
Abstract
Background:
The processes of cell spreading and crawling are frequently associated with mysterious waves and ruffling cycles of the leading edge.
Objective:
To develop a physical model that can account for these phenomena based on a few simple and plausible rules governing adhesion, contractility, polymerization of cytoskeleton, and membrane tension.
Methods:
Extension of a continuum mechanical model of phagocytosis [J Cell Sci. (2006);119(Pt 9):1903–13] adding a simple coupling between membrane curvature and cytoskeletal polymerization.
Results:
We show that our generalized model has just the right nonlinearity needed for triggering of stochastic/chaotic cycles of ruffling similar to those that are observed in real cells.
Conclusions:
The cycles are caused by a branching instability at the leading edge that leads to bifurcations of protrusion into forward moving lamellipodium and upward and rearward folding ruffles. The amplitude of the instability is modulated by the surface tension, with higher tension stabilizing against ruffling (but inhibiting protrusion) and lower tension promoting ruffling and protrusion.
Introduction
For anyone who has observed the motion of ameboid cells in a dish under a microscope, the overall sense is that of a curious mixture of chaotic and purposeful movement. Once a round cell has settled and spread onto a flat surface, the thin lamella at the leading edge undergoes intermittent protrusion along the substratum. Also often observed is ruffling during which protrusion appears to take place upward and perpendicular to the substratum and then fold back upon itself [1,2]. Over time, protrusion dominates, but as mentioned in early work [3] “the impression given is of two opposed and slightly imbalanced forces”. Other centripetally traveling waves are seen to move from the periphery all the way to the cell body [4,5]. While there is general agreement that cytoskeletal polymerization is the main driver of activity, identification of the molecular players and their roles remains the focus of much experimental work [6,7].
The mechanical theory of flattening, lamella progression, and ruffling is less advanced although a number of attempts have been made to provide frameworks that explain the observed behaviors in a physically self-consistent representation of mechanical forces and rheological properties. For instance, we have in the past generated a model of phagocytosis [8] which essentially reproduced lamellar spreading over a surface, but that model did not exhibit any cyclic behavior of the ruffling type.
In this paper, we develop an integrative toy model of the mechanics leading to the flattening of a cell on a substrate, followed by the generation of protrusion and ruffling. Our aim is not literal, but rather to illustrate in a schematic way, the mechanisms by which complex motile behavior can unfold from simple dynamic rules
We begin with a discussion of the model from an intuitive perspective, then present the result of a representative calculation. We end with a discussion of the key take-aways from our approach. To appreciate the essence of this work, it is essential to watch the movie of our simulation viewable at
Methods
General concepts
Our overall goal is to construct a model that will allow us to follow a spherical cell settling on a substratum and turning into a flat structure while displaying the phenomena of lamellar protrusion and ruffling that are commonly observed. For this model, our main requirements are a high degree of simplicity and generality via the incorporation of a number of biologically plausible mechanical processes that is as small as possible. Starting from a spherical cell in tangential contact with the substratum as might be expected in conditions where a cell in suspension first gains contact with the substratum, the key events that mechanisms of motility have to enable are the following:
A process by which the cell expands the contact area with the substratum from the initial point of contact.
A process by which the cell grows a thin, flat, lamellar protrusion more or less parallel to the substratum, but with occasional departures in the vertical direction (ruffling).
A process by which the cell-substratum contact area becomes adherent.
To effect the above we invoke the following mechanisms (depicted schematically in Fig. 1).
When contact is made by the ventral membrane of the cell with the surface of the substratum, there is a delay (we use
Once contact with the substratum has been ongoing for a sufficient long time (we use
As membrane becomes permanently adherent to the substratum, downward contractility comes into effect flattening cytoskeleton and the cell toward the substratum. This contractility remains in effect for a limited time (in our calculation 100 s), so that the contact region furthest away from the leading edge is typically passively stick (see Fig. 1).
Wherever the inward convex curvature of the membrane exceeds a threshold value (here we use
Mechanisms 1 and 2 are well established, in a sense it is a primitive implementation of what has been labeled “graded adhesion” driven by the gradual maturation of focal adhesions under tension [9–11]. Mechanism 3 is not canonical but we have found it necessary to enable the early flattening that drives the contact area to become larger. In prior work, we have also found it necessary to invoke such contractile force in the process of phagocytosis [8,12]. Mechanism 4 was initially mooted by Atligan et al. [13] and subsequently explored by others (e.g. [14]). Relying on the propensity of certain membrane complexes to localize in regions of high curvature, it provides a simple and natural mechanism to provide directional control to the growth of protrusions. As has been surmised and will be confirmed here it also provides an explanation for the chaotic behavior observed at the leading edge as well as the dorsal waves that can propagate centripetally toward the cell body.
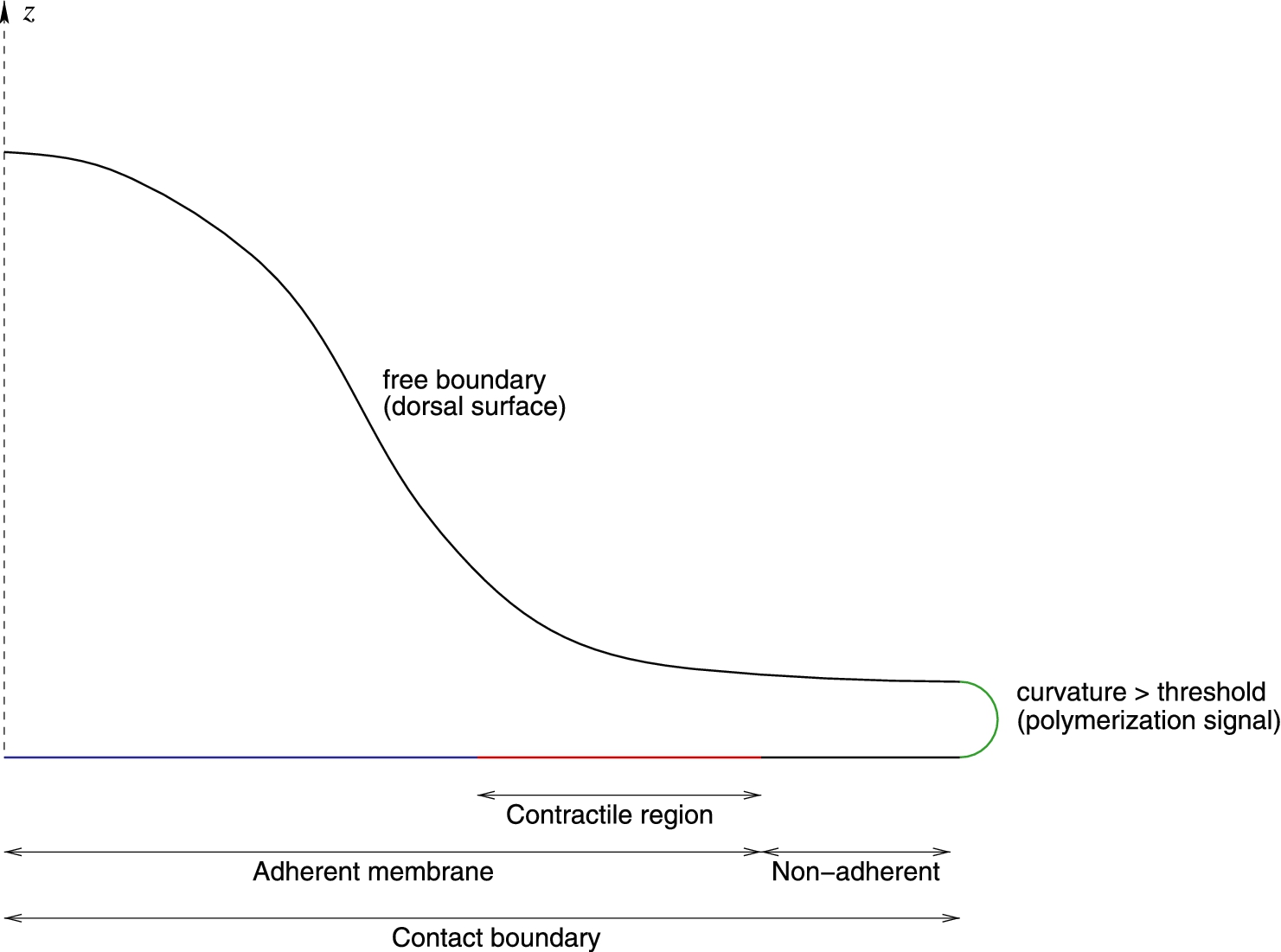
Idealized schematic of boundaries used in the model. The z axis is an axis of cylindrical symmetry for the calculations. The dorsal surface (black) is free. Where the membrane curvature is high (
These mechanism likely represent a set of nearly minimal complexity to achieve the desired behaviors but we acknowledge that there is no limit to the number of features that could be added and likely exist in nature for various cell-types and ambient conditions. We also note that we do not consider here a process for detachment of regions adherent to the substratum as would be required to study whole cell translocation.
The starting point of our approach is the reactive interpenetrating flow formalism developed several decades ago [15], and applied in many mechanical/motile cellular phenomena [8,16,17]. Briefly, it relies on a continuum mechanical view of the cell that corresponds to coarse graining at a scale that is large compared to molecules, but still small compared to the whole cell. The cell interior is conceptualized as a mixture of two phases, the cytoskeleton (the network) and the cytosol (the solvent), enclosed by an envelope, the cell membrane. The cytoskeleton plays the crucial role in controlling the dynamics of cell deformation, whereas the cytosol is a “filler” material whose flow is driven by pressure gradients. The two phases can convert into each other reflecting, for instance, the polymerization/depolymerization cycle of G- and F-actin. A deformable boundary endowed with surface tension represents the cortical membrane. Mass and momentum conservation equations determine the self-consistent time-evolution of the model. The mathematical and numerical details of the implementation of this approach have been presented several times (references above) and we will not repeat those here except for components that are specific to this particular application.
Our template cell is based on a computational model of the human neutrophil that we have previously outlined [8]. Like in previous work, we employ a two-dimensional finite element model of the cell with cylindrical symmetry around the vertical axis. The diameter of the cell in its spherical state is 8.5 µm and the volume remains constant during the calculations.
In the absence of stimulus, equilibrium cytoskeletal density (network volume fraction) is
The network phase is treated as a fluid with a large viscosity which increases linearly with the network volume fraction
For cortical tension we use a simple phenomenological law
Results
The dynamical manifestations of the system that we study here are essential for the appreciation of the underlying mechanisms, and for this reason we strongly recommend that the reader review the movie (play at

At
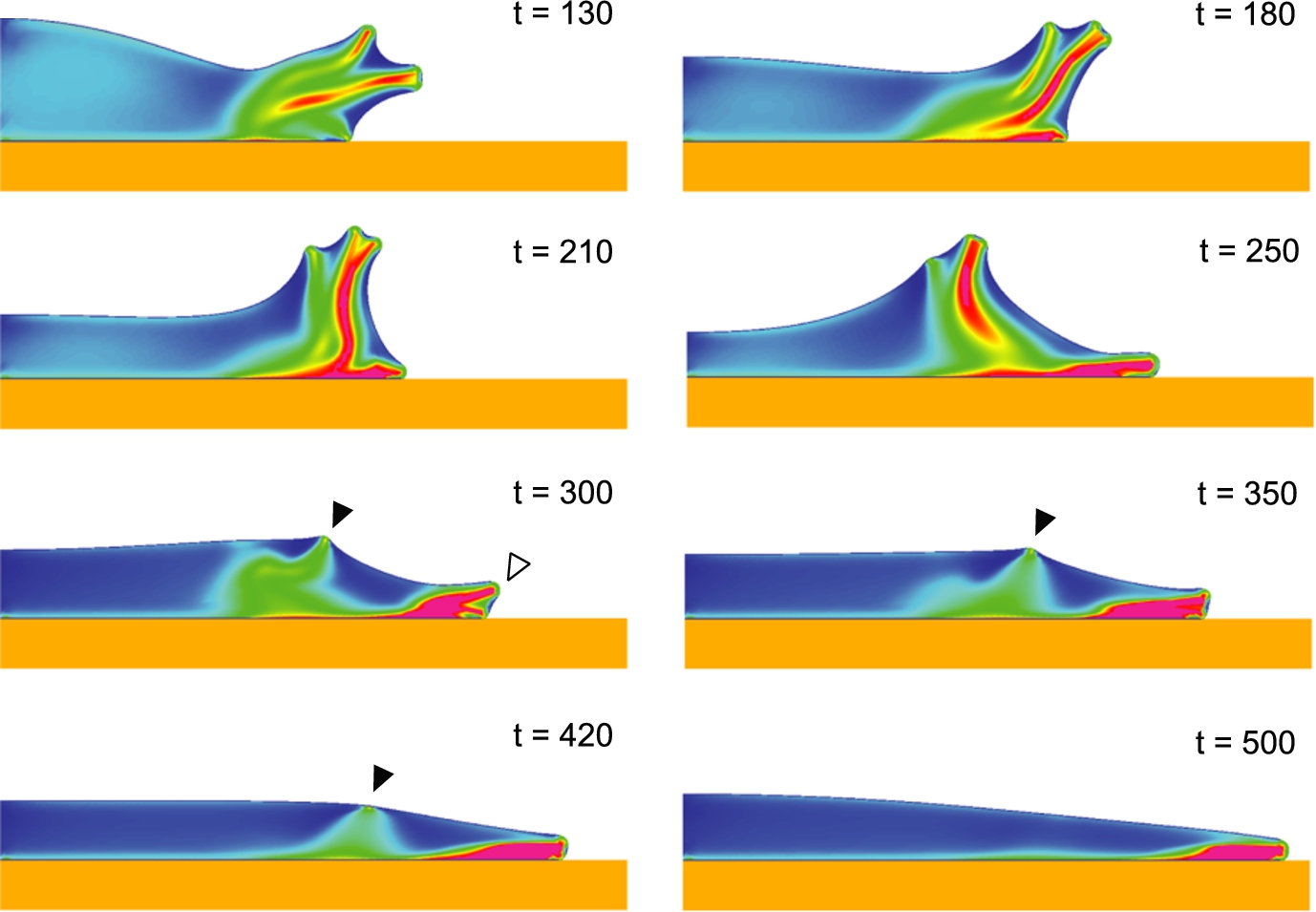
Figure 2 shows the evolution of the model during the initial phase “flattening” phase. We begin with the spherical cell barely tangent with the substratum (
Eventually however, two factors lead to an arrest in the outward progression of the contact line along the substratum. The first factor is that with the departure from sphericity and related increase in surface area, the cortical tension increases and impedes further flattening. The second factor is that since we have a decay of the contractility for areas that have been adherent to the substratum more than 100 s, the flattening driving force eventually falls too (because no new adherent area is created). As a result, after about 500 s of simulation, the contact line is no longer moving out, and as the cell rebounds to a more spherical shape, the contact angle becomes significant (as long as the contact line progresses, the contact angle is zero) and therefore the local curvature of the membrane becomes significant.

Continuing from the flattening phase shown in Fig. 2, the high curvature at the contact line triggers polymerization of network (see open arrowhead in
Continuing from the protrusion phase shown in Fig. 3, a ruffle appears as the lamellipodium rises up and folds back into the rear part of the lamellipodium. As the surface tension becomes higher, the ruffle become a dimple (see filled arrowhead,
The next phase of the evolution of the model corresponds to the generation of a lamella protruding along the substratum (Fig. 3). Because of the contact angle increase, the local membrane curvature is high and this triggers local polymerization of network at the contact line driving a lamellar protrusion outward (see
As a region of high curvature grows into a protrusion, there is a tendency for the foremost portion to become blunter (less curvature) and for the immediate sides of the tip of the protrusion to increase in curvature. The end result is that two foci of polymerization are spawned where there was previously one (see
Once multiple polymerization foci are active, subsequent evolution tends to be stochastic. An individual protrusion can steer away perpendicular to the substratum. However, whenever the contact angle increases (as will be the case when the leading protrusion lifts up) a new focus of polymerization will appear in the contact area and drive a separate protrusion along the substratum. As a protrusion becomes “dorsal” above a lower protrusion at the contact line, it is entrained towards the rear by the retrograde flow of network. From then on, behavior is reminiscent of the behavior of listeria or other cytoplasmic inclusions capable of inducing localized cytoskeletal polymerization, with a jet of polymerization driving a protrusion that is sliding and weaving tangentially along the membrane (e.g. [21,22]). This persists until the increase of surface area drives the cortical tension to a point that the polymerization force is no longer able to maintain a dorsal protrusion with the requisite amount of curvature which leads to an abrupt disappearance of the corresponding ruffle. As the end of the simulation, there is just one region of polymerization at the leading edge of the lamella which is stalled by the surface tension.
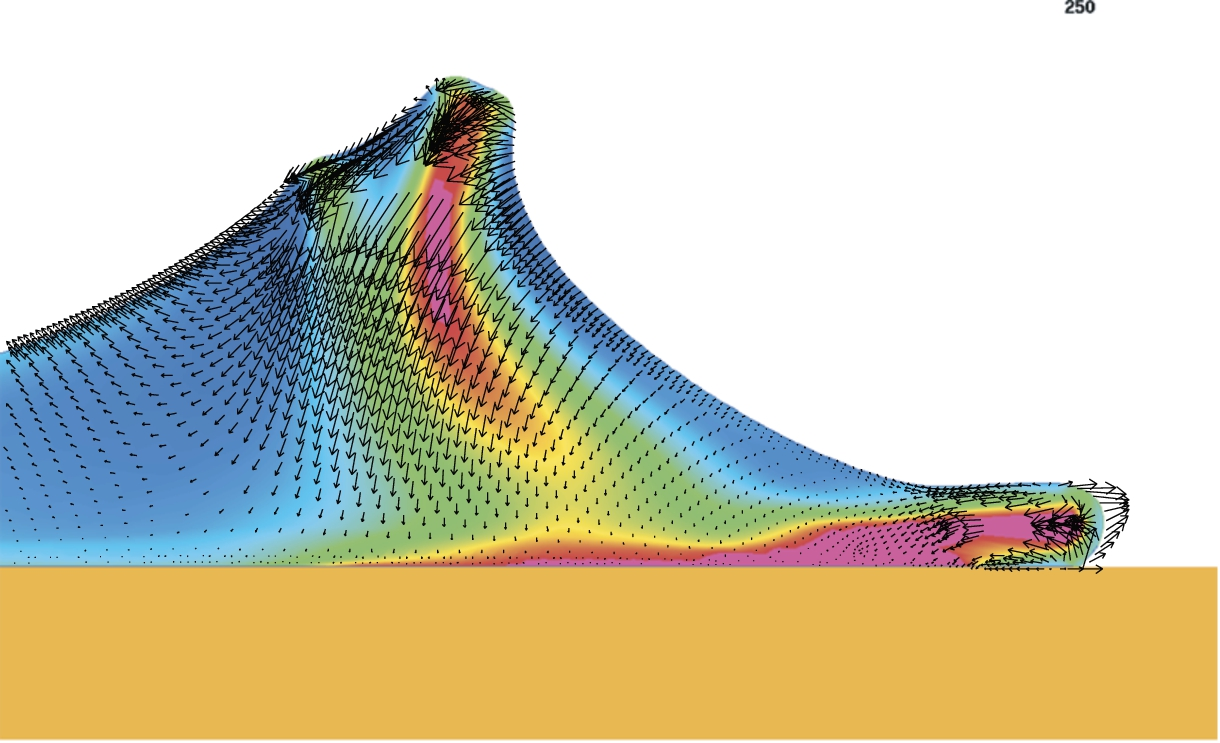
Detail of
Through a very simple, some might even say simplistic, computational model, we have shown here how a few basic physical processes can lead to dynamical behaviors that replicates many in vivo observations of actual cells. The main conclusions of our work are as follow:
At appropriate timescales that match typical strain rates for self-driven cellular motions, a fluid picture of the cell is sufficient for interpreting many aspects of cell motility. While there is no doubt that for rapid, externally imposed deformations, there are memory effects that can best be explained by elasticity, the dynamic turnover of the cytoskeleton precludes persistence of this memory much beyond the turnover timescale.
Membrane curvature appears to be a natural driver of internal polymerization that can explain a number of phenomena observed in living cells. First presented in detail by Atilgan et al. [13], the simple idea that polymerization will occur in regions of high inwardly concave membrane curvature is capable of explaining a number of puzzles in an elegantly simple solution.
Membrane tension is a key component of mechanical cohesion: it enables propagation of mechanical signals across the cell in a way that the cytoplasm cannot provide.
Timing of anchoring of the leading edge plays a critical role in the dynamics of the leading edge. Rapid adhesion promotes persistence of the lamella whereas delayed adhesion enables the more chaotic behavior of the lamellipodium [23,24].
These are not new concepts, but they are here brought together in a novel integrated model of cellular spreading that shows representative emergent properties of the combination of those mechanical processes.
As exemplified by some of the key references in this paper which go back decades, explicit data on the vertical dynamics of the cell (i.e. transverse with respect to the substratum plane) are extremely sparse. Experimental studies where cells are observed top down on a flat substratum are the norm, and typically thickness data are lacking (although we note that the simple mapping with a cytosolic fluorophore volume marker could provide additional insights on lamellar dynamics). Studies based on side views of the lamella and lamellipodiam have been exceedingly rare.
In parallel, computational models of cell mechanics and motility often neglect the vertical dimension for practical reasons and focus on a two dimensional view of the contour of the cell projected on the substratum via a variety of thin film approximations [25–28]. It is to complement those models that we have here chosen an approach that makes abstraction of the direction parallel to the leading edge to focus on the height and protrusion directions. Of course, much more specific and detailed investigations of this type are possible, but they will only be meaningful when more experimental data are available.
Footnotes
Acknowledgement
This work was supported by NIH grants R01 A1072391 and R01 GM72002.