
Abstract
To determine the possible roles of tumor necrosis factor-α (TNFα) and nitric oxide (NO) in the bovine oviduct, ampulla and isthmus collected during the estrous cycle were exposed for 18 h to TNFα, NO donor (NONOate), NO synthase inhibitors (L-NOARG, L-NAME and AMT) and oxytocin (OT) as a positive control. Prostaglandins (PGs) and NO2/NO3 in conditioned media were measured. TNFα stimulated PGF2α secretion on Day 0 (onset of estrus = Day 0) and Days 2–3, in both the ampulla and isthmus, but on Days 18–20 only in ampulla. TNFα increased PGE2 secretion in both fragments in each phase. NONOate did not affect PGF2α secretion on Days 18–20, whereas this NO donor stimulated PGF2α secretion in both fragments on Day 0 and Days 2–3. TNFα increased NO2/NO3 production in every examined phase in the ampulla and on Days 2–3 in the isthmus. L-NAME lowered NO2/NO3 production regardless of phase or fragment. L-NOARG and AMT lowered NO2/NO3 production in both fragments on Day 0 and Days 2–3. The possible role of TNFα, NO or PGs on the oviductal contractility during the early-luteal phase was also examined. Neither TNFα nor NONOate influenced contractility in either fragment. Although PGF2α stimulated the contraction in both fragments, PGE2 decreased it. When taken together, TNFα seems to play some role as a modulator of PGF2α and PGE2 production and for transferring the embryo from the oviduct to the uterus by stimulating NO production in the bovine oviduct.
Introduction
The oviduct plays a key role in the accommodation of gametes, fertilization and early embryo development and its passage to the uterus. It has been shown that products of each oviductal part (i.e. isthmus and ampulla), oviductal epithelial cells, as well as oviductal fluid are necessary to the normal and efficient development of the embryo (1, 2). The oviduct produces some characteristic proteins, but also several steroidal and protein hormones, and eicosanoides—derivatives of the essential fatty acids, which may regulate early embryonic development and play a key role in embryo transport (3, 4).
Tumor necrosis factor-α (TNFα) plays an important role as a regulator in the female reproductive tract (5–7). It was demonstrated that TNFα affects PGs synthesis in the bovine endometrium (8–11) and luteal cells (12). The key role of PGs in animal reproduction is well documented (13). In humans and mice, TNFα is known to be produced by macrophages and epithelial oviductal cells, even by the blastocyst (6, 7). TNFα has also been demonstrated to be produced by bovine oviductal epithelial cells (14). Nitric oxide (NO) acts as a major paracrine mediator during many processes engaged in female reproduction (15–20) and was also demonstrated in bovine oviduct (21). NO is produced by the conversion of L-arginine to L-citrulline by the enzyme NO synthase (NOS) in various tissues and cells. At present, three isoforms of NOS have been described, i.e. neuronal NOS (nNOS) in the nervous system, endothelial NOS (eNOS) as a constitutive NOS mainly in endothelium and inducible NOS (iNOS). These are all produced by activated macrophages, hepatocytes and neutrophiles (22). Transcripts of the eNOS and iNOS were detected in bovine oviductal epithelial cells (BOEC) during the estrous cycle (21). NO activity was low during the late proestrus cycle in the rat oviduct (23) and eNOS in the rat oviduct was most distinctive during the proestrus and estrus (24). NOS was demonstrated in the oviductal epithelium of rat (23, 24) and pig (25).
The oviductal contractility seems to be dependent on the estrous stage. The relaxing effect of NO on smooth muscle and hence, uterine quiescence during pregnancy is well documented (26). On the other hand, oviductal contractility arises around the estrus (27). Several factors may contribute in the regulation, influence or control of the NOS expression and NO production in the oviduct, e.g. estradiol, oxytocin, PGs, endothelin-1 (ET-1) and cytokines (28, 29). Prostaglandin F2α (PGF2α) induces NO synthesis in rat oviduct (30). NO may negatively influence or antagonize the contractile response of PGF2α and, even more, NO releases prostaglandin E2 (PGE2) (31). It has been previously demonstrated that PGE2 increases at estrus and leads to oviduct relaxation in the presence of progesterone in the cow (32). In the present study, we examined the role of NO as a possible mediator of TNFα action in the bovine oviduct. Moreover, the effect of TNFα and its mediators, PGF2α, PGE2 and NO, on oviductal contractility in the early-luteal phase was also studied.
Materials and Methods
Animals and Collection of Oviduct Tissues.
Healthy, normal-cycling Polish Holstein Black and White cows were used for the collection of the oviducts. The animals were culled by the owners (from a farm in Rogale and the “Farmer” in Zalesie near Szczytno, Poland) from the dairy herds because of lower milk production (2005–2008). The estrus of the cows was synchronized using an analogue of PGF2α (dinoprost, Dinolytic; Pharmacia & Upjohn N.V./S.A., Belgium) injections with an 11-day interval, as recently described and recommended (33). The onset of the estrus was determined by a veterinarian via per rectum and ultrasonography examination using DRAMINSKI ANIMALprofi Scanner (Draminski Electronics in Agriculture, Olsztyn, Poland; www.draminski.com) and confirmed by observing the signs of estrus (i.e. vaginal mucus, standing behavior). The onset of estrus was taken at Day 0 of the estrous cycle. Bovine oviducts were obtained at a local slaughterhouse within 20 min of exsanguinations and were transported on ice to the laboratory within 40 min.
All procedures were approved by the Local Animal Care and Use Committee in Olsztyn, Poland (Agreements No. 73/2002/N and 06/2007/N).
Isolation and Culture of Oviduct Explants.
The oviducts from the side of the active ovary (collateral to the ovary with dominant follicle or developing corpus luteum) were collected at three different stages of the cycle: estrus (Day 0; n = 8), early-luteal phase (Days 2–3; n =14) and the follicular phase (Days 18–20; n = 8). The oviducts were dissected from the mesosalpinx, utero-ovarian ligament and other surrounding tissues. The ampulla and isthmus were incised longitudinally and then were cut on 20–30 mg explants. Each of them was placed into a glass vial containing 2 ml of incubating medium DMEM/Ham’s F-12 medium (Sigma Chemical Co., St. Louis, MO, USA, # D-8900) supplemented with 0.1% of bovine serum albumin (BSA) (Sigma, # A9056), 5 ng/ml sodium selenite (Sigma, # S1382), 0.5 mM ascorbic acid (Sigma, # A1417) and 20 μg/ml gentamicin (Sigma, # G-1397). The tissue was cultured in a water bath with mechanical shaker at 38.5°C and controlled atmosphere (95% air with 5% CO2) as described previously (34).
Experiment 1: The Effect of TNFα and NO on PGs and Stabile NO Metabolites Secretion by the Bovine Oviduct.
To determine the possible effect of TNFα and NO on prostaglandin and stabile NO metabolites output from the bovine oviduct, the ampulla and isthmus were collected during estrus (n =8), early-luteal phase (n =8) and follicular phase (n = 8). The oviductal fragment explants were exposed to human recombinant TNFα, (Dainippon Pharmaceutical Co., Ltd., Osaka, Japan, # HF-13; 10 ng/ml); a nitric oxide donor, Spermine Nitric Oxide Complex (NONOate) (Sigma, # S-150; 10−4 M); a competitive inhibitor of endothelial isoform of NO synthase, Nω-nitro-L-arginine (L-NOARG) (Sigma; 10−4 M); a competitive inhibitor of inducible isoform of NO synthase, (±)-2-amino-5,6-dihy-dro-6-methyl-4H-1,3-thiazine (AMT) (Sigma, # A253; 10−4 M); and a nonspecific inhibitor of NO synthase, Nω-nitro-L-arginine methyl ester (L-NAME) (Sigma, # N5751; 10−4 M). Oxytocin (OT) (Sigma, # O4375; 10−7 M) was used as a positive control. Concentrations of all reagents were chosen based on the previous studies (11, 35–37) and confirmed by the preliminary experiments (data not shown). After 18 h of the incubation, the conditioned media were collected in tubes with 10 μl 0.3 M EDTA, 1% aspirin solution (pH 7.3) (Sigma, #A2093), and frozen until measurement of PGF2α and PGE2 using EIA method. Nitric oxide production was quantified by measuring stabile NO metabolites (nitrite/nitrate, NO2 /NO3) concentrations in the medium (36).
Experiment 2: Effect of a Non-Specific NOS Inhibitor (L-NAME) on TNFα-Stimulated PG and Stabile NO Metabolites Secretion by the Bovine Oviduct.
To determine whether NO may mediate TNFα-stimulated PGs synthesis by the bovine oviduct, the explants from the early-luteal phase (Days 2–3 of the cycle; n = 6) were exposed to TNFα (10 ng/ml), L-NAME (10−4 M) or both reagents together (L-NAME and TNFα; 10−4 M and 10 ng/ml, respectively). After 18 h culture, the conditioned media were collected and frozen until measurement of PGF2α, PGE2 and NO2/NO3.
Experiment 3: The Effect of TNFα, NO and PGs on Contractility of the Bovine Oviduct.
The possible effects of TNFα, NO and PGs on the contractility of the bovine oviduct during the early-luteal phase (Days 2–3; n =6) were determined in vitro as described previously (37–39). The 5-mm-long rings from ampulla and isthmus were vertically fixed in separate 10 ml tissue chambers in Krebs’ solution (KRB) (NaCl, 120.3 mM; KCl, 5.9 mM; CaCl2, 2.5 mM; MgCl2, 1.2 mM; NaH2PO4, 1.2 mM; NaHCO3, 15.5 mM; glucose, 11.5 mM; pH 7.3–7.4) at 38.5°C and aerated with 95% O2:5% CO2. The smooth muscle contractility was measured using Hugo Sachs Electronik Apparatus (HSE, D-79233, March-Freiburg, F.R. Germany) equipped in isometrical transducer type 372 (HSE Isometric Force Transducer F30). One edge of the ring was connected with cotton ligature and attached to an isometric contraction transducer (HSE, Type 372, Germany). The other edge was secured with steel hook to the bottom of the tissue chamber. The primary data registered by the transducer were analyzed using HSE-ACAD computer software (HSE).
During 2 h preincubation the basic voltage 10 ± 1.0 was assessed and the spontaneous, regular contractions of the oviduct were observed (stabilization period). After stabilization period the oviductal rings were stimulated with saline (control), TNFα (10 ng/ml), NONOate (10−4 M), PGE2 (10−6 M) and PGF2α (10−6 M). Noradrenalin (NA) (Levonor, Polfa Warszawa, Poland; 10−6) and acetylcholine (Ach) (Sigma, # A6625; 10−6) were used as negative and positive controls, respectively. The concentrations of main experimental reagents (TNFα and NONOate) were chosen in Experiment 1. The concentrations of both prostaglandins and reference substances (NA and Ach) were chosen based on the previous studies (26, 37–39).
The control registration (at least 30 min) was performed before and/or after application of every factor. This period was sufficient to recover a spontaneous pattern of rhythmical smooth muscle contractions. After the investigated substance application, a 10-min period was designated for observation of the tissue reactions. Next, the chamber was rinsed 3 times using KRP solution. After 30 min the stimulation procedure was repeated.
Analytical Methods.
The concentration of PGF2α and PGE2 in the culture medium was determined with the enzyme immunoassay as described previously (11, 34). The PGE2 standard curve ranged from 0.078 to 20 ng/ml and the ED50 of the assay was 1.25 ng/ml. The intra- and interassay coefficients of variation were 6.9% and 9.7%, respectively. The PGF2α standard curve ranged from 0.19 to 50 ng/ml and the ED50 of the assay was 250 ng/ml. The intra- and interassay coefficients of variation were on average 7.9% and 10.4%, respectively.
The concentrations of nitrite/nitrate (NO2/NO3) in the medium were measured by a colorimetric method using the Griess reaction as described by Green et al. (40) with our own modification (36). Nitrate standard (Sigma, # S2252) was diluted in the deionized water to the final dilution 100 μM (6.9 mg/ml). Azotine reductase (0.2 U/2 ml; Sigma, # N7265) was diluted in previously performed buffer with pH 7.2, containing: 10 mM glycine (Sigma, # G4410); 50 mM 3-(N-morpholino)propanosulphate acid (MOPS) (Sigma, # M5162) with pH 7.2; 20% of glycerol (Sigma, # G6279); and 1% of bovine serum albumin (BSA) (Sigma, # 2153). NADPH (Sigma, # N7505) was diluted in deionized water to a final dilution 2 mM (1666.8 mg/l). The assay sensitivity was 0.083 μg/ml and the standard curve ranged from 0.05 μg/ml to 6.9 μg/ml. The intra- and interassay coefficients of variation were on average 8.6% and 14.7%, respectively.
Statistical Analysis.
The data of prostaglandins and nitrite/nitrate concentrations (Experiments 1 and 2) are shown as the mean ± SEM of values obtained in 6 to 8 separate experiments, each performed in triplicate. Tissue weight was used to standardize the results. The statistical differences between controls and treated groups were assessed by a one-way analysis of variance (ANOVA) followed by Bonferroni’s Multiple Comparison Test (GraphPad PRISM version 4; GraphPad Software Inc., San Diego, CA, USA).
The spontaneous and induced contractility of oviductal segments (amplitude, frequency of the picks; Experiment 3) were calculated using GraphPad PRISM curves and regression analysis software. The effects of the applied reagents were calculated as area under the curve. The baseline was defined as: Y =0.0. The effects (area under the curve) of added substances on the contraction during the 10-min experimental period were compared with control values obtained during the same time point and the differences were assessed by a one-way ANOVA, followed by the Bonferroni’s multiple comparison test (GraphPad PRISM).
Results
Effects of TNFα and NO on PGs and Stabile NO Metabolites Secretion by the Bovine Oviduct.
Figure 1 shows PGF2α secretion by oviductal explants in response to OT, TNFα, NONOate, L-NOARG, AMT and L-NAME.
OT (positive control) strongly stimulated PGF2α secretion in the ampulla on Days 18–20, 0, 2–3 of the estrous cycle (P < 0.001), and in isthmus at each phase of the estrous cycle (P < 0.05). TNFα stimulated the secretion of PGF2α on Day 0 and Days 2–3 in the ampulla (P < 0.001), as well as in the isthmus (P < 0.01). However, on Days 18–20, TNFα stimulated the PGF2α secretion only in the ampulla (P < 0.001). NONOate did not stimulate PGF2α secretion on Days 18–20. However, NONOate strongly stimulated the PGF2α secretion in the ampulla on Day 0 and Days 2–3 (P < 0.001) and in the isthmus (P < 0.001). An inhibitor of endothelial isoform of NOS (L-NOARG) as well as inducible isoform (AMT) inhibited PGF2α secretion, both in the ampulla and isthmus, on Day 0 and Days 2–3 of the estrous cycle (P < 0.05). The influence of both specific NOS inhibitors on PGF2α secretion at the follicular phase was not observed. The non-specific NOS inhibitor L-NAME inhibited PGF2α secretion at every examined phase and by oviduct explants (P < 0.05).
Figure 2 shows PGE2 secretion by oviductal explants in response to OT, TNFα, NONOate, L-NOARG, AMT and L-NAME. TNFα strongly increased PGE2 secretion in each examined phase, in both oviductal fragments, compared with control (P < 0.001). NONOate stimulated PGE2 secretion only in the isthmus at each examined phase (P < 0.001). On Days 2–3 of the estrous cycle both specific NOS inhibitors (L-NOARG and AMT) lowered PGE2 secretion in the ampulla (P < 0.05) and in the isthmus (P < 0.01). A competitive inhibitor of endothelial isoform of NO synthase, Nω-nitro-L-arginine (L-NOARG), and AMT on Days 18–20 did not influence PGE2 secretion (P > 0.05). A nonspecific inhibitor of NO synthase, Nω-nitro-L-arginine methyl ester (L-NAME), lowered the PGE2 secretion in the ampulla (P < 0.05) and in the isthmus (P < 0.05) in every examined period.
Figure 3 shows NO secretion by the oviductal explants after OT, TNFα, NONOate, L-NOARG, AMT and L-NAME stimulation. TNFα preferentially stimulated NO production in ampulla on Days 2–3 (P < 0.001) and in the isthmus (P < 0.01). TNFα stimulated NO production only in the ampulla on Day 0 (P < 0.01) and on Days 18–20 (P < 0.05). Neither of the specific inhibitors of NOS isoforms (L-NOARG nor AMT) influenced NO production on Days 18–20. Both L-NOARG and AMT inhibited NO production on Day 0 of the estrous cycle in the ampulla and isthmus (P< 0.05). Both L-NOARG and AMT decreased NO2/NO3 production on Days 2–3 in the isthmus (L-NOARG, P < 0.05; and AMT, P < 0.001), as well in the ampulla (P < 0.001). The non-specific NOS inhibitor (L-NAME) decreased NO2/NO3 production in ampulla as well as in isthmus, on Days 2–3 (P < 0.001) as well as on Day 0 and Days 18–20 (P < 0.01).
Effect of NOS Inhibitor (L-NAME) on TNFα-Stimulated PG and Stabile NO Metabolites Secretion by the Bovine Oviduct.
Figure 4 shows PGF2α and PGE2 secretion in response to TNFα, L-NAME or TNFα together with L-NAME.
TNFα stimulated PGF2α secretion both in the ampulla and isthmus (P < 0.001). TNFα strongly stimulated PGE2 secretion both in the ampulla and isthmus (P < 0.001). The non-specific NOS inhibitor (L-NAME) inhibited basal PGF2α and PGE2 secretion both in the ampulla and isthmus (P < 0.05). TNFα stimulated PGF2α and PGE2 secretion was reduced by L-NAME both in the ampulla and in the isthmus (P < 0.01). TNFα stimulated NO2/NO3 secretion was reduced by L-NAME both in the ampulla and isthmus (P < 0.01).
Effect of TNFα, NO and Prostaglandins on Motor Activity of the Bovine Oviduct.
Figures 5 and 6 show the effect of TNFα, NONOate, PGF2α and PGE2 on the contractility of bovine oviduct explants on Days 2–3. The effects of investigated factors on oviduct contractility are presented as area under the curve (Table 1). Acetylcholine (positive control) increased contractile activity in the ampulla (P < 0.01) and isthmus (P < 0.05), while NA (negative control) decreased such activity (P < 0.001). Neither TNFα nor NONOate influenced contractility in either the ampulla or the isthmus (P > 0.05; Fig. 5; Table 1) on Days 2–3. PGF2α stimulated the contractility of the ampulla (P < 0.001), as well as the isthmus (P < 0.01). PGE2 decreased in contractile activity in both oviductal fragments (P < 0.05).
Discussion
It has been previously shown that TNFα and its receptor are highly expressed in the bovine oviduct (14, 41). In the present study, it was shown that TNFα strongly augments PG production in the bovine oviduct in vitro. The stimulating effect of TNFα on either PGF2α or PGE2 production was partially reduced by non-specific (L-NAME) and specific NOS inhibitors (L-NOARG, AMT). The current data partially confirmed the earlier results of Wijayagunawardane et al. (14), which demonstrated, using the microdialysis system, that TNFα influenced secretion of contraction-related substances (PG, angiotensin II and ET-1) and might modulate local contraction in the bovine oviduct. The highest TNFα impact on prostaglandin synthesis was observed in the early phase of the estrous cycle (post-ovulatory period). However, there is strong evidence that TNF-induced PG production in the bovine oviduct not only depends on the stages of the estrous cycle, but also differs in the oviductal fragments. In the present study, the oviduct was divided into two anatomically different fragments, the ampulla and isthmus, to observe whether they also differed physiologically. It was clearly demonstrated that these two fragments have different responsiveness to TNFα and NO. After ovulation (Days 2–3 of estrous cycle), TNFα and NO increased PGF2α secretion mainly in the ampulla, whereas TNFα and NO increased PGE2 secretion in the isthmus. Moreover, it was found that the basic PGF2α production was higher in the ampulla than in the isthmus, but the basic PGE2 production was higher in the isthmus than in the ampulla. The data confirm the importance of TNFα in the PG production by the bovine oviduct during the periovulatory period, which might optimize the early development of the embryo and possibly regulate contraction to normalize oviductal embryo passage.
It was observed that NO, when stimulated by TNFα, seems to play a role in the secretory function of the oviduct in the early-luteal phase. The stabile metabolites of NO stimulated by TNFα increased in both oviductal fragments mostly on Days 2–3. It was shown that TNFα and NO stimulated PGF2α production and output mainly in the ampulla, and further, it was clearly demonstrated that PGF2α stimulated contractility in both the ampulla and isthmus. NO- and TNFα-dependent PGE2 output was observed mainly in the isthmus. Consequently, PGE2 was found to be a potent inhibitor of oviduct motility. Prostaglandins are potent regulators of the oviductal contractility (42–44). It is well known that the concentrations of PGF2α and PGE2 in the porcine oviduct change during the estrous cycle (44). The highest concentrations of PGs are observed in the follicular phase and the periovulatory period (44). In addition, local production of PGE2, PGF2α and other contraction-related substances (PG, angiotensin II and ET-1) in the cow, as well as their involvement in the cyclic regulation of functions, have been shown in the oviduct (4, 14, 32, 37, 41, 45). PGF2α is involved in contractile activity, whereas PGE2 takes part in relaxant activity of the porcine oviduct (14, 44, 46). Interestingly, PGF2α regulates the contractility stronger in the ampulla, although the muscle layer in the ampulla is not as organized as in the isthmus. The ampulla plays a role in oocyte capture and transports the oocyte, through the ciliated epithelial cell, to the site of fertilization. Thus, it is very possible that the ciliated epithelial cells movement is supported by contractile factors and such mechanisms could occur at ovulation and just before conception. The paracrine factors, through secretion of PGF2α in the ampulla on Days 2–3 of the estrous cycle or pregnancy, may increase the contractility of the isthmus. Thus, the suitable and optimal conditions to transport the embryo from oviduct to the uterus may be provided.
It has been demonstrated that PGF2α may trigger the increase in contractility in both oviductal fragments and PGE2 decreases the muscle tonus. TNFα- and NO-stimulated PGF2α secretion takes place mainly in the ampulla, but TNFα-stimulated PGE2 production takes place mainly in the isthmus. After fertilization, the embryo is located in the ampulla for the 2–3 days. The ampulla has a poorer muscular layer than the isthmus (1) and shortly after ovulation and conception it is necessary to support, by oviduct, the special conditions for the early embryo development which take place in the ampulla. During the retention of ova in the oviducts, the active preovulatory contractions disappear and ovum transport is arrested at the ampullar-isthmic junction (1, 14, 45). It has been suggested that the patterns of oviduct contraction during the periovulatory period are temporally and strictly controlled by local mechanisms that are responsive to the needs of the zygote (14, 45). One could speculate that the ampulla is the main site of PGF2α production and, on the other hand, PGE2 output is strongly regulated in the isthmus at the early-luteal phase. The differences in PGF2α and PGE2 secretion and action in the ampulla and isthmus may indicate their functional divergence in gametes maturation, fertilization and early embryo development in different species.
In the present study the direct impact of NO donor or TNFα on bovine oviductal contractility was not observed under in vitro condition. However, in the female reproductive system, NO is responsible for myorelaxation of the pregnant uterus and, in the same way, prevents premature uterus contractility triggering parturition (15, 16, 26). Pregnancy, as well as the follicular phase of the estrous cycle, is characterized by an increase in eNOS synthesis in the endothelium and elevation in uterine blood flow (17). NO has also been shown to take part in the release of PGE2, mostly at the estrus (32), and to cause relaxation of the oviduct in the presence of progesterone (25). On the other hand, NO facilitates the parturition, triggers myorelaxation cervix of the uterus and increases blood flow in uterine arteries (17). Moreover, NO inhibitor caused muscle contractility of the pig blood vessel in the reproductive tract (39). Thus, we have supposed that NO, in a similar way as in the uterus, may act on the oviductal motor activity. Although neither TNFα nor NO showed direct influence on the basal oviductal contractility in the present study, it might be suggested that they exert its myorelaxative influence in concert with other autocrine/paracrine factors including PG (28–30). The results from in vitro experiments may not adequately represent in vivo condition. The regulation of oviductal motor function in vivo is a composed process and depends on co-operation and interactions with several auto-paracrine factors. It has been previously shown that paracrine factors (PG, ET-1, angiotensin II, OT) may be necessary for both oviductal contractility and early embryo development (45). Rosselli et al. (29) demonstrated that L-NAME (a NO inhibitor) increased oviductal contractility evoked by ET-1. NO could also negatively influence, or even antagonize, the contractile response of PGF2α (21). An inhibitor of NOS (L-NMMA) enhanced PGF2α induced contraction of the oviduct, providing indirect evidence that there is a basal release of NO in the oviduct, which may reduce and/or modulate the contractile effects of PGF2α (30). It means that ET-1 and PGF2α induced contraction of the oviduct is reduced/modulated by endogenous basal release/synthesis of NO (28, 30). It may be supposed that NO does not cause the muscle relaxation directly, but acts as a modulator and/or modulator of the muscle layer tonus, and may antagonize the contractile action of different factors. Moreover, the stimulation of the tubal muscle layer by the NO inhibitor caused myorelaxation of the human oviduct (47). NO inhibitors have been in vivo demonstrated to increase tubal motility that results in accelerated ovum transport and passage of oocytes and embryos to the uterus, probably as a result of intensification of oviductal contractility (48). Ekerhovd et al. (47) showed that the NO precursor provoked an initial, but short-lived, increase in basic tonus in the human oviduct. All these data indicate that NO could act as a paracrine signal between different layers of the oviductal wall, providing a role for endogenous NO in regulation of tubal function.
Taken together, TNFα and NO stimulate the PG secretion dependently on oviductal fragment and phase of the estrous cycle. The most evident effects of TNFα and NO on PGF2α synthesis was observed in the ampulla on Day 0 and Days 2–3. The effects of TNFα on PG secretion by the bovine oviduct are only partially mediated by NO. Although in our in vitro study TNFα and NO do not influence basal oviductal contractility directly, the contribution of both factors on myorelaxation and/or myocontraction of the bovine oviduct might be mediated by other autocrine/paracrine factors including PG.
Effects of Saline (Control), Tumor Necrosis Factor-α (TNFα; 10 ng/ml), a Nitric Oxide Donor (NONOate; 10−4M), Prostaglandin F2α (PGF2α; 10−6 M), Prostaglandin E2 (PGE2; 10−6 M), Acetylocholine (Ach; 10−6 M) and Noradrenaline (NA; 10−6 M) on Contractility of the Ampulla and Isthmus of Bovine Oviduct (Day 2–3 of the Cycle) a
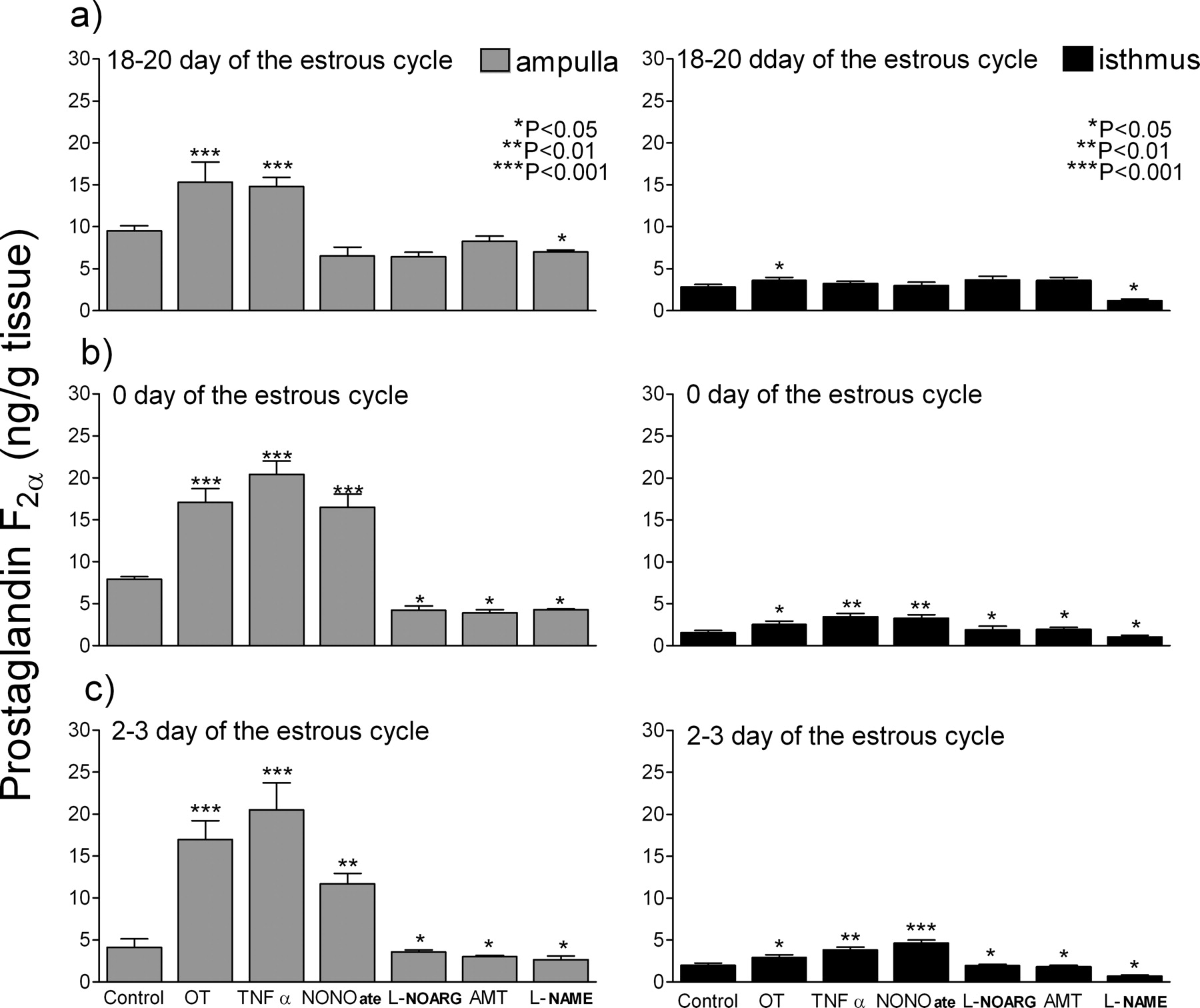
Effects of tumor necrosis factor-α (TNFα; 10 ng/ml); a nitric oxide donor, Spermine Nitric Oxide Complex (NONOate; 10−4 M); a competitive inhibitor of endothelial isoform of NO synthase, Nω-nitro-L-arginine (L-NOARG; 10−4 M); a competitive inhibitor of inducible isoform of NO synthase, (±)-2-amino-5,6-dihydro-6-methyl-4H-1,3-thiazine (AMT; 10−4 M); a nonspecific inhibitor of NO synthase, Nω-nitro-L-arginine methyl ester (L-NAME; 10−4 M); and Oxytocin (OT; 10−7 M) on prostaglandin F2α production in the ampulla and the isthmus at 18–20 Days, 0 Day and 2–3 Days of the oestrus cycle in the bovine oviduct.
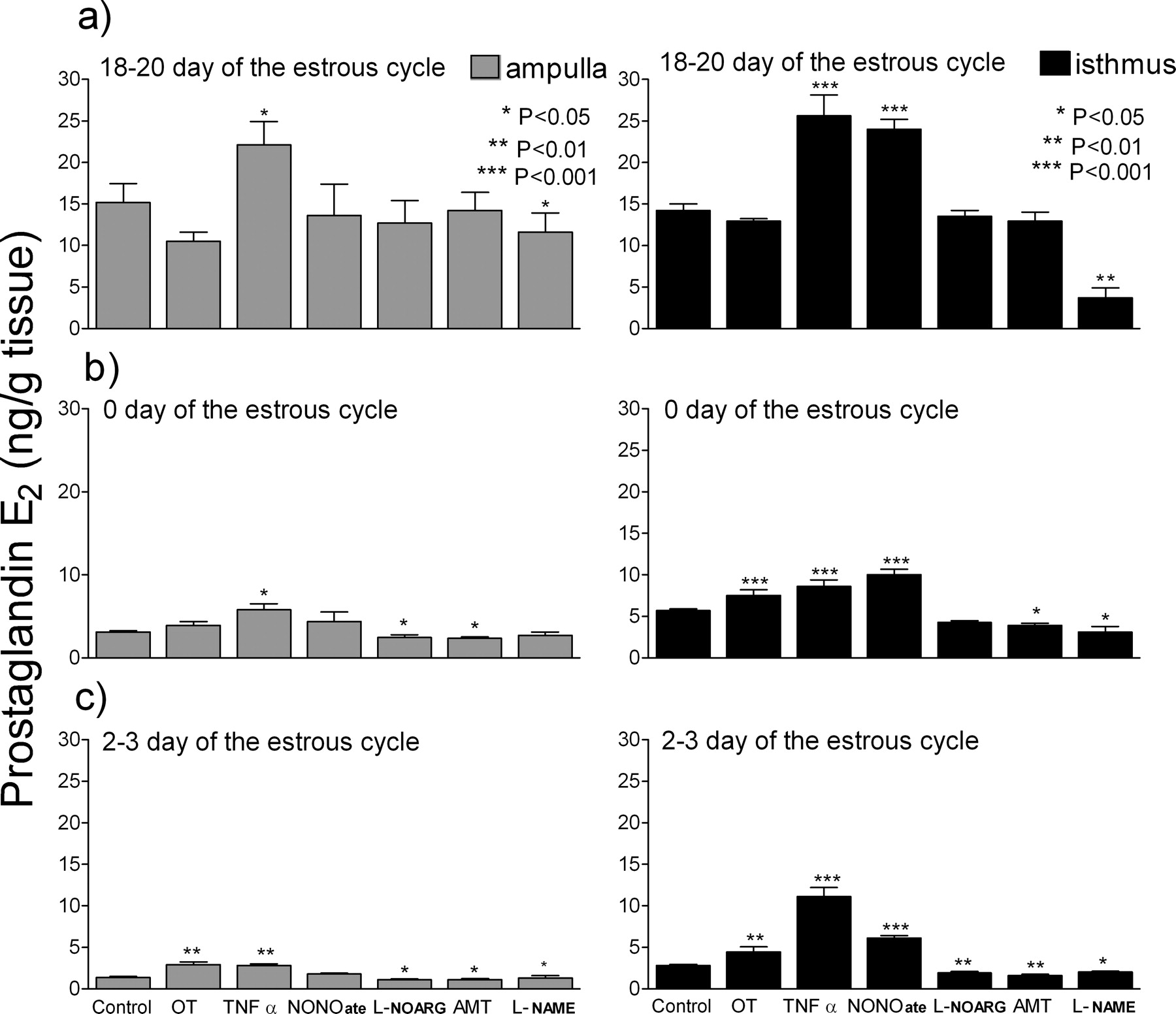
Effects of tumor necrosis factor-α (TNFα; 10 ng/ml); a nitric oxide donor, Spermine Nitric Oxide Complex (NONOate; 10−4 M); a competitive inhibitor of endothelial isoform of NO synthase, Nω-nitro-L-arginine (L-NOARG; 10−4 M); a competitive inhibitor of inducible isoform of NO synthase, (±)-2-amino-5,6-dihydro-6-methyl-4H-1,3-thiazine (AMT; 10−4 M); a nonspecific inhibitor of NO synthase, Nω-nitro-L-arginine methyl ester (L-NAME; 10−4 M); and Oxytocin (OT; 10−7 M) on prostaglandin E2 production in the ampulla and the isthmus at 18–20 Days, 0 Day and 2–3 Days of the oestrus cycle in the bovine oviduct.
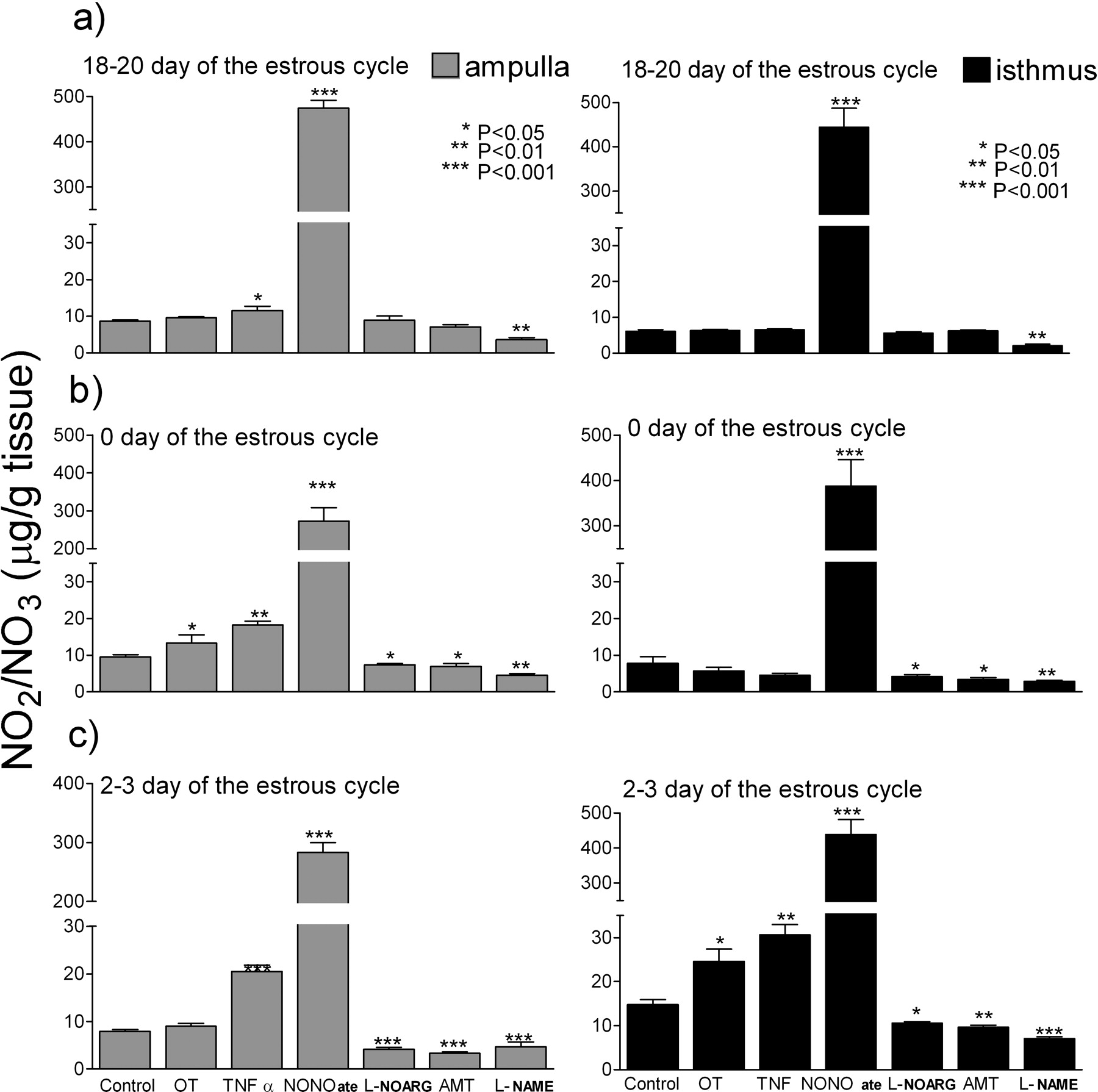
Effects of tumor necrosis factor-α (TNFα; 10 ng/ml); a nitric oxide donor, Spermine Nitric Oxide Complex (NONOate; 10−4 M); a competitive inhibitor of endothelial isoform of NO synthase, Nω-nitro-L-arginine (L-NOARG; 10−4 M); a competitive inhibitor of inducible isoform of NO synthase, (±)-2-amino-5,6-dihydro-6-methyl-4H-1,3-thiazine (AMT; 10−4 M); a nonspecific inhibitor of NO synthase, Nω-nitro-L-arginine methyl ester (L-NAME; 10−4 M); and Oxytocin (OT; 10−7 M) on nitrate (NO2/NO3) production in the ampulla and the isthmus at 18–20 Days, 0 Day and 2–3 Days of the oestrus cycle in the bovine oviduct.
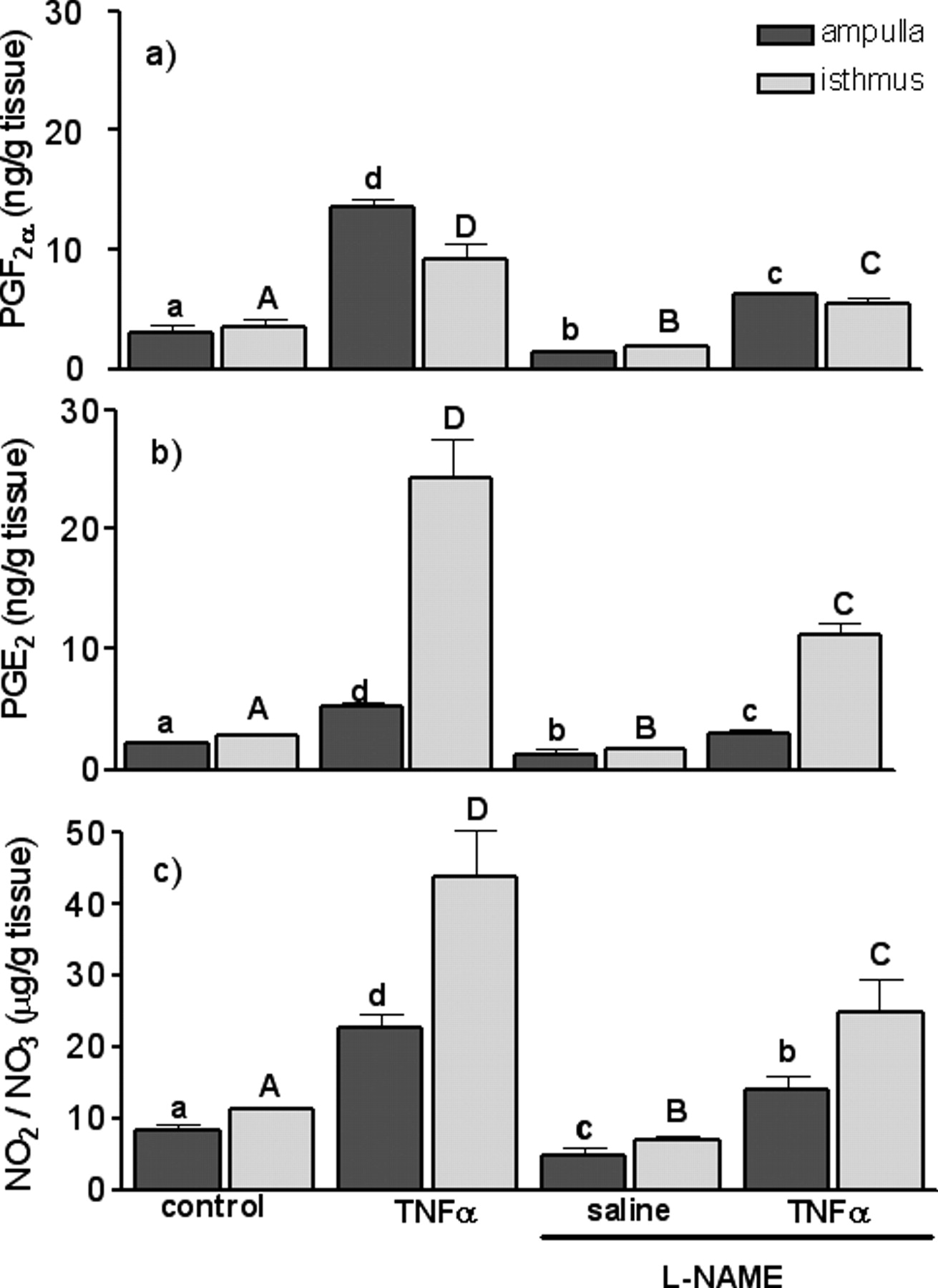
Effects of tumor necrosis factor-α (TNFα; 10 ng/ml) and TNFα (10 ng/ml) together with a non-specific inhibitor of NO synthase, Nω-nitro-L-arginine methyl ester (L-NAME; 10−4 M), on prostaglandin F2α (PGF2α), prostaglandin E2 (PGE2) and nitrate (NO2/NO3) production in the ampulla and the isthmus at 2–3 Days of the estrus cycle in bovine oviduct. Different letters indicate significant differences (P < 0.05) between treatments (lower-case letters, ampulla; capital letters, isthmus).
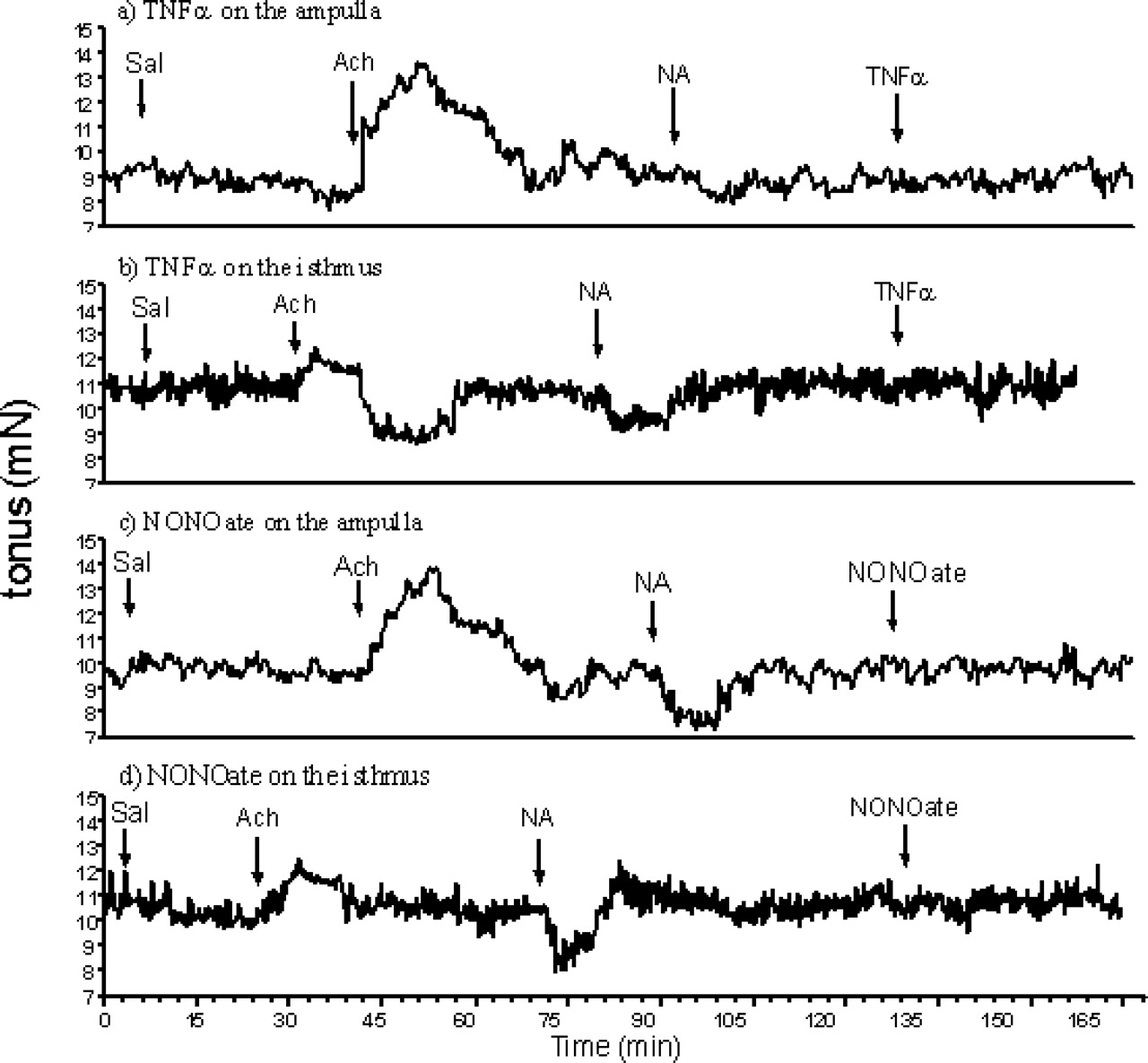
Effects of tumor necrosis factor-α (TNFα; 10 ng/ml; a, b), a nitric oxide donor (NONOate; 10−4 M; c, d), acetylcholine (Ach; 10−6 M) and noradrenalin (NA; 10−6 M) on contractility of ampulla (a, c) and isthmus (b, d) of bovine oviduct. The oviductal explants were collected at Days 2–3 of the cycle. One representative experiment from the six completed.

Effects of prostaglandin E2 (PGE2; 10−6 M; a, b), prostaglandin F2α (PGF2α; 10−6 M; c, d), acetylcholine (Ach) and noradrenalin (NA) on contractility of ampulla (a, c) and isthmus (b, d) of bovine oviduct. The oviductal explants were collected at Days 2–3 of the cycle. One representative experiment from the six completed.
Footnotes
This research was supported by the Polish Academy of Sciences and a joint Polish-Japanese project under the PAS and JSPS agreement. MJS is a guest researcher from the Polish Academy of Sciences supported by the Post-Doctoral Fellowship Program of the Japan Society for the Promotion of Science (No. P 08733). MJS and IW-P were supported by Domestic Grants for Young Scientists of the Foundation for Polish Science (FNP, Program START, 2008 and 2006, respectively).
Acknowledgements
The authors thank Dainippon Pharmaceutical Co., Ltd., Osaka, Japan for recombinant human
TNFα (HF-13), and Stanislaw Brukwald of the animal farm in Rogale and Hanna and Jerzy
Kostuch of the animal farm “Farmer” in Zalesie, Poland, for their excellent cooperation
and permission to use their animals in the study. The authors are grateful to Draminski
Electronics in Agriculture (Olsztyn, Poland, ![]() ) for their
excellent cooperation and the possibility to test and use the USG scanner.
) for their
excellent cooperation and the possibility to test and use the USG scanner.