might still function as effective carriers of antioxidant polyphenols. Such complexes (Fig. 11) might serve as protective agents against tissue damage in a variety of pathophysiological systems as well as against mutagenesis.
Modulation of luminol dependent chemiluminescence induced by the glucose oxidase (GO) cocktail by polyphenols (10 μM) and by red wine and cranberry beverages (10 μL). Note that line f represents the same activities induced by 5 agents, line g represents the activities of quercetin and gallic acid, and line h represents the activities of wine and cranberries. Data are presented as average of 4 replications. Modulation of luminol dependent chemiluminescence induced by the glucose oxidase (GO) cocktail by a mixture of 8 probiotic species (VSL#3) pre-coated by polyphenols (1 mM) and by red wine (100 μL). Data are presented as average of 4 replications. DPPH patterns expressed as gallic acid equivalents (GAE) (μM) of a mixture of 8 microbial probiotic species (VSL#3) pre-coated by polyphenols (1 mM) by red wine and by a cranberry beverage at 100 μL (see Materials and Methods). Note that the most potent scavengers of the DPPH radical were tannic acid and red wine. Data are presented as mean values ± SD (P < 0.05, n = 6). FRAP patterns expressed as gallic acid equivalents GAE (μM) of a mixture of 8 microbial probiotic species (VSL#3) pre-coated by polyphenols (1 mM), by red wine and by a cranberry beverage at 100 μL (see Materials and Methods). Note that as shown in Figure 3, the most potent inhibitors of iron reduction were wine and tannic acid. Data are presented as mean values ± SD (P < 0.05, n = 6). Folin-Ciocalteu’s phenol assay patterns, expressed as gallic acid equivalents (GAE) of a mixture of 8 microbial probiotic species (VSL#3) pre-coated by polyphenols (1 mM), by red wine and by a cranberry beverage at 100 μL (see Materials and Methods). Data are presented as mean values ± SD (P < 0.05, n = 6). A cyclic voltammogram of a mixture of 8 microbial probiotic species (VSL#3) complexed with increasing concentrations of tannic acid (0.1–1 mM). Note that while VSL#3 alone (a) did not show any anodic waves, when used at 100 μM, tannic acid alone (f) showed three anodic waves (at 0.22, 0.35 and 0.63 Volts). When the VSL#3 preparation was combined with increasing concentrations of tannic acid (0.1, 0.25, 0.5 and 1.0 mM; b–e, respectively) and washed to remove unbound material, a dose-dependent increase in the current was obtained. The cluster of 6 lines on top represents cathodic waves which indicate that the reaction is not electrochemically reversible. These findings support the assumption that the bacterial cells firmly bound tannic acid (a representative experiment). VSL#3 preparation stained by various polyphenols, beverages and by bile salts. A thick suspension of VSL#3 was incubated for 10 minutes with (a) saline, (b) polyphenon-60, (c) tea extract, (d) red wine, (e) catechin, (f) morin, (g) bilirubin and (h) biliverdin and then washed with saline to remove unbound materials. The concentrations used for this experiment were 500 μM for purified materials and 100 μL from beverages’ extracts (see Materials and Methods). Note the ability of the various colored agents to irreversibly bind to the bacterial cells. The upper series represents the visual appearance of the colored bacteria and the lower series represents the pellets obtained following 5 minutes of centrifugation (at 1000 g) of the reaction mixtures. A color version of this figure is available online. Enhanced oxidant-scavenging capacities of Staphylococcus aureus and Micrococcus lysodeikticus (ML) following incubation with various polyphenols, as measured by the FRAP assay and expressed as gallic acid equivalents GAE (μM). The figure shows that wine at 100 μL and tannic acid and curcumin at 1 mM endowed the bacteria significant oxidant-scavenging capacities. The figure also shows that EGCG was effective only when bound to Micrococcus lysodeikticus. Data are presented as mean values ± SD (P < 0.05, n = 6). Four voltammograms of Escherichia coli complexed with catechin (a), morin (b), quercetin (c) and red wine (d). Figures 9a, 9b and 9d were analyzed using Differential Pulse Voltammetry analyzing technique (DPV). Figure 9c was analyzed using Cyclic Voltammetry (CV) technique (see Materials and Methods). Note that while E. coli alone did not show any anodic waves (upper curves-solid triangle, ▴), all four agents tested alone showed wide anodic waves (lower curve-solid star, ★) and bacteria coated by the four agents showed anodic waves representing the agent attached to the bacteria (middle curves-solid square, ▪). These findings support the assumption that the bacterial cells firmly bound these agents (a representative experiment). Luminol-dependent chemiluminescence of bacteria-polyphenol-cationic peptide complexes. Suspensions of a mixed anaerobic flora (BACT), isolated from the oral cavity (see Materials and Methods) were pre-coated by various polyphenols in the presence or absence of the cationic agents poly-L-histidine (PHSTD) or Copaxone® (CPX). Following several washes to remove unbound material, samples were tested for their capacity to quench light induced by the GO cocktail. The figure shows that expressed as area under the curve (AUC) and measured after 6 minutes, the most potent enhancement of antioxidant activity was obtained by bacteria pre-treated by combinations of polyphenols and cationic peptides. Note: the numbers indicated on bars represent % inhibition of AUC. Data are presented as mean values ± SD (P < 0.05, n = 6). A schematic representation of the possible interactions among bacteria, polyphenols and cationic peptides, which are likely to occur in vivo in inflammatory sites. Note that TOSC can be enhanced by polyphenols interacting either directly with the bacterial surface or via a cationic ligand likely to be released by activated neutrophils in inflammatory sites. This might contribute to the attenuation of oxidative stress in infectious and inflammatory sites.



Footnotes
This work was supported by an endowment fund by the late Dr. S. M. Robbins of Cleveland, OH, USA, by the Israel Science Foundation (grant no. 241/04) and by the Ydidut Foundation (Mexico).
Acknowledgements
R. K. is affiliated with the David R. Bloom Center for Pharmacy and the Brettler Center for Research in Molecular Pharmacology and Therapeutics, School of Pharmacy, The Hebrew University of Jerusalem.