
Abstract
Background:
Leukocytes and platelets typically fulfil their functions through adhesion to the walls of vessels with different size, haematocrit and shear rate.
Objective:
We aimed to investigate differential effects of these variables on leukocyte and platelet adhesion.
Methods:
Blood with varying haematocrit was perfused at a range of wall shear rates through capillaries of depth 100 or 300 µm coated with P-selectin or collagen.
Results:
Adhesion of leukocytes was much more efficient in the smaller capillaries, but was equal on the upper and lower surfaces and showed nearly identical shear rate dependence for either size of vessel. Platelets also adhered more efficiently in the smaller vessels (although the effect of size was not so great), and equally on upper and lower surfaces, but their adhesion was much less sensitive to increasing shear rate. In previous studies using vertically-orientated capillaries, leukocyte adhesion increased with increasing haematocrit (Am. J. Physiol.
Conclusions:
Platelets adhere efficiently over a wider range of sizes and shear rates, and at high haematocrit. Leukocytes adhere better in smaller vessels and at low haematocrit in horizontal vessels. The different behaviours may represent ‘rheological adaptation’ to functions in inflammation vs. haemostasis.
Introduction
The functions of leukocytes in immunity and of platelets in haemostasis require them to adhere to the walls of inflamed or damaged blood vessels. Both cells types show initial unstable, rolling adhesion upon capture from flow which is stabilised by signals which induce conformational change in integrin receptors allowing them to form firm bonds [1,2]. However, capture has quite different rheological characteristics for the two. It is evident that platelets can adhere at much higher shear rates than leukocytes [3–6]. In addition, red cell aggregation promotes attachment of leukocytes but inhibits adhesion of platelets [7]. Red cell aggregation has been well-described to promote margination of leukocytes in the blood stream [8], but this does not seem to be the case for platelets [7]. We previously ascribed the above differences in adhesion to differences in cell size which affect near-wall velocity before capture and shear force experienced upon attachment, and also determine margination in the blood stream and effects of the narrow red cell-depleted layer at the periphery of vessels [7]. These observations fit with physiological observations that leukocytes predominantly adhere in post-capillary venules (with low shear rate and more extensive red cell aggregation, especially in inflammation) while platelets act more efficiently in arteries where shear rates are high and aggregation minimised.
However, there are other physical factors that are relevant to the differences in adhesive behaviour. Firstly, leukocyte adhesion occurs generally in vessels with smaller diameter, which may influence the proportion of cells delivered to the wall, as well as reducing the haematocrit due to the Fahraeus effect (i.e., tube haematocrit decreasing relative to systemic haematocrit as vessel diameter decreases below about 300 µm) [9]. Studies of purposeful variation in haematocrit for a given vessel size have previously shown that red cells promote contact and adhesion of leukocytes up to a haematocrit of about 40%, after which there is little effect [10]. This was in vertically orientated vessels to avoid effects of blood cell sedimentation. Adhesion of platelets increased steadily for haematocrits up to 60–70% in horizontal flow chambers at wall shear rates ∼1000 s−1 [11,12]. In fact, the effects of vessel orientation (horizontal vs. vertical) and of cell sedimentation on adhesion have been largely ignored, although sedimentation is more rapid for larger cells. In the absence of red cells, leukocytes sediment efficiently in narrow horizontal tubes [13], so that adhesion from flow occurs on the lower but not upper surface [14]. Platelets sediment more slowly. Adhesion during perfusion over everted vessel segments was inefficient in the absence of red blood cells [12], and sparse adhesion to upper or lower surfaces might be expected for perfusion of platelets without red cells through a horizontal flow chamber. However, the patterns of adhesion of leukocytes or platelets on the upper or lower surfaces for perfused blood are unknown, as are the effects of the interaction between haematocrit and sedimentation.
To obtain a better understanding of rheological factors differentially affecting adhesion of leukocytes or platelets in different vessels, we carried out studies in which blood was perfused through glass capillaries coated with appropriate adhesive substrates (P-selectin or fibrillar collagen respectively). We also made observations in non-adhesive capillaries to assess numbers marginated near the wall without attachment. Adhesion was compared on upper and lower surfaces of different-sized capillaries (depths of 100 vs. 300 µm) and also at a range of haematocrits. We found that different dependence of leukocyte and platelet adhesion on vessel size, orientation and haematocrit appeared to be adapted to their functions in venules and arteries respectively.
Methods
Blood collection and manipulation of haematocrit
Blood was drawn from healthy adult volunteers with informed consent and with ethical approval of the Life and Health Sciences Ethical Review Committee of the University of Birmingham. The blood was mixed 9:1 with anticoagulant citrate phosphate dextrose adenine (CPDA; pH 7.4; Sigma-Aldrich Company Ltd). Leukocytes were counted using a Coulter Counter (Counter Multisizer II, Coulter Electronics Ltd, Luton, UK). Platelets were counted using a Pentra 60 haematology cell counter (Horiba ABX, Northampton, UK). The haematocrit was measured by the micro-haematocrit method after blood was centrifuged at 15,000g for 5 minutes. To alter haematocrit for studies of platelet adhesion, several tubes containing 10 ml of blood of known haematocrit were centrifuged at 550g for 10 minutes. An amount of the supernatant platelet-rich-plasma (PRP) was removed from one tube to leave blood of 50% haematocrit. PRP harvested from the tubes was then added to aliquots of this blood to obtain haematocrits of 10, 20, 30, 40 and 50. We counted platelets in the PRP and found the platelet concentration to be approximately the same as that in the original blood. Thus, platelet count did not vary systematically when we varied haematocrit. To alter haematocrit in the leukocyte adhesion studies, autologous plasma was added to the blood of known haematocrit and leukocyte count, to obtain haematocrits of 10, 20, 30, and 40%. This procedure predictably reduced the leukocyte count in these samples. In all experiments, leukocyte adhesion was normalised for the leukocyte count in the blood (see below), and thus the effect of dilution accounted for. All haematocrits were confirmed by micro-haematocrit measurement.
Perfusion of blood and evaluation of wall shear rate, wall shear stress and viscosity
The known, measured and derived experimental parameters are summarised in Table 1.
Known, measured and derived experimental parameters
Known, measured and derived experimental parameters
Microslides (glass capillaries with length (L) 50 mm, and rectangular cross-section height (h) × width (w) of
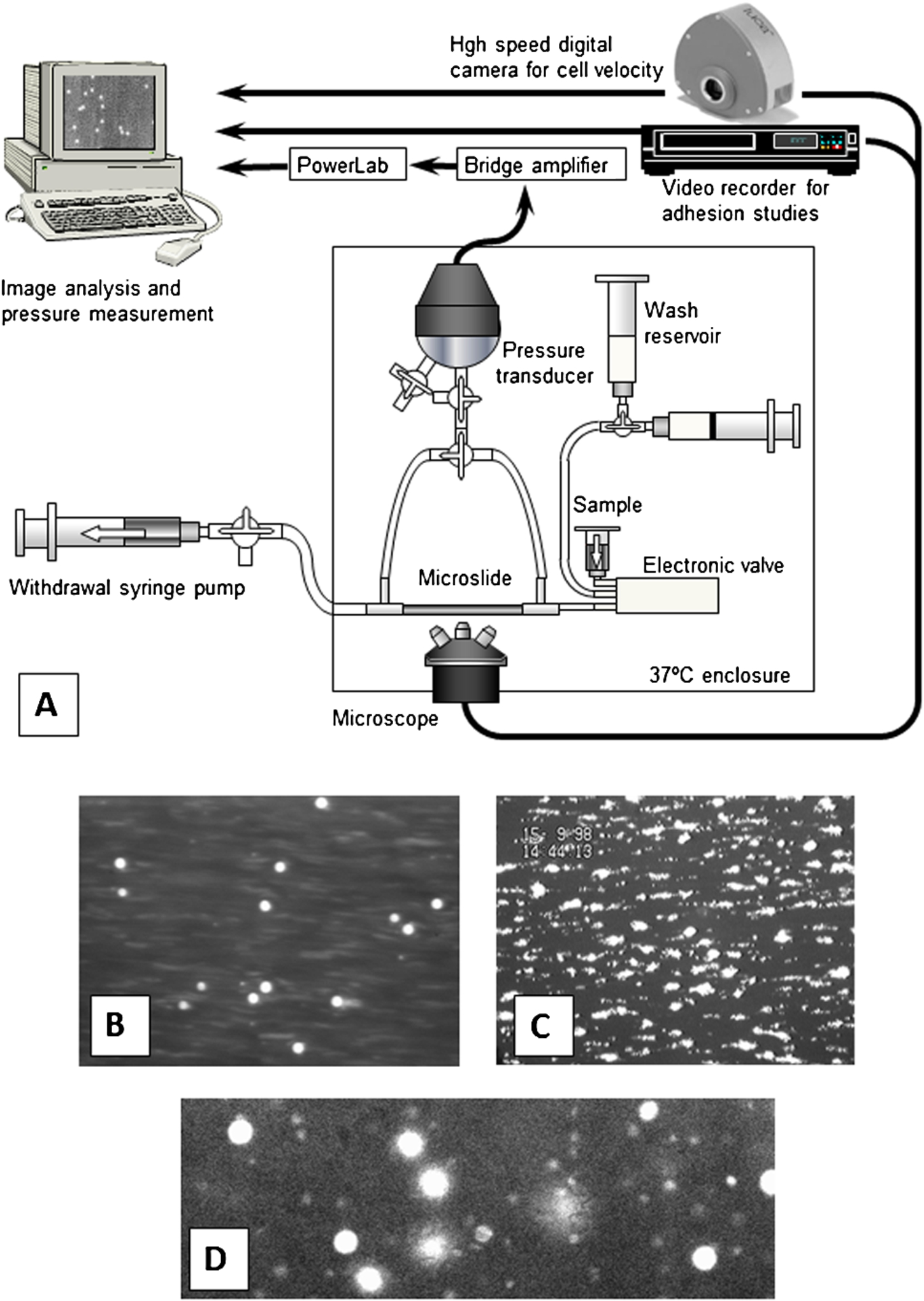
Flow assay and outcome for perfusion of blood over differently-coated surfaces. (A) Schematic of apparatus for analysis of adhesion or margination of leukocytes or platelets in flowing whole blood, and for measurement of pressure drop and calculation of the apparent viscosity of the blood. (B) Digitised video image of rhodamine-labelled leukocytes adherent to P-selectin, visible as bright circles which were moving (rolling) slowly over the surface. (C) Rhodamine labelled platelets adherent to collagen, forming aggregates after initial adhesion. (D) High speed camera frame of rhodamine-stained leukocytes and platelets flowing adjacent to the wall of a microslide coated with albumin alone. (Colors are visible in the online version of the article;
Microslides were attached to a flow system which allowed blood to be perfused at chosen volumetric flow rates at 37°C as described [7,10]. To minimise sedimentation in the inflow system, the blood sample reservoir was directly attached to an electronic valve with a small internal dead volume whose outflow was directly attached to the microslide by a short segment of silicon tubing; final dead-space was approximately 20 µl (see Fig. 1(A)). A pressure transducer was connected to either end of the microslide and absolute pressure was recorded at each end in turn and the pressure drop calculated for each flow rate or condition used. From the pressure drop measured at an imposed volumetric flow rate (Q), and the microslide dimensions, apparent viscosity of the blood (
The fluorescent dye rhodamine 6G (R6G) was added to blood at 5 µg/ml to label leukocytes and platelets [14] and their adhesion to P-selectin or collagen was analysed as recently described in detail [7]. Microslides filled with blood are effectively opaque to transmitted light. Thus microslides were mounted on either an Olympus BX51 upright fluorescence microscope, or an Olympus IX70 invert fluorescence microscope, for observation of the upper or lower inner surfaces respectively. Video recordings were made during perfusion of blood for 4 min, and subsequently sequences were digitised and analysed off-line using Image Pro software (Media Cybernetic, Inc, Bethesda, MD, USA). For P-selectin, the number of adherent leukocytes (rolling or stationary) was counted (see Fig. 1(B)) and converted to cells/mm2/106 perfused using the known image area, concentration in the blood and volume perfused. When desired, this was multiplied by the sum of the upper and lower surface areas of the microslides, to calculate the percentage of all the leukocytes perfused which adhered, thus providing a measure of the efficiency of adhesion. The velocity of the rolling cells was calculated by measuring the distance moved in 10 s. For collagen, individual platelets adhered initially and small aggregates built up over time (see Fig. 1(C)). The area covered by platelets was analysed as a percentage of the total area of the field, after image thresholding and area analysis using Image Pro. The area was normalised per ml of blood perfused to allow for different volumes perfused at different shear rates. When comparing different sized microslides, we also calculated the absolute area (mm2) covered per ml of blood, to again obtain a measure of comparative efficiency independent of size.
For microslides coated with albumin, the numbers of free flowing cells near the wall and their velocities were measured as recently described [7]. Digital images were captured using a Luca EM-CCD camera (ANDOR Luca DL-658M-TIL, Belfast, UK) with short exposure time (0.0005 s) and rapid frame rates (95/s). An oil immersion objective with 25× magnification and a numerical aperture of 0.8 (LD LCI Plan-Apochromat, Carl Zeiss Ltd, Herts, UK) gave a thin depth of focus (∼2 µm), but with low enough magnification to include enough cells in a frame for reliable analysis. The short exposure time resulted in images of free-flowing cells that were not blurred, and rapid frame rate allowed measurement of velocity (see Fig. 1(D)).
Statistical analysis
Data are shown as mean ± SEM. Effects of varying conditions were evaluated using analysis of variance (ANOVA), followed by Dunnett’s test for comparison of an individual condition to control, or Bonferroni test for comparison between multiple conditions. All tests were performed using Minitab (Minitab Inc, State College, PA).
Results
Effects of vessel size and wall shear rate on leukocyte and platelet adhesion
In studies of vessel size and shear rate, whole blood was perfused at the native haematocrit which averaged 42.5%. We first compared adhesion of leukocytes to P-selectin in microslides with depth 100 or 300 µm over a range of shear rates. Adhesion to the upper or lower surface was analysed separately. Numerous fluorescent leukocytes were visible attaching to the surfaces, where they rolled slowly with velocity ∼100 times slower than unattached cells visible as blurred streaks (see e.g., Fig. 1(B)). Figure 2(A), (B) shows that for either size of microslide, adhesion fell off sharply as wall shear rate was increased from 70 to 280 s−1, and was similar on the upper or lower surfaces. Surface densities of adhesion per million leukocytes perfused were much higher in the smaller microslides but this parameter did not allow for the larger surface area presented by the deeper microslide. To take this into account, an efficiency of attachment was calculated, i.e., percentage of all perfused cells becoming adherent including upper and lower surfaces. This efficiency remained about seven-fold higher in the smaller capillaries over the range of shear rates tested (Fig. 2(C)). Examining the velocity of the rolling adherent cells, this did not vary significantly with shear rate, was similar on the upper and lower surfaces, but was slower in the larger capillaries (Fig. 2(D), (E)). To assess margination, the number density of leukocytes flowing close to the wall was first quantified (corrected for the blood count) at a wall shear rate of 140 s−1. It was almost exactly equal for the 300 vs. 100 µm microslides (

Effects of varying wall shear rate on the adhesion of leukocytes to P-selectin when blood was perfused at native haematocrit through microslides of different size; comparison of adhesion to upper and lower surfaces. (A), (B) Number of perfused leukocytes adherent on the upper or lower surfaces of 100 µm or 300 µm microslides. (C) Percentage of perfused leukocytes adhering in 100 µm vs. 300 µm microslides (sum or upper and lower surfaces). (D), (E) Rolling velocity of leukocytes adherent on the upper or lower surfaces of 100 µm or 300 µm microslides. Data are mean ± SEM from 3 experiments. ANOVA showed significant effects of shear rate on adhesion (
For platelets, adhesion was studied over wall shear rates from 300–1500 s−1 for the 100 µm microslides but only 70–560 s−1 for the 300 µm microslides because of the volume of blood required. Deposition of platelets increased with increasing shear rate, but when corrected for volume of blood delivered, area coverage per ml blood perfused showed a downward trend at the higher shear rates used in the 100 µm slides (Fig. 3(A)). At the low shear rates tested in the larger capillaries, adhesion tended to be higher as the wall shear rates approached arterial levels (Fig. 3(B)). Again, adhesion was similar on the upper and lower surfaces over the different ranges of shear rates. The percentage of the surface covered per ml of blood perfused was much greater for the smaller microslides. However, this does not account for the much larger surface to be covered in the larger slides, and so we also calculated the absolute area (mm2) covered with platelets per ml of blood perfused as a measure of total deposition. This was still greater for the smaller slides, by a factor of 2.7, averaged for the nearly equal wall shear rates of 300–600 s−1 vs. 280–560 s−1.
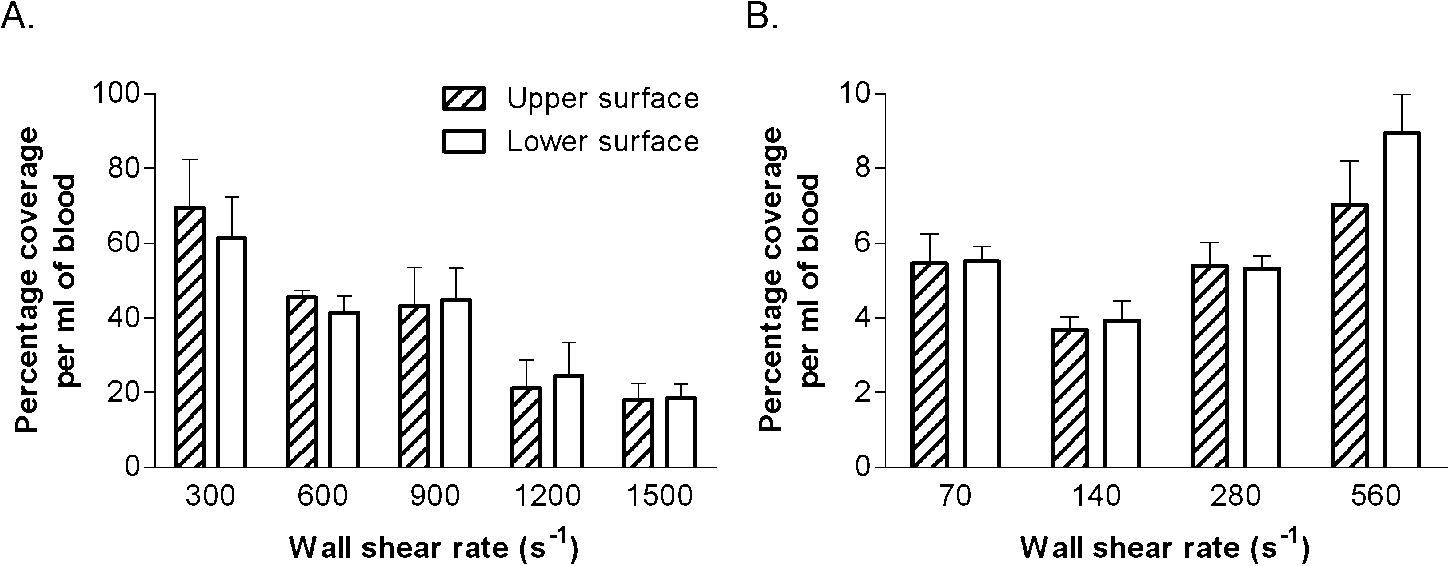
Effects of varying wall shear rate on the adhesion of platelets to collagen when blood was perfused at native haematocrit through, (A) 100 µm, (B) 300 µm microslides; comparison of adhesion to upper and lower surfaces. Data are mean ± SEM for the percentage of the area covered by platelets for 4 (A) or 3 (B) experiments. ANOVA showed significant effects of shear rate on adhesion (
The above comparisons are presented in terms of wall shear rates in the different microslides, calculated assuming parabolic flow between infinite plates. We also measured pressure drop along the microslides, which is directly proportional to the wall shear stress and also allowed calculation of the apparent blood viscosity. This viscosity would not necessarily be equal for a given shear rate in the two differently-sized capillaries because of the Fahraeus–Lindqvist effect [9]. Indeed, while the apparent viscosity of the blood was nearly constant for wall shear rates >300 s−1 and increased at lower shear rates for both sizes of microslide (Fig. 4(A)), the apparent viscosity for the 300 µm microslides was significantly higher than for the smaller microslides. The ratio of viscosities for 300:100 µm microslides varied between about 1.35 and 1.6, and was higher at lower shear rate (Fig. 4(B)).
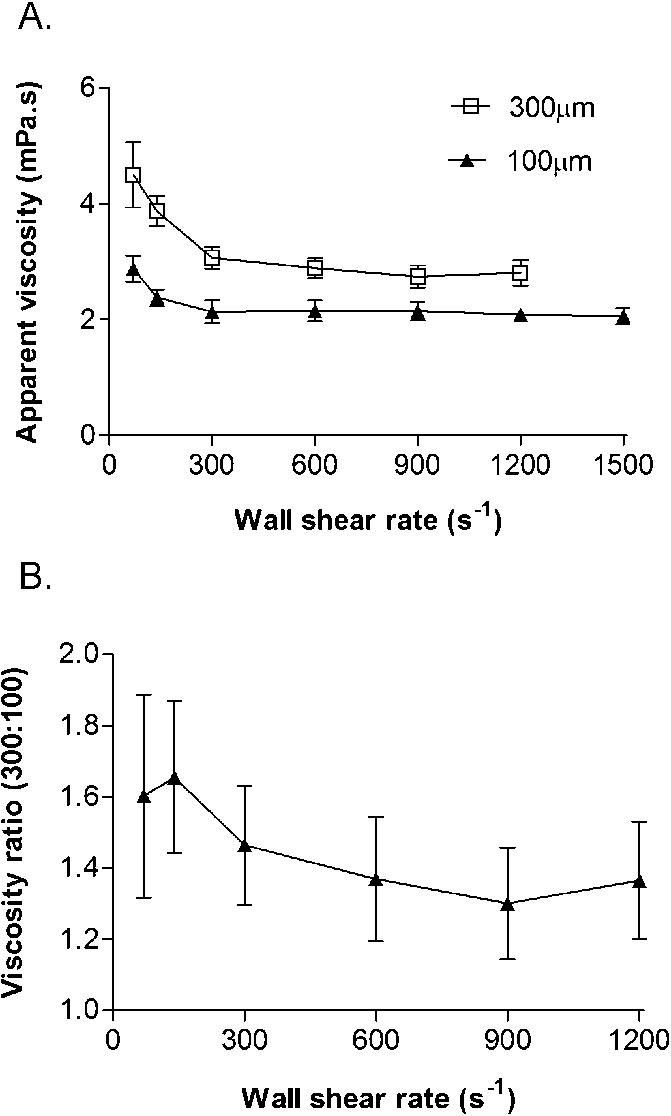
Effects of varying wall shear rate on the apparent viscosity of blood perfused at native haematocrit through microslides of different size. (A) Absolute values for viscosity. (B) Ratio of viscosities for 300:100 µm microslides. Data are mean ± SEM from 3 experiments. ANOVA showed significant effects of shear rate and vessel size on viscosity (A) or shear rate on the ratio of viscosities (B) (
We previously reported that in vertically orientated 300 µm microslides, there was negligible adhesion of leukocytes in the absence of red cells, and that adhesion increased with increasing haematocrit up to 30%, after which it was nearly constant [10]. Based on our earlier observation of equal adhesion on upper and lower surfaces of horizontal microslides at 40% haematocrit, here we studied adhesion to the lower surface of 100 µm microslides for haematocrit varied between 10–40% for a wall shear rate of 140 s−1. We found very high levels of adhesion at 10% haematocrit which decreased progressively for haematocrits of 20% or 30%, but then increased slightly at 40% (Fig. 5(A)). The values at 10 and 20% haematocrit were much higher than 40% (about 3.5-fold in the latter case). Observing the cells that were rolling, their velocity tended to be higher at the higher haematocrits, but this trend was not statistically significant (Fig. 5(B)). Examining the upper surface at the 10% haematocrit, we did not observe rolling adherent leukocytes or fluorescent streaks. This indicated that leukocytes had sedimented away from the upper surface, presumably because at this low haematocrit and low shear rate, red cells had also sedimented so much that they no longer forced leukocytes to the upper surface. There were, however, many leukocytes adhering on the lower surface alone. In a single additional experiment, we directly compared adhesion at 20% or 40% haematocrit for 100 µm and 300 µm microslides, and found about three-fold higher adhesion at 20% than 40% in both sizes.
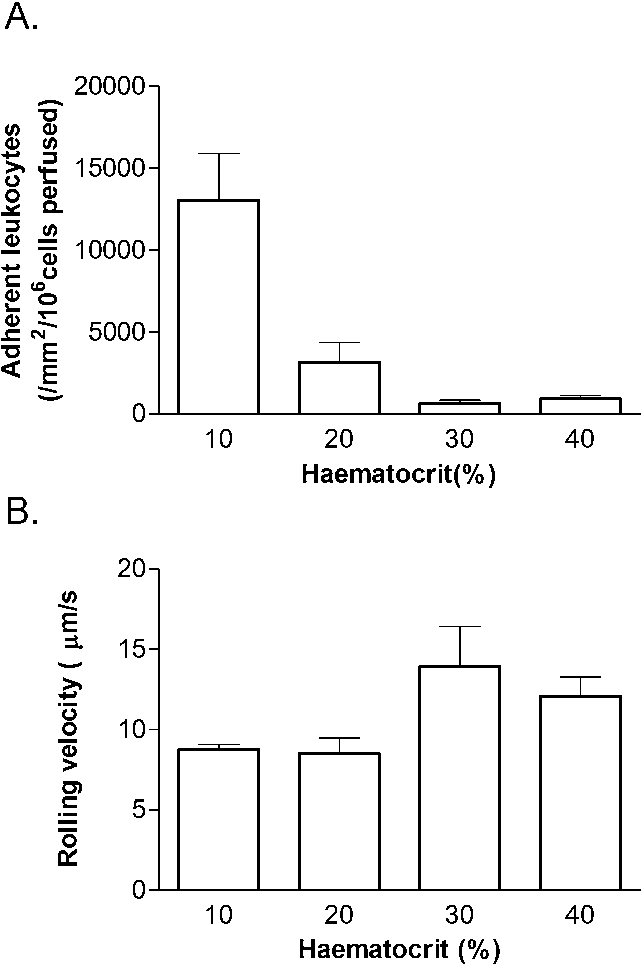
Effects of varying haematocrit on (A) adhesion of leukocytes, (B) rolling velocity of adherent leukocytes, for 100 µm microslides coated with P-selectin and wall shear rate of 140 s−1. Data are mean ± SEM from 3 experiments. ANOVA showed significant effects of haematocrit on adhesion (A) (

Effects of haematocrit on the adhesion and margination of platelets for 100 µm microslides and wall shear rate of 300 s−1. (A) Percentage surface covered by platelets for collagen-coated microslides. (B) Number of free flowing platelets near the wall for albumin-coated microslides. (C) Velocity of free-flowing platelets. Data are mean ± SEM from 4 experiments. ANOVA showed a significant effect of haematocrit on percentage coverage and margination (
Platelet adhesion was also studied on the lower surface of 100 µm microslides for haematocrits from 10 to 50%. At a wall shear rate of 900 s−1, we found a significant, nearly linear increase in platelet coverage as haematocrit was increased from 10–50% (Fig. 6(A)). In contrast to leukocytes, we did not notice loss of adhesion to the upper surface at the lower haematocrits. We thus checked the number and velocity of platelets flowing near the upper surface of non-adhesive albumin-coated microslides as a function of haematocrit. This was done at a wall shear rate of 300 s−1 because at 900 s−1 it became difficult to track individual platelets. There were similar numbers observed at 20 and 30%, although the numbers did tend to increase at 40 and 50% (Fig. 6(B)). However, the velocity of the platelets near the wall did not change with haematocrit (Fig. 6(C)).
Varying haematocrit might influence delivery of cells to the wall, but would also be expected to increase viscosity and hence shear stress. The latter effect might vary in magnitude with shear rate. The apparent viscosity of the blood was thus measured at wall shear rates up to 1500 s−1 using 100 µm microslides and haematocrits from 10 to 50%. Shear rate dependence of viscosity was again evident below 300 s−1, and as expected, viscosity increased with increasing haematocrit at all wall shear rates (Fig. 7(A)). When the effect of haematocrit on viscosity was compared for 140 and 900 s−1 (the wall shear rates used for the adhesion assays), the viscosity was higher for the lower shear rate at all haematocrits, and the ratio of viscosities for 140/900 s−1 was effectively constant at ∼1.12 (Fig. 7(B)).
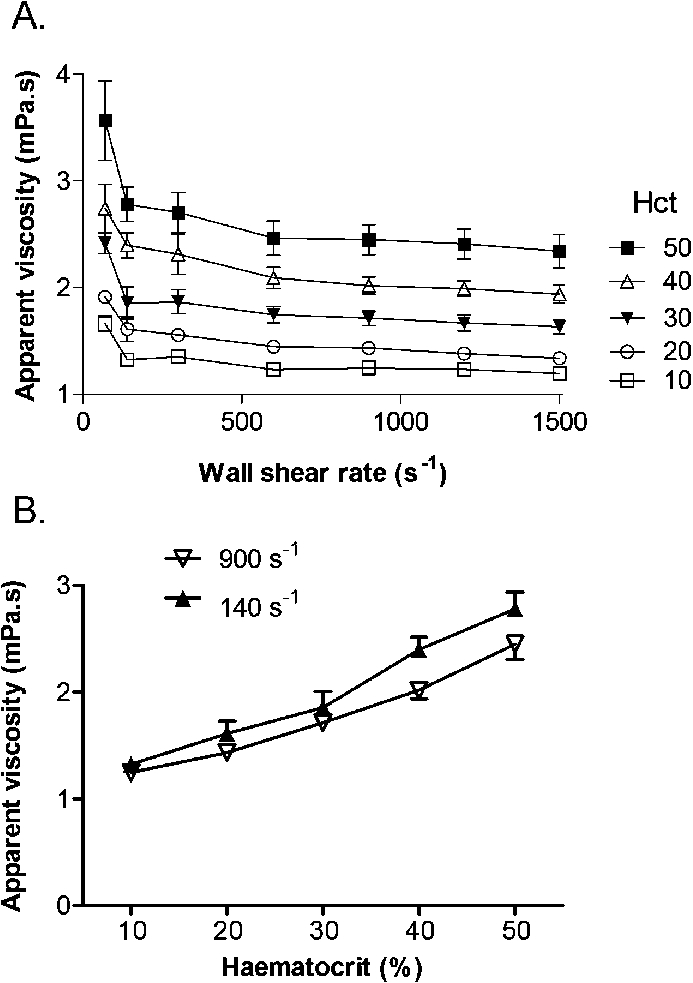
Effect of varying haematocrit on the apparent viscosity of blood in 100 µm microslides. (A) Viscosity shown as a function of wall shear rate for different haematocrits. (B) Viscosity shown as a function of varying haematocrit for wall shear rates of 140 or 900 s−1. Data are mean ± SEM from 3 experiments. In (A) or (B), ANOVA showed significant effects of shear rate and haematocrit on viscosity (
The processes underlying adhesion of leukocytes and platelets to the vessel wall are ‘rheologically evolved’ so that the cells can carry out their functions in different regions of the circulation. Adaptation is manifest at the macro or bulk-flow level, investigated here, and at the micro or molecular level where specialised receptors have evolved specifically for capture from flow [16,17]. Leukocytes are relatively large, rigid blood cells which are only efficiently marginated in presence of red cell aggregation and at low shear rate [8]. Their size also appears to limit the wall shear rates and stresses at which they can successfully form attachments. In contrast, the small size of platelets adapts them to be marginated in the crowded blood at any wall shear rate, regardless of red cell aggregation, and to be efficiently driven into contact with wall in the narrow red cell-depleted marginal zone. Indeed, red cell aggregation impairs their efficacy perhaps because it widens the cell-depleted layer [7]. Their small size also reduces their velocity and shear forces experienced while forming attachments at the vessel wall. Here we extended such observations by considering how extrinsic factors such as vessel size and orientation, and haematocrit interacted to influence capture of the two cells types at the wall of vessels. At haematocrit around 40%, levels of adhesion of leukocytes or platelets were similar on the upper or lower surfaces over a range of shear rates (the range being more restricted for leukocytes than platelets, as expected). In general, adhesion was more efficient in smaller (100 µm) then larger (300 µm) vessels (i.e., relative to the number of cells perfused), but the effect was stronger for the leukocytes than platelets. Increasing haematocrit increased platelet adhesion monotonically, but the effects were more complex for leukocytes. In horizontal tubes at 140 s−1, adhesion was very efficient at low haematocrit but restricted to the lower surface. With increasing haematocrit, adhesion initially decreased but then increased again slightly and was equal on upper and lower surfaces. Interestingly, comparing haematocrits of 20% vs. 40% (roughly equivalent to average microcirculation vs. systemic levels) adhesion of leukocytes at 20% was higher than at 40%, but for platelets the opposite was true.
Several factors may be expected to make adhesion in smaller vessels more efficient per volume of blood perfused. The surface area:volume ratio is greater in the smaller vessel and the distance for cells to be marginated to reach the wall is less, making contact with the wall more likely. For a given wall shear rate, the average blood velocity is also lower in the smaller vessel, and blood residence time longer, again promoting margination and contact. Thus, a greater proportion of cells would be expected to be near the wall in the narrower vessels. We observed a similar absolute numbers of leukocytes near the wall of 100 µm and 300 µm microslides, but this represents a larger proportion of cells present in the smaller vessel.
In addition, there is an implicit assumption in our comparison of the different vessels, that if wall shear rate and stress are matched, the effects on cell adhesion are matched. Thus, for example, it is assumed the near wall velocity will similar for a given wall shear rate. However, there is a secondary effect, particularly for leukocytes, because the cell diameter is not a negligible proportion of the vessel half-depth (especially for the smaller microslides). This means that the shear rate is not uniform across the cell diameter (because of the parabolic flow velocity profile). In effect, velocity increases more rapidly with distance from the wall for the larger vessels. Since cells are predicted to have velocities similar to that for the undisturbed fluid stream at a distance from the wall equal to their radius, this should be greater for the larger vessels. An effect of this nature was seen; leukocytes near the wall were travelling nearly 30% faster on average in the 300 vs. 100 µm microslides. However, the observed cells included some further into the blood stream as well as those grazing the wall, and so this difference is greater than predicted for those grazing. A similar argument applies to the force acting on cells that adhere, which is assumed to be proportional to the wall shear stress. However, fluid shear stresses will drop off more slowly with distance from the wall for the larger vessels, so that again one would predict that leukocytes experience greater force integrated over their surface force for a given wall shear stress in a larger vessel. Thus, efficiency of attachment for given wall shear rates and stresses may be less in larger vessels, especially for leukocytes.
Other likely contributing factors are the Fahraeus and Fahraeus–Lindqvist effects, i.e., haematocrit and apparent viscosity decrease with decreasing vessel diameter below ∼300 µm for a given feed (or systemic) haematocrit [9,18]. Measured viscosity (and hence wall shear stress for a given wall shear rate) was about 50% higher in the 300 µm compared to the 100 µm vessels. This effect would again be expected to affect leukocyte adhesion especially, since their adhesion is more sensitive to this parameter.
The outcome for adhesion of the Fahraeus effect on haematocrit is less easy to predict. We could not directly measure haematocrit in the microslides, but did vary it ‘systemically’. In the case of platelets, the outcome was quite straight forward and in agreement with previous publications [11,12]. There was a monotonic increase in adhesion with increasing haematocrit, and adhesion was similar for the upper and lower surfaces. Adhesion was inefficient at 10% haematocrit, and it seems that increasing red cell concentration increasingly marginates and forces the platelets against the wall, even though viscosity and hence shear stress increases. For leukocytes the outcome of varying haematocrit is evidently complicated by effects of orientation and sedimentation. At 40% haematocrit, adhesion was the same on upper and lower surfaces, and previous studies also indicated little effect of vessel orientation at this haematocrit [10]. At low haematocrit the result was very different. Whereas we previously found lower adhesion at lower haematocrit in vertically-perfused vessels [10], here in horizontal vessels we observed many more adherent leukocytes at 10% haematocrit, although only on the lower surface. Direct microscopic observation indicated that leukocytes had sedimented as none were visible near the upper surface, and presumably red cells had also sedimented leading to a failure of upward margination of leukocytes. Sedimentation of leukocytes in suspension alone perfused though horizontal channels does give highly efficient adhesion on the lower surface alone [14]. In the present case, at 10% haematocrit, it appears that sedimentation of leukocytes was also effective in the presence of dilute red cells, even though one would expect the latter to sediment faster if aggregated. It seems that crowding at the lower surface was not enough to inhibit the leukocyte delivery and efficient adhesion resulted. Even at 20% haematocrit the value was still higher than at 40% (ratio
Other interesting observations related to rolling velocity of leukocytes. This was remarkably constant with increasing shear rate and hence shear stress for either vessel size. Increasing haematocrit (and hence shear stress) in the smaller microslides was associated with faster rolling, but this effects was not statistically significant. In a previous study, we did not detect increased rolling velocity with increasing haematocrit [10]. On the other hand, rolling velocity was slower in the larger vessels. This is even though shear stress is predicted to be greater due to the Fahraeus–Lindqvist effect. However, it has been pointed out that the force applied to cells adhering to the wall in Poiseuille flow depends on the vessel diameter if the cell diameter is a significant proportion of the vessel radius [19]. As pointed out above, the force experienced for a given wall shear stress is greater for the smaller vessel. However, increasing stress is associated with increasing deformation of adherent leukocytes which may stabilise their attachment, and contribute to invariance of rolling velocity with increasing shear rate [20,21]. Overall, the disparity in rolling velocities between the vessel sizes remains incompletely explained.
The experiments described here were practically limited with regard to the vessel dimensions used and indeed shape. Larger vessels would require larger blood volumes to obtain comparable wall shear rates, while smaller capillaries are hard to handle and need very low absolute flow rates, so that e.g., dead volumes become problematic. Practically, we have not used chambers outside the range 100–400 µm in depth for adhesion studies in our laboratory. Commercial microfluidic devices have become available in recent years and used for analysis of cell adhesion, but we are not aware of their use in studies utilising whole blood. The depths of the microslides used (100 or 300 µm) represent relevant scales for arterioles and post-capillary venules but the rectangular shape and width are evidently not representative of blood vessels. The experiments are thus to some extent 2-D representations of phenomena expected to occur in 3-D in circular tubes. The rectangular shape however, provides the excellent optical qualities and larger area in focus needed for the experimental measurements.
The approach taken here has largely been to present adhesion as a function of the volume of blood or number of cells delivered to the ‘vessel’ as a whole, as being most relevant to the physiological efficiency of the process. This does not explicitly show the efficiency of the attachment process itself, because this requires separate knowledge of the transport of the adherent cells to the wall. In a previous report [7], we explicitly addressed this by separately evaluating the number of cells near the wall and the number adhering as functions of shear rate and of red cell aggregation and rigidity. Here we only showed data for numbers of leukocytes marginated near the wall at one wall shear rate for the two microslides or numbers of platelets near the wall for different haematocrits. These assist understanding of the final levels of adhesion, but are not sufficient to allow separate discussion of the control of transport to the wall and adhesion once cells get there. Hence our use of effectively ‘lumped’ data.
These studies provide new insight into the rheological factors affecting adhesion in the circulation, and the adaptation of leukocytes and platelets for their function. Adhesion of leukocytes is particularly strongly influenced by vessel size and, depending on orientation, rather less sensitive to low haematocrit; indeed adhesion in horizontal tubes is effective at very low values. This predicts optimal adhesion in post-capillary venules in inflammation, where the Fahraeus effect means haematocrits are low, and where red cell aggregation will also promote margination and attachment. The dependence of efficiency of adhesion of platelets on vessel size is less marked, but dependence on haematocrit more marked. Allied with the fact that red cell aggregation decreases adhesion, these results are consistent with the requirement for platelets to function efficiently in larger, faster flowing arterial vessels with systemic haematocrit. Conversely, it is likely that ineffective platelet adhesion at low haematocrit contributes to impaired haemostasis in anaemic individuals [22].
Footnotes
Acknowledgements
This work was supported by a research grant EP/C52845X/1 from the Engineering and Physical Sciences Research Council, UK. Tim Watts’ PhD was funded by a studentship from the School of Chemical Engineering.