
Abstract
We examined the capacity of delayed inhibition of plasminogen activator inhibitor-1 (PAI-1) to reduce tubulointerstitial fibrosis induced by unilateral ureteral obstruction (UUO) in mice. Small peptides mimicking parts of urokinase (uPA) and tissular plasminogen activator (tPA) and serving as decoy molecules for PAI-1 were administered daily during the late stages (3 to 8 days) of UUO. Treatment with PAI-1 decoys reduced interstitial deposition of fibronectin, collagen III and collagen IV without changes in macrophage and myofibroblast infiltration. Interestingly, while PAI-1 activity was reduced and the combined uPA and tPA activity was increased, the antifibrotic effect was obtained without modification of plasmin activity but with increased of hepatocyte growth factor (HGF) expression. We show for the first time that treatment with small PAI-1 decoy peptides reduces established tubulointerstitial fibrosis. This protective effect probably resulted from increased degradation of the extracellular matrix by an HGF dependent mechanism.
Introduction
End stage renal disease (ESRD) is preceded, in nearly all cases, by the progressive development of renal fibrosis, corresponding to exaggerated accumulation of extracellular matrix (ECM). Tubulointerstitial fibrosis (TIF), compared to glomerular sclerosis, is strongly correlated with evolution towards ESRD (1). Its development is schematically divided into i) inflammation, macrophage and lymphocyte infiltration and increase in circulating cytokines and chemokines, inducing ii) a disequilibrium between apoptosis and proliferation of tubular cells as well as fibroblast accumulation, followed by iii) secretion of ECM by activated fibroblasts. This increase in fibrotic tissue causes a steady decline of renal function leading to ESRD (1).
The only drugs routinely used in clinical practice to slow down the progression of chronic kidney diseases are the inhibitors of renin/angiotensin system. In addition, there are a limited number of drugs in early phase clinical trials specifically aiming to reduce renal fibrosis. Among the most promising drugs, Pirfenidone, Pentoxifylline, Ruboxistaurin and Rosiglitazone are able to reduce the production of profibrotic cytokines and chemokines (2–5).
In the search for antifibrotic drugs, plasminogen system has drawn attention. Indeed ECM degradation is thought to be largely dependent on this system primarily via the activation of latent matrix metalloproteinases (MMPs) (6). Plasmin is a serine protease which can directly degrade matrix proteins fibronectin, laminin (7) and type IV collagen (8) and activate MMPs (9). Plasmin is generated from plasminogen by two types of plasminogen activators (PAs), uPA and tPA, and the activity of both PAs is inhibited by PAI-1. Because PAI-1 is upregulated in many renal diseases (6), its inhibition should reactivate ECM degradation and thereby induce reversal of fibrosis.
The tubulointerstitial fibrogenic response to UUO is attenuated in PAI-1-knockout mice (10). However in this study, PAI-1 inhibition was established before induction of the pathology. Yet in a clinical context, fibrosis is often detected at a late stage. Therefore the current study was designed to evaluate the effect of delayed PAI-1 blockade on the development of TIF. For this purpose, we used a mixture of small peptides mimicking parts of the protease domains of PAs, thereby serving as PAI-1 decoy molecules which prevent the PAI-induced inhibition of uPA and tPA (11). We explored the efficacy of decoys to reduce pre-installed fibrosis after UUO, a model encompassing the main features of the development of TIF (12).
Materials and Methods
PAI-1 Decoy.
The decoy mixture was composed of two peptides: H-Arg-Asp-Glu-Lys-Thr-Gln-Met-Ile-Tyr-Gln-Gln-OH and H-Asp-Arg-Arg-Leu-Thr-Trp-Glu-Tyr-OH (11), in a 50/50 ratio. Peptides were synthesized by GL Biochem (Shanghai, China) and had an average purity of 97% as assessed by HPLC analysis. Both peptides bound with the kringle-2 domain of PAI-1 and consequently prevented complex formation with tPA and uPA in a reversible manner (11).
Experimental Protocols.
UUO was performed on pathogen free male ED-1 mice (8 weeks) by ligation of the left ureter as previously described (12, 13). To study the effect of delayed PAI-1 inhibition on TIF, the treatment with PAI-1 decoy peptide mix was initiated on day 3 of UUO by daily iv injection of decoy mixture (1 mg/kg/d). Eight days after UUO, mice were sacrificed and the kidneys were removed and sectioned into 4 parts for different analysis: histology (middle section), PAI-1, PAs and plasmin activities (1/2 of a kidney pole), MMPs activities (1/2 of a kidney pole) and HGF expression (kidney pole). To evaluate the level of fibrosis that exists after 3 days of UUO, other mice were killed at 3 days after surgery and histological analysis of TIF was performed.
All experiments reported were conducted as stated in the National Institutes of Health, USPHS, Guide for the Care and Use of Laboratory Animals and were approved by the Institution’s Animal Care and Use Committee.
Immuno-Histochemistry Analysis.
Parts of kidney were fixed in Carnoy’s solution, dehydrated, embedded in paraffin and then sectioned at 3 μm. Then they were subjected to evaluate interstitial fibrosis by immuno-histochemical staining collagen III, collagen IV and fibronectin for ECM detection, F4/80 for macrophages detection and α-smooth muscle actin (α-SMA) for myofibroblasts detection, as previously described (12, 13). Histomorphometric analyses were performed as previously described (12, 13).
PAI-1, PAs and Plasmin Activities.
For protein extraction, parts of kidney were rapidly mixed with 150 μl of ice-cold extraction buffer (13 mM sodium citrate, 0.33 M sodium acetate, pH 3.9) and were manually homogenized on ice for 10 sec using a potter device (Piston Pellet 1.5 ml, Treff AG, Switzerland) followed by centrifugation for 5 min at 12000 × g at 4°C. Supernatant (100 μl) was added to 5 μl of 20% acetic acid, gently mixed and stored at −80°C for PAs and plasmin activities determination or it was directly stored at −80°C for PAI-1 activity. Protein concentrations were measured using a Bradford assay (Bio-Rad Laboratories, France). After dilution (1/20) of samples in 0.9% of NaCl, PAs and PAI-1 activities were measured by chromogenic assay (101101 Spectrolyse®/Fibrin, Biopool Int., Sweden and BI101201 Spectrolyse (pl) PAI, Kordia, the Netherlands, respectively). Plasmin activity was determined using a highly selective chromogenic substrate for plasmin (S-2403, Chromogenix-Instrumentation Laboratory SpA, France).
MMP-2 and MMP-9 Activities.
Renal proteins were extracted as described for PAI-1 activity excepted that the extraction buffer contained 10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA. MMP-2 and MMP-9 activities were determined by zymography. For this, 20 μl of samples were mixed with non-reducing SDS sample buffer and loaded onto 8–10% SDS-polyacrylamide gels containing 1 mg/ml of gelatin. After electrophoresis, gels were soaked 2 × 20 min in 2.5% (vol/vol) of Triton X100 at room temperature and incubated overnight at 37°C in activation buffer (10 mM Tris-HCl pH 7.5, 1.25% (v/v) Triton X100, 5 mM CaCl2 and 1 μM ZnCl2). Gels were then stained for 3 h with 0.25% (w/v) Coomassie Brilliant Blue in a mixture of methanol/acetic acid/water (50/10/40 v/v/v) and destained in the same mixture without Coomassie Brilliant Blue.
HGF Expression.
Renal proteins were extracted as described for MMPs zymography excepted that the extraction buffer was supplemented with SDS (0.1%), NP-40 (1%), PMSF (0.36 mg/ml), aprotein and leupeptin (10 μg/ml). Then HGF expression was assayed in samples using immunoassay kit (DHG00, R&D Systems, MN, USA).
Statistical Analysis.
All the data are expressed as mean ± standard deviations (SD). Statistical analysis was performed using Prism software. One-Way ANOVA with Bonferroni post-test were performed for comparison between the different groups. P < 0.05 was considered statistically significant.
Results
PAI-1 Decoy Treatment Was Initiated at Day 3 of UUO, when TIF Is Already Detectable.
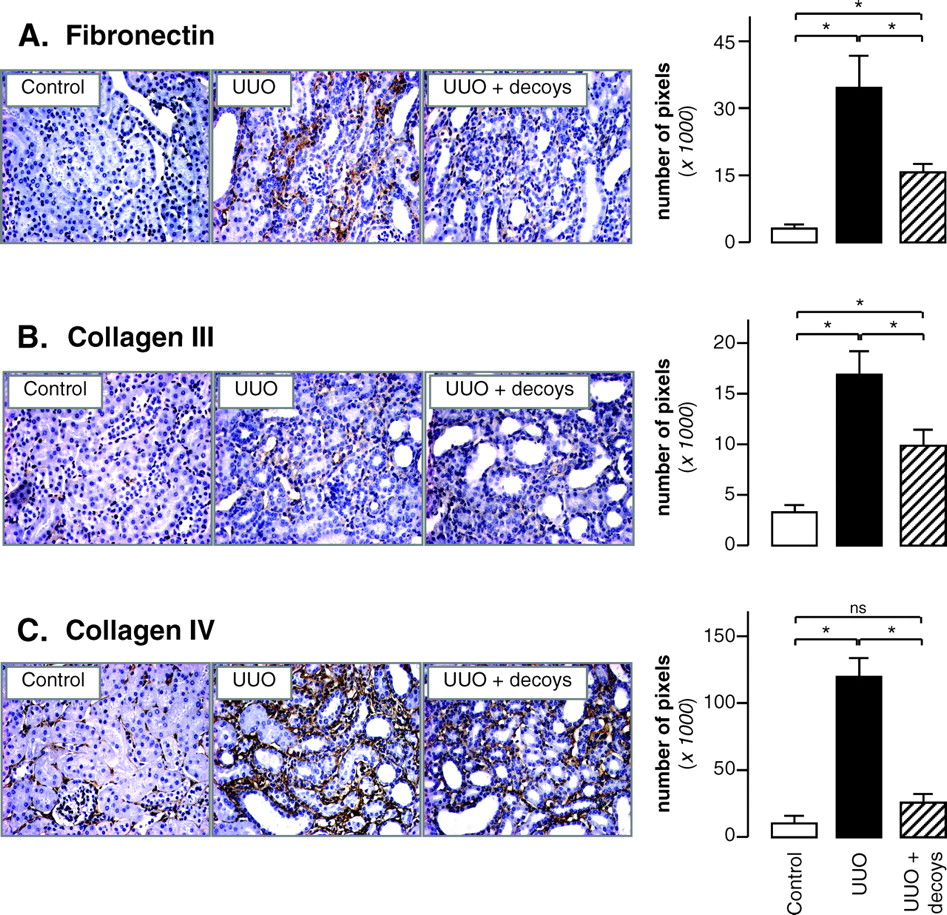
The objective of this study was to examine the efficacy of PAI-1 decoys to reduce UUO-induced pre-installed TIF. Consequently, to choose when to begin the administration of PAI-1 decoys, a time course of TIF appearance in the obstructed kidneys was first undertaken. TIF is characterized by ECM deposition which is the result of the elevated synthesis and decreased degradation of ECM compounds, including fibronectin. As presented in Figure 1, fibronectin accumulation increased in accordance with the duration of UUO. Moreover, renal fibronectin levels are slightly, but significantly, increased after 3 days of UUO. This result, with those from other laboratories (12, 14–19), demonstrates that 3 days of obstruction appeared sufficient to induce a moderate tubulointerstitial fibrosis. So in the following experiments, PAI-1 decoy treatment was initiated at day 3 of UUO and its therapeutic potential to treat established TIF was evaluated.
PAI-1 Inhibition Reduces the Severity of Fibrosis.
To test the effect of delayed PAI-1 decoy treatment on the development of TIF, we quantified renal content of fibronectin in 8d-UUO mice receiving or not PAI-1 decoy injections. As shown in Figure 2A, the increase of fibronectin accumulation induced by 8 days of UUO was significantly attenuated by PAI-1 decoys. Despite this attenuation, fibronectin levels were however higher than those measured in 3d-UUO (Fig. 1), when treatment started, suggesting that decoys decrease—but do not stop—ECM deposition. To confirm the protective effect of PAI-1 decoys, immuno-histochemical staining of collagen III and collagen IV, two other ECM proteins, was also realized. As for fibronectin, the 8d-UUO increased the expression of both collagens and this effect was partially blocked by decoy administration (Fig. 2B–C). Thus, delayed PAI-1 inhibition impairs UUO-induced TIF, making the decoy peptides attractive candidates to slow down the progression of chronic nephropathies.
Interstitial Macrophage and Myofibroblast Accumulation Is Not Affected by PAI-1 Inhibition.
In parallel with ECM deposition, infiltration of macrophages and myofibroblasts is observed in interstitium of the UUO kidney. Myofibroblasts are principal cell type involved in the production of matrix proteins. Macrophages participate in the activation of fibroblasts into myofibroblasts via cytokines secretion and can be chemo-attracted to PAI-1 (10). Therefore, we studied whether PAI-1 inhibition could modify macrophage and myofibroblast recruitment. However, no effect was observed (Fig. 3A–B). These data suggest that in the UUO kidney, delayed PAI-1 decoy treatment exerts its protective effect by promoting the ECM degradation rather than modulating cells involved in ECM synthesis.
PAI-1 Decoys Increase PAs Activity but Not Plasmin Activity.
PAI-1 plays an important role in the degradation of ECM proteins via its effect on PAs which themselves activate plasminogen into plasmin. So we studied the effect of PAI-1 decoy treatment on the activity of these enzymes. PAI-1 is normally not produced in healthy kidneys but its expression is strongly induced by ureteral obstruction (10). In addition, the enzymatic activities of tPA, uPA and plasmin are increased in UUO, despite a raise in PAI-1 (10). In our study, 8d-UUO stimulated PAI-1 and plasmin activities but did not change PAs activity (Fig. 4A–C). Since PAI-1 inhibits PAs and PAs activates plasmin, it might be expected that the treatment of UUO mice with PAI-1 decoys would result in an increase of PAs activity and in a more important increase of plasmin activity. As expected, PAI-1 activity was reduced (Fig. 4A) and PAs activity was stimulated (Fig. 4B) in UUO mice receiving decoys compared to UUO alone. However, the UUO-induced stimulation of plasmin activity was unaffected by decoy administration (Fig. 4C). These results suggest that PAI-1 inhibition attenuated TIF by a mechanism depending mainly on PAs.
PAI-1 Inhibition Has No Effect on MMPs Activity.
MMPs are a large family of matrix-degrading enzymes. Among them are MMP-2 and MMP-9, two gelatinases highly expressed in renal tissue that can be stimulated by PAs (20, 21). In renal mesangial cells, uPA activates MMP-2 via membrane type 1-MMP, independently of plasmin (21). In addition, uPA-induced activation of MMP-9 in monocytes is partially blocked by PAI-1 activation but is not affected by plasmin inhibitors (20). Since PAI-1 decoys increased PAs activity without change in plasmin activity in UUO kidney, it is therefore possible that they also stimulated both MMP-2 and MMP-9. So we tested this hypothesis using gelatin zymography. As shown in Figure 4D–E, MMP-2 and MMP-9 activities were raised in 8d-UUO but they were not significantly modified by PAI-1 decoys administration. Thus, antifibrotic effect of delayed PAI-1 inhibition was independent of MMP-2 and MMP-9 activities.
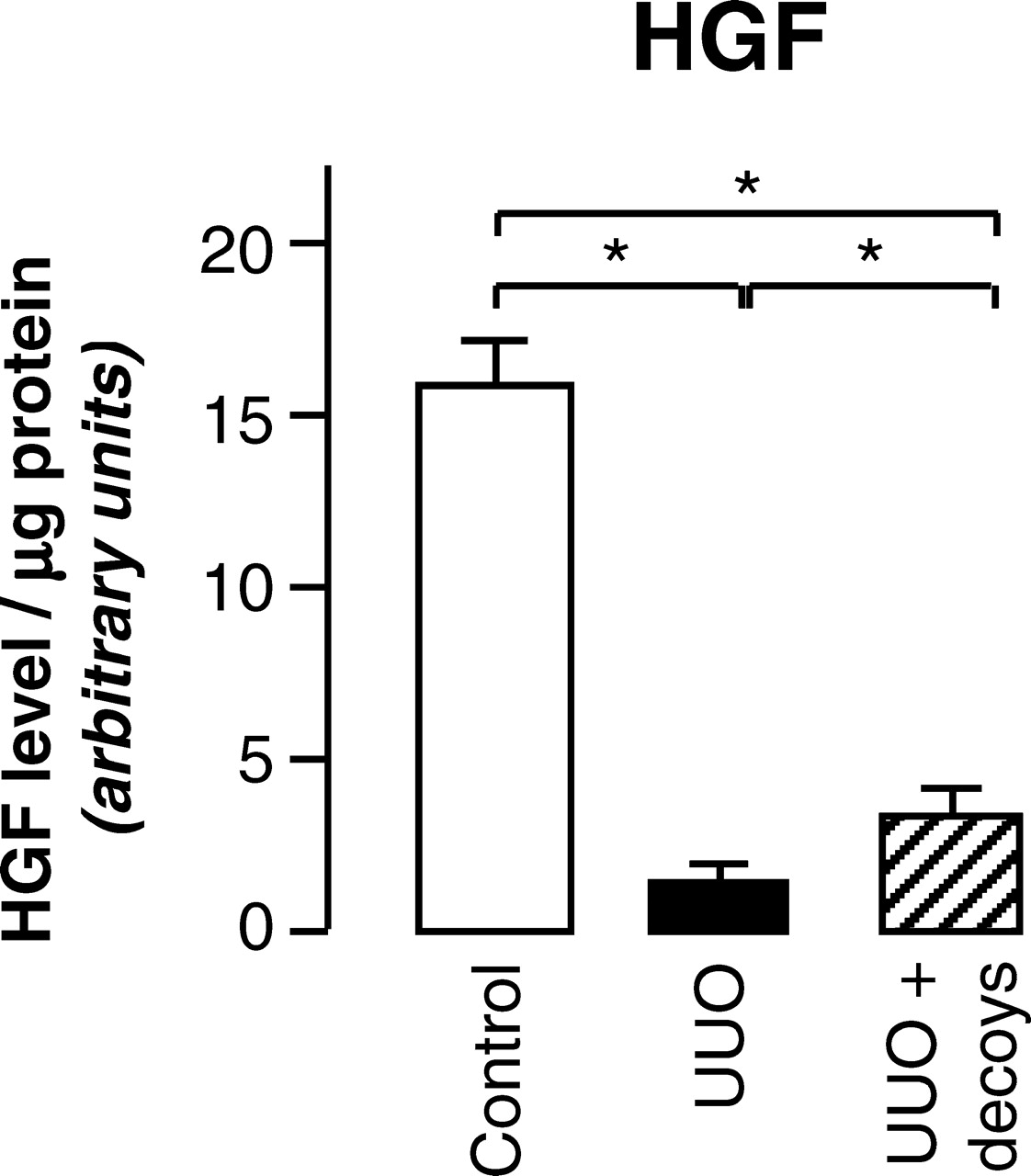
HGF Protein Increases upon PAI-1 Decoy Administration.
TGF-β plays an important role in ECM deposition and fibrotic changes. However, PAI-1 decoys administration did not modify TGF-β mRNA expression in UUO (data not shown), suggesting that another mediator accounts for the antifibrotic effect of decoy peptides. HGF is a putative candidate because i) HGF administration ameliorates TIF in UUO mice (22) and ii) activation of HGF is observed concomitantly with attenuation of fibrotic lesions in the liver of PAI-1 knockout mice with bile duct ligation (23). The involvement of HGF in protective effect of PAI-1 decoys was thus examined measuring its protein expression by ELISA. As presented in Figure 5, 8d-UUO caused a decrease in HGF expression and this effect was attenuated by PAI-1 decoys. Similar results were obtained with HGF immunoblot (not shown). Thus it is reasonable to think that part of the renal protective effect of delayed PAI-1 inhibition goes through increased HGF expression.
Discussion
In the current study we show that the delayed inhibition of PAI-1 by daily i.v. administration of a mixture of two small PAI-1 decoy peptides efficiently reduced UUO-induced TIF. This effect was obtained without modification of plasmin activity but accompanied by increased HGF level.
Early blockade of PAI-1 expression prevented the development of fibrosis in UUO and experimental glomerulonephritis (10, 24). However, the effects of a drug differ if administration begins before or after disease induction (25). Therefore, the current study is the first example showing that delayed inhibition of PAI-1 reduced established TIF. This result is especially important in the context of renal disease which is often only detected at a later stage. In addition, it shows the feasibility of the use of small PAI-1 decoy peptides for antifibrotic treatment. The attenuation of UUO induced-TIF by PAI-1 decoys administration is accompanied by stimulation of PAs activity. This result contrasts with previous studies where uPA−/− or tPA−/− mice subjected to UUO had similar and lower TIF than normal mice, respectively (26, 27). It can be explained by different experimental conditions since, in our study, the inhibition of PAI-1 was delayed and led to activation of both uPA and tPA.
PAI-1 decoys do not change the number of interstitial macrophages and myofibroblasts. This result contrasts with studies using PAI-1 knockouts (10) or transgenic PAI-1 overexpressing mice (14) and with experiments using a dominant negative PAI-1 (24). However, induction of TIF by PAI-1 involves early effects on interstitial cell recruitment while the more delayed effects are associated with decreased uPA activity (14). In addition, in the previous studies (10, 14), the expression of the entire PAI-1 molecule, and consequently related effects, are modified while only the PAI-1 ability to inhibit PAs is blocked in our study. PAI-1 can directly interact with other proteins such as vitronectin and low density lipoprotein receptor-related protein (LRP) to regulate cellular adhesion and migration and have monocyte chemoattractant properties (6). These interactions being probably not modified by PAI-1 decoys might also explain the lack of effect on cell recruitment in our model.
Although PAI-1 decoy administration restored PAs activity, no effect on plasmin activity was observed. The absence of correlation between both activities has already been described in PAI-1−/− or tPA−/− mice subjected to UUO (10, 23, 27). Therefore it is possible that the antifibrotic effects of PAI-1 decoys involve plasmin-independent pathways. The absence of effect of PAI-1 inhibition on renal MMP-2 and MMP-9 activities confirms this hypothesis. Other proteolytic activities of PAs can be proposed. First, increased PAI-1 decoys-induced PAs activity may be directly involved in proteolysis because some ECM proteins, such as fibronectin, are direct substrates of PAs (28). Second, increased PAs levels might activate MMPs other than MMP-2 and MMP-9. Indeed, PAs can activate membrane type 1-MMP (21) which digests collagen and fibronectin (29). Finally, it was shown that the uPA receptor (uPAR) can dimerize with other receptors including LRP, an endocytosis receptor for fibronectin (30). So it is not excluded that the decoys-induced increase of PAs activity stimulates the uPAR/LRP pathway of endocytosis, thus promoting decreased ECM expression.
Even if TGF-β plays an important role in fibrogenic mechanisms, we did not observe modification of expression of this profibrotic cytokine. TGF-β is therefore probably not involved in the beneficial effects of PAI-1 decoys administration. HGF is a renotrophic protein which delays the progression of established TIF in the UUO model (16). Because PAI-1 decoys increased by 75% the HGF protein levels compared to non-treated UUO, their antifibrotic effect might also be mediated by HGF. How decoys treatment activated HGF was not investigated. However, in vitro, PAs can directly cleave pro-HGF into HGF (31). In addition, renal HGF expression decreased after UUO in uPAR knockout mice, independently of plasminogen (32). So PAI-1 decoy peptides could increase HGF levels by a mechanism dependent on PAs and uPAR. It is possible that decoys increase uPAR expression at cell surface. Indeed, PAI-1 can bind to uPAR-bound uPA and the ternary uPAR/ uPA/PAI-1 complex is known to rapidly associate with LRP to be internalized (6). As PAI-1 decoys prevent interaction between PAI-1 and PAs, they may block the intracellular sequestration of the uPAR/uPA/PAI-1 complex, leading therefore to maintain uPAR at cell surface and thus increase PAs activity.
In conclusion, we have shown the efficacy/feasibility to use small peptides inhibiting PAI-1 activity to slow down the progression of tubulointerstitial fibrosis after induction of the pathology. This is a situation more realistic than treatment before initiation of the pathology as it resembles more to what occurs in a clinical setting. Although the exact mechanism is not clear, HGF activation might be responsible for the antifibrotic effects of PAI-1 decoys.
UUO induces tubulointerstitial fibrosis. Renal accumulation of fibronectin was quantified by immuno-histochemistry in mice before UUO (day 0) and after 3 days or 8 days of UUO. Data are expressed as means ± SD from 5 mice. * P < 0.05 versus day 0, # P < 0.05 versus day 3. PAI-1 decoy peptides decrease ECM accumulation. Renal accumulation of fibronectin (A), collagen III (B) and collagen IV (C) was quantified by immuno-histochemistry in control mice (control) and in 8d-UUO mice treated (UUO +decoys) or not (UUO) by PAI-1 decoys. Data are expressed as means ± SD from 5 mice and pictures display corresponding light micrographs. * P < 0.05, ns: not significant. Infiltration of macrophages and myofibroblasts is not modified by PAI-1 decoys. Renal accumulation of macrophages (A) and myofibroblasts (B) was quantified by anti F4/80 and anti-α-SMA immuno-histochemistry, respectively, in control mice (control) and in 8d-UUO mice receiving (UUO + decoys) or not (UUO) PAI-1 decoys. Data are expressed as means ± SD from 5 mice. Pictures display corresponding light micrographs. * P < 0.05, ns: not significant. Effect of inhibition of PAI-1 on renal plasminogen system. Renal activity of PAI-1 (A), plasminogen activator (B), plasmin (C), MMP-2 (D) and MMP-9 (E) was measured in control mice (control) and in 8d-UUO mice treated (UUO+decoys) or not (UUO) by PAI-1 decoys. Data are expressed as means ± SD from 5 mice. * P < 0.05, ns: not significant. PAI-1 blockade partially restores HGF expression. Renal HGF expression was analyzed by ELISA in control mice (control), 8d-UUO mice (UUO) or UUO mice supplemented with PAI-1 decoys (UUO + decoys). Results represent the mean ± SD from 5 experiments. * P < 0.05.


Footnotes
J. Gonzalez, J. Klein, R. Chaaya and M. Miravete received a grant from the “Ministère de l’Education Nationale de la Recherche et de la Technologie.” S.D. Chauhan was a recipient of grants from both the “Entente Cordiale” and the “Fondation de la Recherche Médicale.”
1
These authors contributed equally to this work.