Microscopic tridomain model of electrical activity in the heart with dynamical gap junctions. Part 2 – Derivation of the macroscopic tridomain model by unfolding homogenization method
Available accessResearch articleFirst published online April 4, 2023
Microscopic tridomain model of electrical activity in the heart with dynamical gap junctions. Part 2 – Derivation of the macroscopic tridomain model by unfolding homogenization method
We study the homogenization of a novel microscopic tridomain system, allowing for a more detailed analysis of the properties of cardiac conduction than the classical bidomain and monodomain models. In (Acta Appl.Math.179 (2022) 1–35), we detail this model in which gap junctions are considered as the connections between adjacent cells in cardiac muscle and could serve as alternative or supporting pathways for cell-to-cell electrical signal propagation. Departing from this microscopic cellular model, we apply the periodic unfolding method to derive the macroscopic tridomain model. Several difficulties prevent the application of unfolding homogenization results, including the degenerate temporal structure of the tridomain equations and a nonlinear dynamic boundary condition on the cellular membrane. To prove the convergence of the nonlinear terms, especially those defined on the microscopic interface, we use the boundary unfolding operator and a Kolmogorov–Riesz compactness’s result.
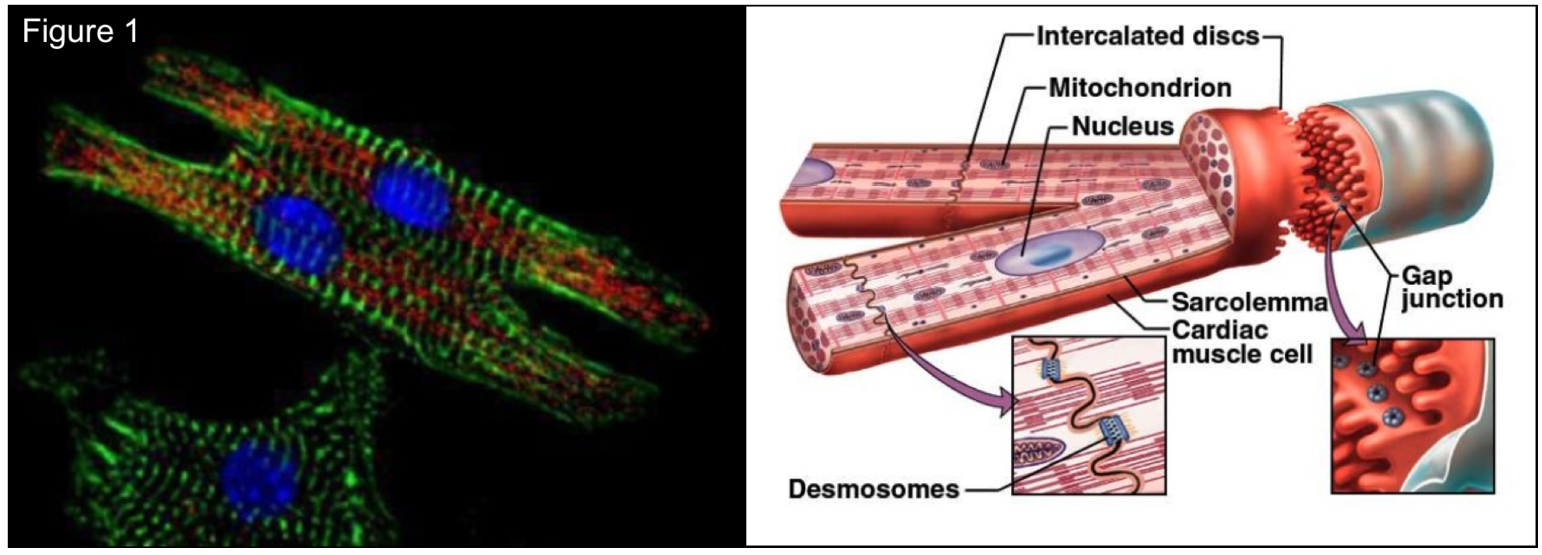
The conduction of electrical waves in cardiac tissue is key to human life, as the synchronized contraction of the cardiac muscle is controlled by electrical impulses that travel in a coordinated manner throughout the heart chambers. Under pathological conditions cardiac conduction can be severely reduced, potentially leading to reentrant arrhythmias and ultimately death if normal propagation is not restored properly. At a sub-cellular level, electrical communication in cardiac tissue occurs by means of a rapid flow of ions moving through the cytoplasm of cardiac cells, and a slower inter-cellular flow mediated by gap junctions embedded in the intercalated discs (see Fig. 1). Gap junctions are inter-cellular channels composed by hemichannels of specialized proteins, known as connexions, that control the passage of ions between neighboring cells.
Representation of the cardiomyocyte structure.
Starting from a more accurate microscopic (cell-level) model of cardiac tissue, with the heterogeneity of the underlying cellular geometry represented in great detail, it is possible to derive the macroscopic tridomain model (tissue-level) using the homogenization method. The microscopic tridomain model consists of three quasi-static equations, two for the electrical potential in the intracellular medium and one for the extracellular medium, coupled by ordinary differential equations describing the dynamics of the ions channels at each membrane (the sarcolemma) and at gap junctions. These equations depend on scaling parameter ε whose is the ratio of the microscopic scale from the macroscopic one. The microscopic tridomain model was proposed three years ago [15,20] in the case of just two coupled cells. Recently, we have extended in [5] this microscopic tridomain model to larger collections of cells. Further, we have established the well-posedness of this problem and proved the existence and uniqueness of their solutions based on Faedo–Galerkin method.
The macroscopic tridomain model is used as a quantitative description of the electric activity in cardiac tissue with dynamical gap junctions. The relevant unknowns are the two intracellular for and extracellular potentials, along with the so-called transmembrane potential for and the so-called gap potential . In this model, the intra- and extracellular spaces are considered at macro-scale as two separate homogeneous domains superimposed on the cardiac domain. Conduction of electrical signals in cardiac tissue relies on the flow of ions through cell membrane and gap junctions. Each intracellular domain and extracellular one are separated by the cell membrane while the two intracellular domains are connected by gap junctions (see Fig. 2). The macroscopic tridomain model can be viewed as a PDE system consisting of three degenerate reaction-diffusion equations involving the unknowns (, , ). These equations are supplemented by a ODE system for the dynamics of the ion channels through the cell membrane (involving the gating variable for ).
(Left) Periodic heterogeneous domain Ω. (Right) Reference cell Y at ε-structural level.
Regarding the classical bidomain model in the literature, there are formal and rigorous mathematical derivations of the macroscopic model from a microscopic description of heart tissue. From a mathematical point of view, Krassowska et al. [17] applied the two-scale method to formally obtain this macroscopic model (see also [1,13] for different approaches). Furthermore, Pennachio et al. [19] used the tools of the Γ-convergence method to obtain a rigorous mathematical form of this homogenized macroscopic model. Amar et al. [2] studied a hierarchy of electrical conduction problems in biological tissues via two-scale convergence. While, the authors in [6] proved the existence and uniqueness of solution of the microscopic bidomain model based on Faedo–Galerkin technique. Further, they used the periodic unfolding method at two scales to show that the solution of the microscopic biodmain model converges to the solution of the macroscopic one. Recently, we have developed the meso-microscopic bidomain model by taking account three different scales and derived a new approach of its macroscopic model using two different homogenization methods. The first method [3] is a formal and intuitive method based on a new three-scale asymptotic expansion method applied to the meso- and microscopic model. The second one [4] based on unfolding operators which not only derive the homogenized equation but also prove the convergence and rigorously justify the mathematical writing of the preceding asymptotic expansion method.
The main contribution of our paper is to provide a simple homogenization proof that can handle some relevant nonlinear membrane models (the FitzHugh-Nagumo model), relying only on unfolding operators. More precisely, we show that the solution constructed in the microscopic tridomain problem converge to the solution of the macroscopic (homogenized) tridomain model. So, we will derive the homogenized tridomain model of cardiac electro-physiology from the microscopic one using the periodic unfolding technique. The latter method not only makes it possible to derive the homogenized equation but also to prove the convergence and to rigorously justify the mathematical writing of the preceding formal method. The homogenization method that we propose allows us to investigate the effective properties of the cardiac tissue at each structural level, namely, micro-macro scales.
The paper is organized as follows: Section 2 is devoted to the geometrical setting and to the introduction of the microscopic tridomain problem. In Section 3, we state our main homogenization results. Next, some notations and properties on the domain and boundary unfolding operators are introduced in Section 4. Finally, Section 5 is devoted to homogenization procedure based on unfolding operators.
Tridomain modeling of the heart tissue
The aim of this section is to describe the geometry of the cardiac tissue and to present the microscopic tridomain model of the heart.
Geometrical setting of heart tissue
Let Ω be an open connected bounded subset of , . The typical periodic geometrical setting is displayed in Fig. 2.
Let be a small positive parameter, related to the characteristic dimension of the micro-structure and which takes values in a sequence of strictly positive numbers tending to zero. Under the one-level scaling, the characteristic length is related to a given macroscopic length L (of the cardiac fibers), such that the scaling parameter ε introduced by:
From the biological point of view, the cardiac cells are connected by many gap junctions. Therefore, geometrically, Ω represents the region occupied by the cardiac tissue and consists of two intracellular media for , that are connected by gap junctions and extracellular medium (for more details see [15,20]). Each intracellular medium and the extracellular one are separated by the surface membrane (the sarcolemma) which is expressed by:
while the remaining (exterior) boundary is denoted by . We can consider that the intracellular zone as a perforated domain obtained from Ω by removing the holes which correspond to the extracellular domain .
We can divide Ω into small elementary cells , with are positive numbers. These small cells are all equal, thanks to a translation and scaling by ε, to the same reference cell of periodicity called the reference cell . So, the ε-dilation of the reference cell Y is defined as the following shifted set :
where represents the translation of with and .
Therefore, for each macroscopic variable x that belongs to Ω, we define the corresponding microscopic variable that belongs to Y with a translation. Indeed, we have:
Since we will study the behavior of the functions which are -periodic, by periodicity we have . By construction, we say that belongs to Y.
We are assuming that the cells are periodically organized as a regular network of interconnected cylinders at the microscale. The microscopic reference cell Y is also divided into three disjoint connected parts: two intracellular parts for , that are connected by an intercalated disc (gap junction) and extracellular part . Each intracellular parts and the extracellular one are separated by a common boundary for . So, we have:
with . In a similar way, we can write the corresponding common periodic boundary as follows:
with denote the same previous translation, and for .
In summary, the intracellular and extracellular media can be described as follows:
where , and , are respectively defined as (1)–(2) for .
Microscopic tridomain model
The electric properties of the tissue at cellular level are described by the intracellular for and extracellular , potentials respectively with the associated conductivities and . In [5], we presented and studied in details the non-dimensional tridomain model with respect the scaling parameter ε, as well as the models chosen for the membrane and gap junctions dynamics. More precisely, we consider the following microscopic tridomain model:
with and each equation corresponds to the following sense: (4a) Intra quasi-stationary conduction, (4b) Extra quasi-stationary conduction, (4c) Transmembrane potential, (4d) Continuity equation at cell membrane, (4e) Reaction condition at the corresponding cell membrane, (4f) Dynamic coupling, (4g) Gap junction potential, (4h) Continuity equation at gap junction, (4e) Reaction condition at gap junction.
Observe that the tridomain equations (4a)–(4b) are invariant with respect to the scaling parameter ε. As usual in homogenization theory, the electrical potentials are assumed to have the following form
where each function depends on time , slow (macroscopic) variable x and the fast (microscopic) variable . Similarly, the transmembrane potential , the gap junction potential and the corresponding gating variable for have the same previous form. Furthermore, the conductivity tensors are considered symmetric and dependent both on the slow and fast variables, i.e. for , we have
satisfying the elliptic and periodicity conditions: there exist constants , such that and for all :
We complete system (4) with no-flux boundary conditions on :
where and n is the outward reference normal to the exterior boundary of Ω. We impose initial conditions on transmembrane potential , gap junction potential and gating variable as follows:
with .
Next, we introduce some assumptions on the ionic functions, the source term and the initial data.
Assumptions on the ionic functions. The ionic current at each cell membrane can be decomposed into and , where with . Furthermore, the nonlinear function is considered as a function and the functions and are considered as linear functions. Also, we assume that there exists and constants and , such that:
with for .
Now, we represent the gap junction between intra-neighboring cells by a passive membrane:
where is the conductance of the gap junctions. A discussion of the modeling of the gap junctions is given in [14].
Assumptions on the source term. There exists a constant C independent of ε such that the source term satisfies the following estimation for :
Assumptions on the initial data. The initial condition , and satisfy the following estimation:
for some constant C independent of ε. Moreover, , and are assumed to be traces of uniformly bounded sequences in with .
Finally, we observe that the equations in (4) are invariant under the change of , and into , , for any . Therefore, we may impose the following normalization condition:
Main results
In this part, we highlight the main results obtained in our paper. Based on the a priori estimates and unfolding homogenization method, we can pass to the limit in the microscopic equations and derive the following homogenized problem:
(Macroscopic Tridomain Model).
Assume that conditions (6)–(
12
) hold. Then, a sequence of solutionsof the microscopic tridomain model (4) converges asto a weak solutionsatisfying the following conditions:
(Algebraic relation).
(Regularity).
(Initial conditions).
(Boundary conditions).
(Differential equations).
where,, (resp.) is the ratio between the surface membrane (resp. the gap junction) and the volume of the reference cell. Furthermore,nrepresents the outward reference normal to the boundary of Ω. Herein, the homogenized conductivity matricesforare respectively defined by:where the componentsofforare respectively the corrector functions, solutions of the ε-cell problems:for,, the standard canonical basis in.
The proof of Theorem 1 is proved rigorously in Section 4.2 using unfolding homogenization method. The uniqueness of the solutions to the macroscopic model can be proved similar as that of the microscopic model with minor changes (see [5]). This implies that all the convergence results remain valid for the whole sequence. Furthermore, it is easy to verify that the macroscopic conductivity tensors of the intracellular and extracellular spaces are symmetric and positive definite (see Remark 6).
The authors in [6] treated the microscopic bidomain problem where the gap junction is ignored. They considered that there are only two intra- and extracellular media separated by a single membrane (sarcolemma). Comparing to [6], the microscopic tridomain model in our work consists of three elliptic equations coupled through three boundary conditions, two on each cell membrane and one on the gap junction which separates between two intracellular media. The macroscopic tridomain model is more general and complex than the classical monodomain and bidomain models. Using periodic unfolding homogenization method, we derive a new approach of the homogenized model (13) from the microscopic tridomain problem (4).
Regarding the classical bidomain model [6,9], we can derive this model from our tridomain problem if we take .
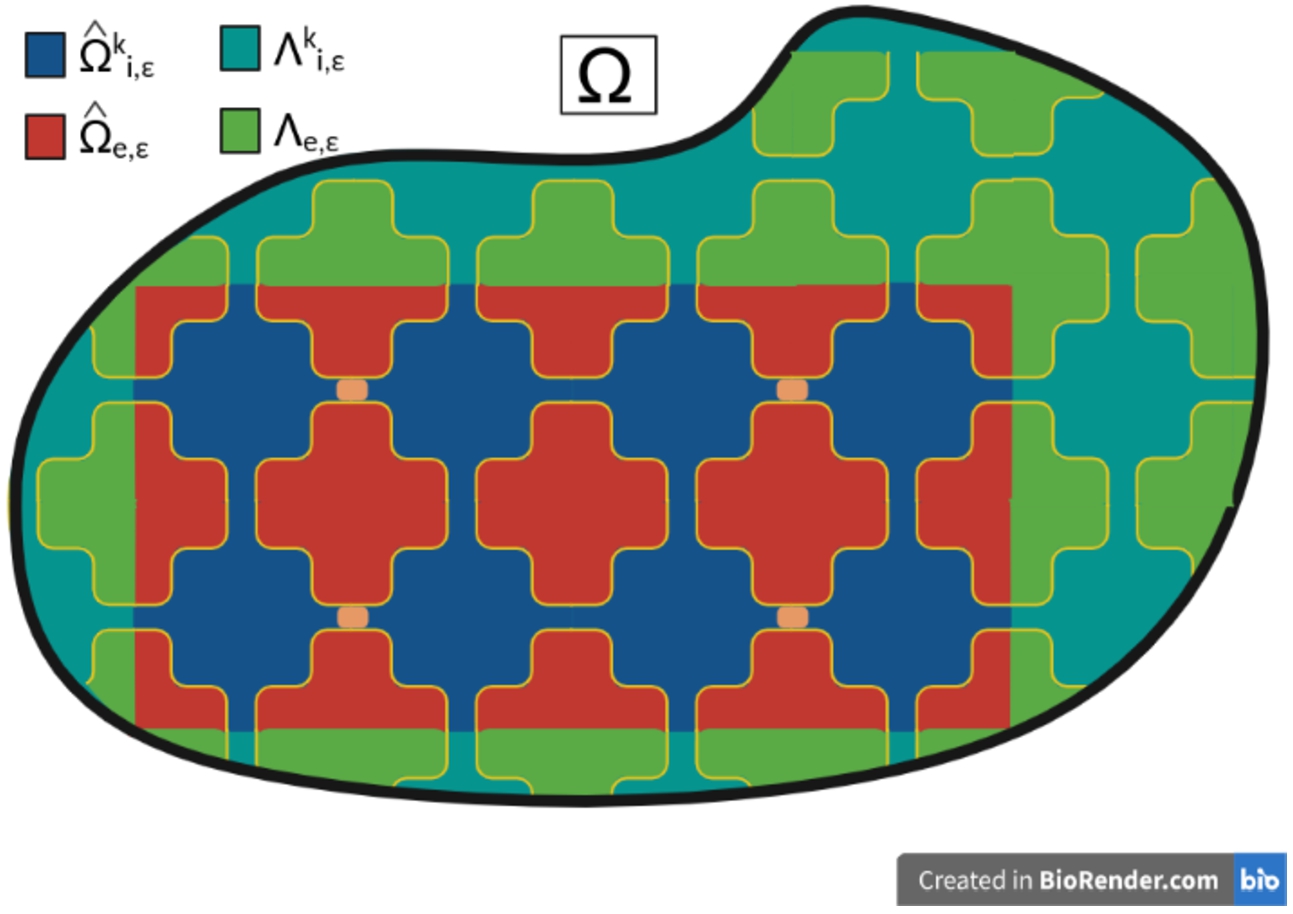
The sets for (in blue), (in red), (in dark cyan) and (in green).
Time-depending unfolding operators
Unfolding operator and some basic properties
Under the notation (3), we begin with introducing the unfolding operator and describe some of its properties. For more properties and proofs, we refer to [7,8]. First, we present the unfolding operators defined for perforated domains on the domain . Then we define boundary unfolding operators one on the membrane , and the other on the gap junction .
In order to define an unfolding operator, we first introduce the following sets in (see Fig. 3)
,
interior ,
interior ,
interior ,
, ,
,
,
,
, , ,
,
where . For all , let be the unique integer combination of the periods such that . We may write for all , so that for all , we get the unique decomposition:
Based on this decomposition, we define the unfolding operator in intra- and extracellular domains.
For any function ϕ Lebesgue-measurable on the intracellular medium for , the unfolding operator is defined as follows:
where denotes the Gauβ-bracket. Similarly, we define the unfolding operator on the domain . We readily have that:
For any function φ Lebesgue-measurable on the membrane for , the boundary unfolding operator is defined as follows:
Similarly, we define the boundary unfolding operator on the gap junction .
Properties of the unfolding operator
In the following proposition, we state some basic properties of the unfolding operator which will be used frequently in the next sections.
(Some properties of the unfolding operator [7,8]).
The operatorandare linear and continuous forand. Similarly, we have the same properties for the unfolding operatorand for the boundary unfolding operator.
Forand, it holds thatand, withand.
For,, we have
For, withand. Then we have
Let, withand. Ifstrongly inas, then
For,, it holds thatwith.
The unfolding operators and for are related in the following sense:
for and a.e. . In particular, by the standard trace theorem in , there is a constant C independent of ε and t such that
From the properties of in Proposition 1, it follows that
Similarly, the trace theorem in holds for (which can be found as Remark 4.2 in [7]).
In the sequel, we will define the periodic Sobolev space as follows:
Let be a reference cell and . Then, we define
where . Its duality bracket is defined by
Furthermore, by the Poincaré-Wirtinger’s inequality, the Banach space has the following norm:
We denote by for .
Microscopic tridomain model
We start by stating the weak formulation of the microscopic tridomain model as given in the following definition.
(Weak formulation of microscopic system).
A weak solution to problem (4)–(7) is a collection of functions satisfying the following conditions:
(Algebraic relation).
(Regularity).
(Initial conditions).
(Variational equations).
for all , with
for ,
,
for .
Then, the existence of the weak solution for the microscopic tridomain problem (4)–(7) is given in the following theorem whose proof is the main issue of the article [5], by using the Faedo–Galerkin method.
(Microscopic Tridomain Model).
Assume that the conditions (6)–(
11
) hold. Then, System (4)–(
7
) possesses a unique weak solution in the sense of Definition
3
for every fixed.
Furthermore, this solution verifies the following energy estimates: there exists constants,,,independent of ε such that:Moreover, if,, then there exists a constantindependent of ε such that:
By summing the two first equations in (19)–(21) and since , we can rewrite the weak formulation as follows:
Unfolding homogenization method
Our derivation of the tridomain model is based on a new approach describing not only the electrical activity but also the effect of the cell membrane and gap junctions in the heart tissue. Our goal in this section is to describe the asymptotic behavior, as , of the solution given by System (4)–(7). We do this by following a three-steps procedure: In Step 5.1, the weak formulation of the microscopic tridomain model (4)–(7) is written by another one, called “unfolded” formulation, based on the unfolding operators stated in the previous part. As Step 5.2, we can pass to the limit as in the unfolded formulation using some a priori estimates and compactness argument to get the corresponding homogenization equation. In Step 5.3, we take a special form of test functions to obtain finally the macroscopic tridomain model.
Unfolded formulation of the microscopic tridomain model
Based on the properties of the unfolding operators, we rewrite the weak formulation (26)–(27) in the “unfolded” form. First, we denote by with the terms of the equation (26) which is rewritten as follows (to respect the order):
Using property (4) of Proposition 1, then the first and second term of (26) is rewritten as follows:
Similarly, we rewrite the third and fourth term using the property (3) of Proposition 1:
Due to the form of , we use the property (2)-(4) of Proposition 1 to obtain for and . Thus, we arrive to:
Similarly, we can rewrite the last two terms of (26) by taking account the form of as follows:
Collecting the previous estimates, we readily obtain from (26) the following “unfolded” formulation:
Similarly, the “unfolded” formulation of (27) is given by:
Convergence of the unfolded formulation
In this part, we pass to the limit in (28)–(29). First, we prove that:
by making use of estimates (22)–(25). So, we prove that when and the proof for the other terms is similar. First, by Cauchy–Schwarz inequality, one has
In addition, we observe that and . Consequently, by Lebesgue dominated convergence theorem, one gets for :
Finally, by using Hölder’s inequality, the result follows by making use of estimate (23) and assumption (6) on .
Let us now elaborate the convergence results of . Using property (5) of Proposition 1 and due to the regularity of test functions, we know that the following strong convergence hold:
and
Next, we want to use the a priori estimates (22)–(25) to verify that the remaining terms of the equations in the unfolded formulation (28)–(29) are weakly convergent. Using estimation (23), we deduce that there exist , for and such that, up to a subsequence (see for instance Theorem 3.12 in [7]), the following convergences hold as ε goes to zero:
and
with the space given by (18). Thus, since a.e. in for and a.e. in , one obtains:
Furthermore, we need to establish the weak convergence of the unfolded sequences that corresponds to , , and for . In order to establish the convergence of , we use estimation (25) to get for
So there exists such that weakly in with . By a classical integration argument, one can show that . Therefore, we deduce that
Thus, we obtain
By the same strategy for the convergence of , there exits such that weakly in . Similarly, we get . Thus, one has
Now, making use of estimate (22) with property (4) of Proposition 1, one has
Then, up to a subsequences,
So, by linearity of and of we have respectively:
Similarly, exploiting assumption (10) on , we obtain the following convergence:
Proceeding exactly as in [4], we prove that the limits and s coincide respectively with for and . Furthermore, since we have assumed that the initial data , for and introduced in (7), are also uniformly bounded in the adequate norm (see assumption (11)). Then, using the weak formulation (28)–(29), we prove similarly that a.e. on Ω, since, by construction, a.e. on for . The same argument holds for the initial condition of for and of .
It remains to obtain the limit of containing the ionic function . By the regularity of , it sufficient to show the weak convergence of to in . Due to the non-linearity of , the weak convergence will not be enough. It is difficult to pass to the limit of this term on the microscopic membrane surface. Therefore, we need the strong convergence of to in for that we obtain by using Kolmogorov–Riesz type compactness criterion that can be found as Corollary 2.5 in [11]:
(Kolmogorov–Riesz type compactness result).
Letbe an open and bounded set. Letfor a Banach space B and. Forand, we define. Then F is relatively compact inif and only if
for every measurable setthe setis relatively compact in B,
for all,and,, there holdswhereand,
for, there holdsfor.
To cope with this, in the following lemma, we derive the convergence of the nonlinear term :
The following convergence holds for:as. Moreover, we have for:as.
We follow the same idea to the proof of Lemma 5.3 in [6]. The proof of the first convergence is based on the Kolmogorov compactness criterion 2. So, we want to verify that the sequence of unfolded membrane potentials satisfies the assumptions of Proposition 2 with for and . It is carried out by proving three conditions:
(i) Let a measurable set. We define the sequence as follows:
It remains to show that the sequence is relatively compact in the space for . Since the embedding is compact, we have to show that the sequence is bounded in with .
We first observe that for
In view of Fubini theorem, Cauchy–Schwarz inequality and estimate (22), it follows that for
Next, we only need to bound the semi-norm and this is done as follows. Since for , we use again Fubini theorem and Jensen inequality together with the trace inequality in Remark 3 to obtain
Hence, integrating over and using the a priori estimates (23), we have showed that the sequence is bounded in for .
By a similar argument and making use of the estimate (25) on , we can also show that
Finally, we deduce that the sequence is bounded in and due to the Aubin-Lions Lemma the sequence is relatively compact in with .
(ii) Due to the decomposition of the domain given in Section 4.1, Ω can always be represented by a union of scaled and translated reference cells. Fix and let , be an index set such that
Note that . For every fixed , we subdivide the cell into subsets with , defined as follows
for a given . It holds .
We use the same notation as in Proposition 2. Now, we compute for the following norm
Proceeding in a similar way to [10,18], we first estimate using the above decomposition of the domain as follows:
which by using the integration formula (4) (for ) of Proposition 1 is equal to
For a given small , we can choose an ε small enough such that . This amounts to saying that in order to estimate , it is sufficient to obtain estimates for given , of
where with an open set.
In order to estimate the norm (30), we test the variational equation the weak formulation (26) with for and , where is a cut-off function with , in Q and zero outside a small neighborhood of Q. Proceeding exactly as Lemma 5.2 in [6], Gronwall’s inequality and the assumptions on the initial data give the following result:
where C is a positive constant. Then, we obtain by using the previous estimate
Hence, we can deduce that as uniformly in ε, as in [12]. Indeed, to prove that
one identifies two cases:
For : take , then, from (31), we get that condition (32) holds for .
For : we remark that since ε tends to , there are only finitely many elements ε in the interval , say with , . Moreover, by the continuity of translations in the mean of -functions, for every n, such that , condition (32) holds. Thus choosing together with the argument for the translation with respect to time, property (32) is proved.
It easy to check that
Hence, we can deduce that as uniformly in ε. Indeed, to prove that
one identifies two cases:
For ε small enough, say , , then .
For : we remark that since ε tends to , there are only finitely many elements ε in the interval , say with , . Moreover, by the continuity of translations in the mean of -functions, for every n, such that , condition (33) holds. Thus choosing together with the argument for the translation with respect to time, property (33) is proved.
This ends the proof of the condition (ii) in Proposition 2.
(iii) The last condition follows from the a priori estimate (24). Indeed, we have for :
The conditions (i)–(iii) imply that the Kolmogorov criterion for holds true in for . This concludes the proof of the first convergence in our Lemma.
It remains to prove the second convergence which will be done as follows. Note that from the structure of and using property (2) in Proposition 1, we have
Due to the strong convergence of in , we can extract a subsequence, such that a.e. in with . Since is continuous, we have
Further, we use estimate (24) with property (4) of Proposition 1 to obtain for
Hence, using a classical result (see Lemma 1.3 in [16]):
Moreover, we obtain, using Vitali’s Theorem, the strong convergence of to in and . This finishes the proof of our Lemma. □
Finally, we pass to the limit when in the unfolded formulation (28) to obtain the following limiting problem:
Similarly, we can prove also that the limit of (29) for as ε tends to zero, is given by:
Since the linear term H is not varying at the micro scale and since does not depend on y, it can be proven, using Assumption (8b), that the solution of
is unique for all for hence it is independent of the variable y.
Derivation of the macroscopic tridomain model
The convergence results of the previous part allow us to pass to the limit in the microscopic equations (19)–(21) and to obtain the homogenized model formulated in Theorem 1.
To this end, we choose a special form of test functions to capture the microscopic informations at each structural level. Then, we consider that the test functions have the following form:
with functions and for defined by:
where , , and are in , in and in for . Then, we have:
Due to the regularity of test functions and using property (5) of Proposition 1, there holds for , when :
Since for and , then it holds also:
where for and .
Collecting all the convergence results of obtained in Section 4.2, we deduce the following limiting problem:
Similarly, we can prove also that the limit of the coupled dynamic equation for as ε tends to zero, which is given by:
Now, we will find first the expression of in terms of the homogenized solution for . Then, we derive the cell problem from the homogenized equation (37). Finally, we obtain the weak formulation of the corresponding macroscopic equation.
We first take , and for are equal to zero, to get:
Since is independent on the microscopic variable y then the formulation (39) corresponds to the following microscopic problem:
Hence, by the y-periodcity of and the compatibility condition, it is not difficult to establish the existence of a unique periodic solution up to an additive constant of the problem (40) (see [3] for more details).
Thus, the linearity of terms in the right-hand side of (40) suggests to look for under the following form in terms of :
where is a constant with respect to y and each element of satisfies the following ε-cell problem:
for . Moreover, the compatibility condition is imposed to guarantee the existence and uniqueness of solution to problem (42) with is given by (18).
Finally, inserting the form (41) of into (37) and setting , to zero, one obtains the weak formulation of the homogenized equation for the intracellular problem:
with , , and the coefficients of the homogenized conductivity matrices defined by:
Similarly, we can decouple the cell problem in the extracellular domain and define the homogenized matrix . This completes the proof of Theorem 1 using periodic unfolding method.
Since the conductivity matrices for are symmetric then the homogenized conductivity matrices defined by (14a)–(14b) are also symmetric for .
We can rewrite the homogenized conductivity matrices as follows
Indeed, we recall that is the solution of (42). Choosing as test function in (42), one has
Hence, one obtains
On the other hand, since
formula (44) can be written as follows:
Summing (46) from (47) gives (45). Similarly, we can rewrite the other matrix in terms of the corresponding corrector function .
Since the conductivity matrices for satisfy the elliptic conditions defined by (6), then the homogenized conductivity matrices , verify the following elliptic conditions: there exits such that
Indeed, let and . To prove (48a), then from (45) it follows that
Setting and using the ellipticity of defined by (6), we get
Let us show that this inequality implies that
If this were not true. In view of (49), one would have some such that
This means that
Thus, one has
and this impossible since the right-hand side function is y-periodic by definition and . To end the proof of ellipticity, we know that the function is continuous on the unit sphere which is a compact set of . Hence, this function achieves its minimum on and, due to the previous result, this minimum is positive. So, there exists such that
Consequently,
since the vector belongs to . This ends the proof of inequality (48a) and by the same way we obtain the second inequality.
References
1.
M.Amar, D.Andreucci, P.Bisegna and R.Gianni, On a hierarchy of models for electrical conduction in biological tissues, Mathematical Methods in the Applied Sciences29(7) (2006), 767–787. doi:10.1002/mma.709.
2.
M.Amar, D.Andreucci, P.Bisegna, R.Gianniet al., A hierarchy of models for the electrical conduction in biological tissues via two-scale convergence: The nonlinear case, Differential and Integral Equations26(9/10) (2013), 885–912.
3.
F.Bader, M.Bendahmane, M.Saad and R.Talhouk, Derivation of a new macroscopic bidomain model including three scales for the electrical activity of cardiac tissue, Journal of Engineering Mathematics131(1) (2021), 1–30. doi:10.1007/s10665-021-10174-8.
4.
F.Bader, M.Bendahmane, M.Saad and R.Talhouk, Three scale unfolding homogenization method applied to cardiac bidomain model, Acta Applicandae Mathematicae176(1) (2021), 1–37. doi:10.1007/s10440-021-00459-6.
5.
F.Bader, M.Bendahmane, M.Saad and R.Talhouk, Microscopic tridomain model of electrical activity in the heart with dynamical gap junctions. Part 1–modeling and well-posedness, Acta Applicandae Mathematicae179(1) (2022), 1–35. doi:10.1007/s10440-022-00498-7.
6.
M.Bendahmane, F.Mroue, M.Saad and R.Talhouk, Unfolding homogenization method applied to physiological and phenomenological bidomain models in electrocardiology, Nonlinear Analysis: Real World Applications50 (2019), 413–447. doi:10.1016/j.nonrwa.2019.05.006.
7.
D.Cioranescu, A.Damlamian, P.Donato, G.Griso and R.Zaki, The periodic unfolding method in domains with holes, SIAM Journal on Mathematical Analysis44(2) (2012), 718–760. doi:10.1137/100817942.
8.
D.Cioranescu, A.Damlamian and G.Griso, The Periodic Unfolding Method, Series in Contemporary Mathematics Vol. 3, (2018). doi:10.1007/978-981-13-3032-2.
9.
P.Colli-Franzone, L.F.Pavarino and S.Scacchi, Mathematical and numerical methods for reaction-diffusion models in electrocardiology, in: Modeling of Physiological Flows, Springer, 2012, pp. 107–141. doi:10.1007/978-88-470-1935-5_5.
10.
S.Dobberschütz, Homogenization of a diffusion-reaction system with surface exchange and evolving hypersurface, Mathematical Methods in the Applied Sciences38(3) (2015), 559–579. doi:10.1002/mma.3089.
11.
M.Gahn and M.Neuss-Radu, A characterization of relatively compact sets in lp (ω, b), Stud. Univ. Babes-Bolyai Math61(3) (2016), 279–290.
12.
M.Gahn, M.Neuss-Radu and P.Knabner, Homogenization of reaction–diffusion processes in a two-component porous medium with nonlinear flux conditions at the interface, SIAM Journal on Applied Mathematics76(5) (2016), 1819–1843. doi:10.1137/15M1018484.
13.
C.S.Henriquez and W.Ying, The bidomain model of cardiac tissue: From microscale to macroscale, in: Cardiac Bioelectric Therapy, Springer, 2009, pp. 401–421. doi:10.1007/978-0-387-79403-7_16.
14.
H.Hogues, L.J.Leon and F.A.Roberge, A model study of electric field interactions between cardiac myocytes, IEEE transactions on biomedical engineering39(12) (1992), 1232–1243. doi:10.1109/10.184699.
15.
K.H.Jæger, A.G.Edwards, A.McCulloch and A.Tveito, Properties of cardiac conduction in a cell-based computational model, PLoS computational biology15(5) (2019), e1007042. doi:10.1371/journal.pcbi.1007042.
16.
J.-L.Lions, Quelques méthodes de résolution des problemes aux limites non linéaires, 1969.
17.
J.C.Neu and W.Krassowska, Homogenization of syncytial tissues, Critical reviews in biomedical engineering21(2) (1993), 137–199.
18.
M.Neuss-Radu and W.Jäger, Effective transmission conditions for reaction-diffusion processes in domains separated by an interface, SIAM Journal on Mathematical Analysis39(3) (2007), 687–720. doi:10.1137/060665452.
19.
M.Pennacchio, G.Savaré and P.Colli Franzone, Multiscale modeling for the bioelectric activity of the heart, SIAM Journal on Mathematical Analysis37(4) (2005), 1333–1370. doi:10.1137/040615249.
20.
A.Tveito, K.H.Jæger, M.Kuchta, K.-A.Mardal and M.E.Rognes, A cell-based framework for numerical modeling of electrical conduction in cardiac tissue, Frontiers in Physics5 (2017), 48. doi:10.3389/fphy.2017.00048.