
Abstract
Background:
Cell–cell and cell–surface adhesion modulated by water-soluble polymers continues to be of current interest, especially since prior reports have indicated a role for depletion-mediated attractive forces.
Objective:
To determine the effects of concentration and molecular mass of the neutral polymer dextran (40 kDa to 28 MDa) on the adhesion of human red blood cells (RBC) to coated glass coverslips.
Methods:
Confocal-reflection interference contrast microscopy (C-IRM), in conjunction with phase contrast imaging, was utilized to measure the adhesion dynamics and contact mechanics of RBC during the initial stages of cell contact with several types of substrates.
Results:
Adhesion is markedly increased in the presence of dextran with a molecular mass ⩾ 70 kDa. This increased adhesiveness is attributed to reduced surface concentration of the large polymers and hence increased attractive forces due to depletion interaction. The equilibrium deformation of adhering RBC was modeled as a truncated sphere and the calculated adhesion energies were in close agreement with theoretical results.
Conclusions:
These results clearly demonstrate that polymer depletion can promote RBC adhesion to artificial surfaces and suggest that this phenomenon may play a role in other specific and non-specific cell–cell interactions, such as rouleau formation and RBC–endothelial cell adhesion.
Introduction
Multiple specific and non-specific forces govern cell–cell and cell–surface interactions. In the past, most research investigating red blood cell (RBC) adhesion onto artificial surfaces has dealt with either specific forces (e.g. lock and key) [1] or non-specific forces (e.g. electrostatic or undulation) [2]. However, only very few reports have considered depletion interaction [3,4].
The reversible aggregation of human red blood cells continues to be of interest in the fields of basic science and clinical hemorheology. RBC form aggregates or rouleaux in vitro when they are suspended in either plasma or buffer solutions containing large neutral polymers [5]. In vivo RBC aggregation occurs at low shear forces or stasis and is a major determinant of low-shear blood viscosity and thus in vivo flow dynamics [6]. Abnormal increases of RBC aggregation have been observed in several diseases associated with vascular disorders (e.g., diabetes mellitus, hypertension) [7].
There are presently two co-existing yet mutually exclusive proposed mechanisms for RBC aggregation. One is the so-called bridging model in which red blood cell aggregation occurs when the bridging forces due to the adsorption of macromolecules onto adjacent cell surfaces exceed disaggregation forces (i.e. electrostatic repulsion, membrane strain and mechanical shearing) [8]. Another mechanism that has been proposed is the so-called depletion model, which is based upon the formation of a depletion layer near a surface in contact with a polymer solution; this layer forms if the loss of configurational entropy of the polymer is not balanced by adsorption energy. Within this depletion layer, the polymer concentration is lower than in the bulk phase. Thus, as two particles approach, the difference in solvent chemical potential (i.e., the osmotic pressure difference) between the intercellular polymer-poor depletion zone and the bulk phase results in solvent displacement into the bulk phase and hence depletion interaction [9]. Due to this interaction, an attractive force develops that tends to minimize the polymer-depletion space between the cells, thus resulting in flocculation or aggregation.
The existence of depletion layers at RBC surfaces has been confirmed via particle electrophoresis studies of cells suspended in various polymer solutions [10] and the energetics of RBC interaction in neutral polymer solutions and the effects of the glycocalyx have been reported [9]. More recently, qualitative evidence has shown enhanced contact to albumin-coated glass surfaces for RBC suspended in 500 kDa dextran solutions due to depletion interaction [11]. Experimental findings showing that RBC aggregation and electrophoretic mobility (EPM) are affected by the neutral polymer dextran suggest that the dextran concentration and the electrical potential profile near the interface between the membrane and the continuous phase are important determinants of adhesive behavior.
The present study was undertaken to develop a quantitative understanding of the adhesion energies involved in depletion-induced RBC adhesion. For this purpose, confocal-reflection interference contrast microscopy (C-IRM), in conjunction with phase contrast imaging, were utilized to measure the adhesion dynamics and contact mechanics of RBC during the initial stages of cell contact with several types of substrates [12]. Our results are in agreement with existing literature, and suggest that depletion may be an important determinant of the known increases of RBC adhesiveness and aggregability in the presence of non-adsorbing polymers.
Materials and methods
Materials
3-aminopropyl-triethoxy-silane (APES), anhydrous acetone (99.5%), bovine serum albumin (BSA), dextran (average molecular weight fractions from 40 kDa to 28 MDa), bovine serum albumin (BSA) and fibrinogen were purchased from Sigma Aldrich, St. Louis, MO and used as received. Isotonic phosphate buffered saline (PBS) was obtained from 1st Base Pte Ltd, Singapore.
RBC preparation
Fresh blood was obtained by sterile finger prick into a 1.5 ml Eppendorf centrifuge tube and used within four hours. RBC were separated from whole blood by centrifugation (1,000g, 10 min) and washed twice in PBS with 0.2% BSA. RBC were then suspended in either dextran-free PBS or in dextran-PBS solutions. Microscopic observations performed within the four hours indicated no morphologic changes from the normal biconcave shape.
Substrate preparations
The method for attaching a positive amine surface, termed amination, to glass coverslips has been described elsewhere [11]. Briefly, round coverslips (VWR, Micro Cover Glasses, Round, No. 1 Dia. 25 mm) were soaked in 69% nitric acid overnight and washed thoroughly with distilled water for 5–10 min. The remaining water was removed with tissue and compressed air, the coverslips sonicated in anhydrous acetone for five min and rinsed once in fresh acetone. They were then dipped into a 2% (v/v) solution of APES in anhydrous acetone for 30 min, rinsed with acetone followed by distilled water, dried at 50°C, and stored in a dust-free environment until use. To coat the aminated coverslips with protein, they were incubated with 2% bovine serum albumin or 2% fibrinogen in PBS at room temperature for two hours to allow binding to APES. The modified coverslips were then briefly rinsed with PBS, and remained in contact with PBS until utilized. Coverslips without APES were prepared using the same protocol. Using this approach, six different surfaces were available for testing: three untreated with APES (cleaned only, BSA coated, fibrinogen coated) and three treated with APES (APES only, APES+BSA, APES+Fibrinogen).
Cell microscopy
RBC suspended in dextran free PBS or dextran-PBS were transferred to the surface of coverslips secured in a chamber that was then fixed on a chamber holder and placed on the stage of an inverted microscope. The system was equipped with a temperature control chamber (SEC Engineering, Korea) and all images were acquired at room temperature. The bottom of the coverslips was in contact with an oil immersion objective, thereby allowing: (1) phase contrast images in order to investigate RBC shape and spreading; (2) C-IRM images to observe the contact dynamics of adherent cells.
RBC shape and spreading dynamics
The shape of adhering RBC were evaluated using a 510 Meta confocal laser scanning microscope system (C-LSM, Carl Zeiss, Germany) equipped with a 63× oil-immersion objective (Neofluar, NA = 1.25) and a cross-polarized and transmitted light analyzer (Carl Zeiss, Germany). A series of phase contrast images were obtained and analyzed using an image analysis software program (SM5 Image Browser, Carl Zeiss, Germany) to determine “spreading area” (see Section 3).
Contact dynamics of adherent cells
The C-LSM microscope was also used to study detailed interactions between RBC and the surface of interest. Light from an argon-ion laser with a maximum power of 1 mW and an excitation wavelength of 488 nm was focused through the microscope objective (69 µm pinhole) on the cell–substrate interface through the microscope objective to record images via C-IRM. RBC suspended in PBS or PBS-dextran were deposited on coverslips and cells allowed to settle for three minutes, following which a series of C-IRM images were captured to monitor the contact dynamics of adherent cells.
In the C-IRM images, close contact zones between the RBC and the surface appear as a dark region with interference patterns, often ring-like, formed by polarized light reflected from the upper surface of the coverslip and the lower surface of the RBC; SM5 analysis software was used for all image acquisition and analysis. The close contact area was determined by drawing a region of interest (ROI) with a PC mouse-based sub-routine. The normalized contact area of adhering cells was then calculated as the ratio of the contact area to the average mid-plane area, as measured from phase contrast images, at various time points.
Data analysis
The details of modeling the contact mechanics have been reported previously [13]. In brief, the equilibrium geometry of an RBC adhering to a non-deformable substrate is modeled as a truncated sphere with a mid-plane radius R and
It has been shown previously that the adhesion energy, W, is [14]:
By taking the square root of the ratio of the projected area obtained from phase contrast images and the adhesion contact area from C-IRM microscopy,
Results and discussion
Different surface modifications
RBC adhesion onto artificial surfaces is known to be strongly dependent on the surface properties of both the RBC membrane and the surface itself. Thus, it is necessary to clearly recognize and minimize non-specific interactions in order to evaluate the effects of different surfaces [16]. Figure 1 shows C-IRM images of RBC interacting with six different surfaces and Table 1 presents calculated area and deformation results obtained from such images. As is evident by the extent of the dark areas, RBC binding to APES-only coverslips results in about a 2-fold increase of close contact area as well as a significantly larger degree of deformation (
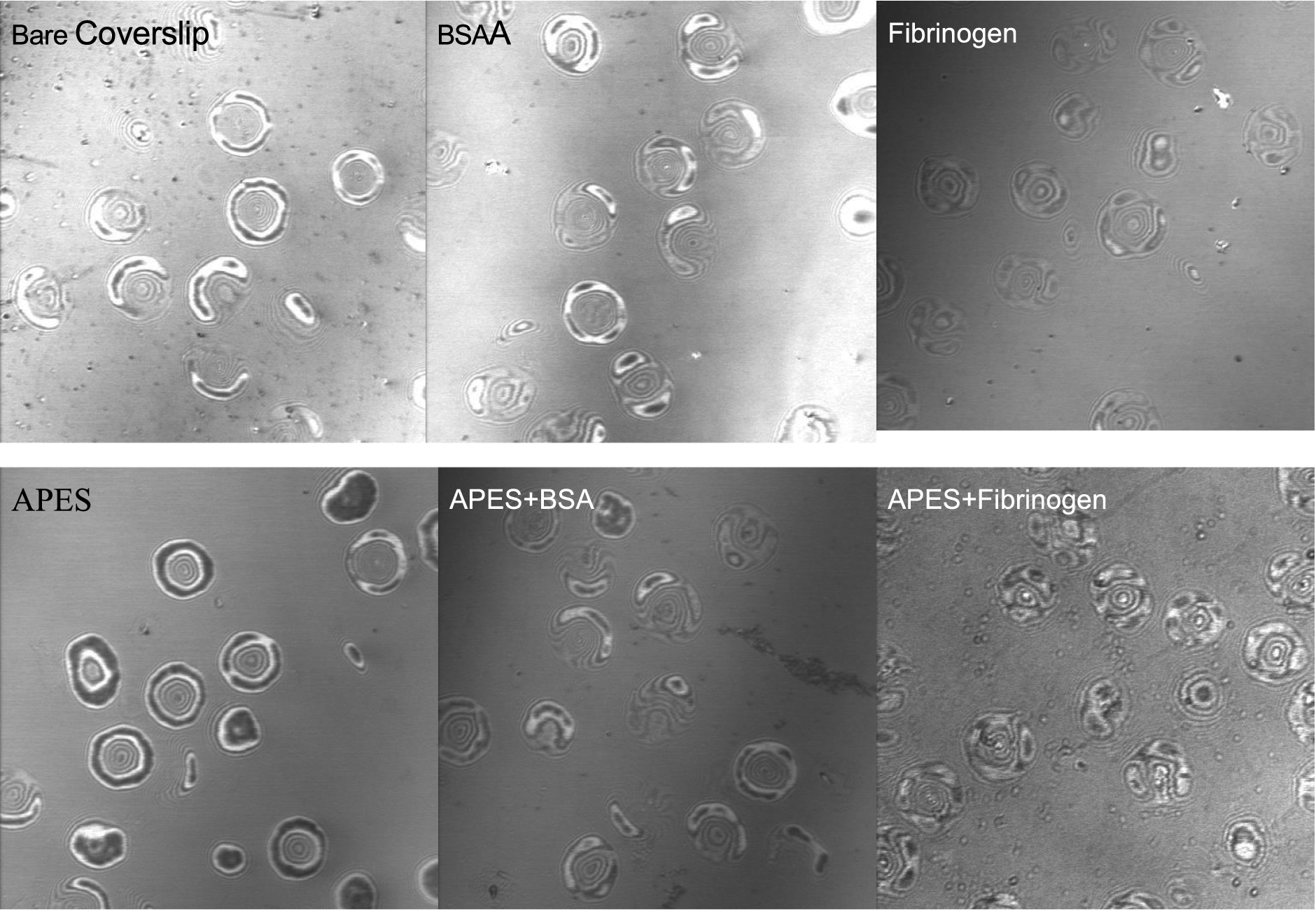
Representative confocal interference reflection microscopy (C-IRM) images of RBC settled onto six different surfaces; RBCs were suspended in dextran-free PBS. All images were taken at 5 min after placing RBC suspension onto coverslips. Darker areas of cell–surface images indicate smaller distances between RBC and surface. Top three panels are for surfaces not treated with APES (cleaned only, BSA coated, fibrinogen coated, left to right) and bottom three panels treated with APES (APES only, APES+BSA, APES+Fibrinogen, left to right).
RBC spreading area, contact area and degree of deformation for six different surfaces for cells suspended in dextran-free solution
Notes: Data are mean ± SD,
RBC adhesion onto BSA-coated surfaces (bare or APES pre-treated) is intermediate, with no apparent change of the cell morphology (i.e., spreading area): RBC spreading areas are comparable for all six surfaces (Table 1). In terms of contact area, RBC bind least to bare glass or fibrinogen, bind intermediately to BSA or APES-BSA or APES-fibrinogen and, as mentioned above, bind strongly to APES-only coating. This strong binding to APES-only surfaces mainly originates from the electrostatic attraction between the aminated glass coverslip and negatively charged RBC membrane; such behavior is consistent with previous reports [11,17]. Given this effect of APES-only surfaces, APES-BSA surfaces were employed to investigate depletion-induced adhesion in order to minimize the contribution of attractive electrostatic interactions between the surfaces.
C-IRM in combination with phase contrast microscopy is an unique technique that allows visual examination of cell–surface contacts and separation, thus making it an interesting tool for studying depletion-mediated cell adhesion to artificial surfaces. Figure 2 presents a time series of phase contrast images (upper panels) and corresponding C-IRM images (lower panels) of RBC suspended in dextran-free PBS adhering to BSA-coated coverslips over a period from 9 min to 61 min. The phase contrast images indicate that RBC retained their normal biconcave morphology over the entire period and hence without a meaningful change of contact area (see also Fig. 4). Note also the almost complete lack of dark areas (i.e., close contact areas) in the lower C-IRM panels even after 61 minutes.

A temporal series of images of RBC in dextran-free PBS settled onto to a BSA-coated coverslip from 9 min to 61 min after application of the cell suspension. The top section of each image was obtained via phase contrast light microscopy and the lower portion via C-IRM.
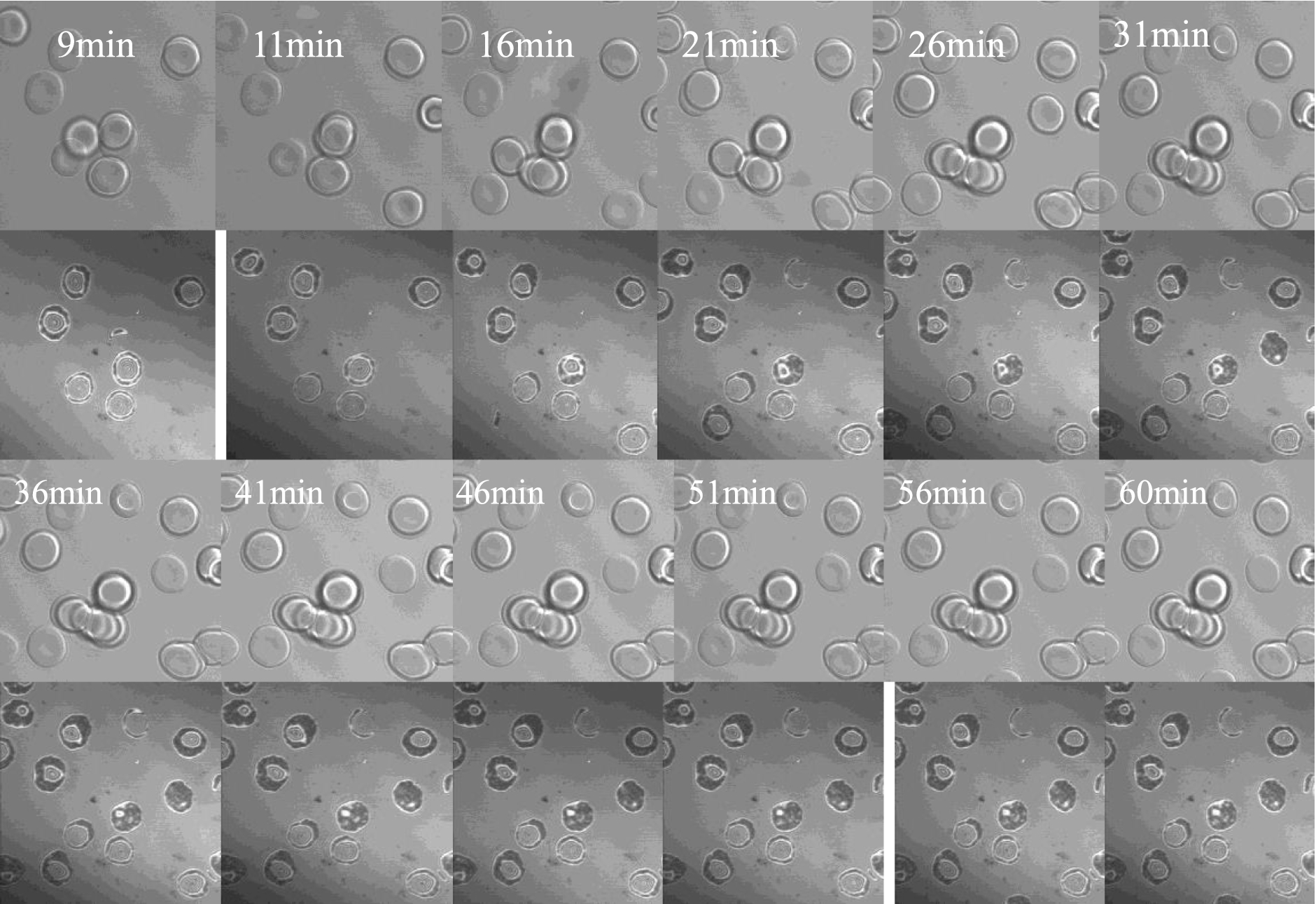
A temporal series of images of RBC in PBS containing 0.5 g/dl 500 kDa dextran settled onto to a BSA-coated coverslip from 9 min to 61 min after application of the cell suspension. The top section of each image was obtained via phase contrast light microscopy and the lower portion via C-IRM.
In marked contrast to cells in PBS (Fig. 2), RBC suspended in PBS-dextran (0.5 g/dl 500 kDa) exhibited stronger RBC adhesion to BSA-coated coverslips, with the extent of adhesion increasing with time as evident by the increased amount of darker areas (Fig. 3). During the initial stages of observation for cells in PBS-dextran (
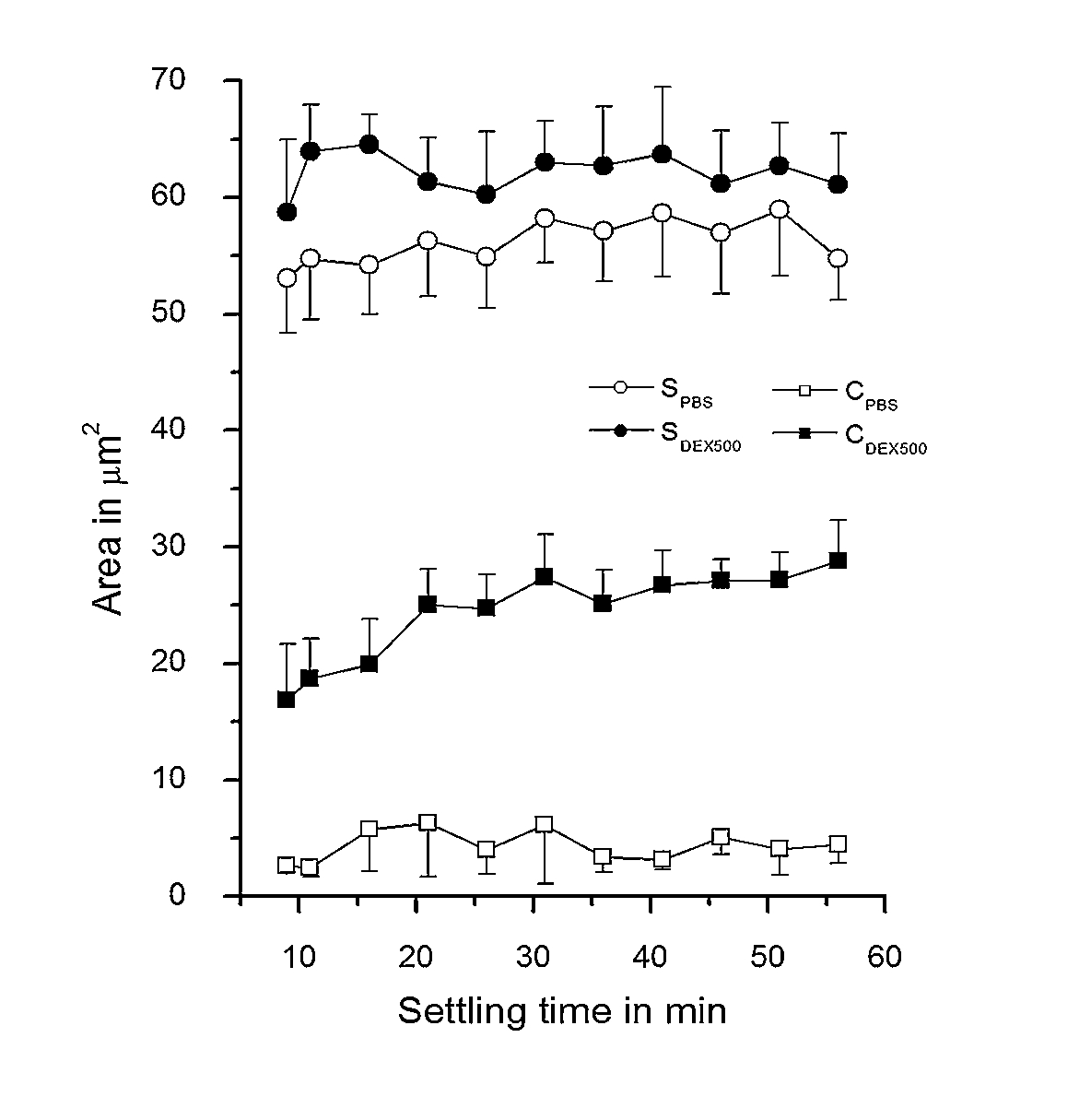
Dependence of the contact (C) and spreading area (S) of RBC in polymer-free solution (PBS) and in 0.5 g/dl dextran 500 kDa (DEX500) on their settling time. Data are mean ± SD,
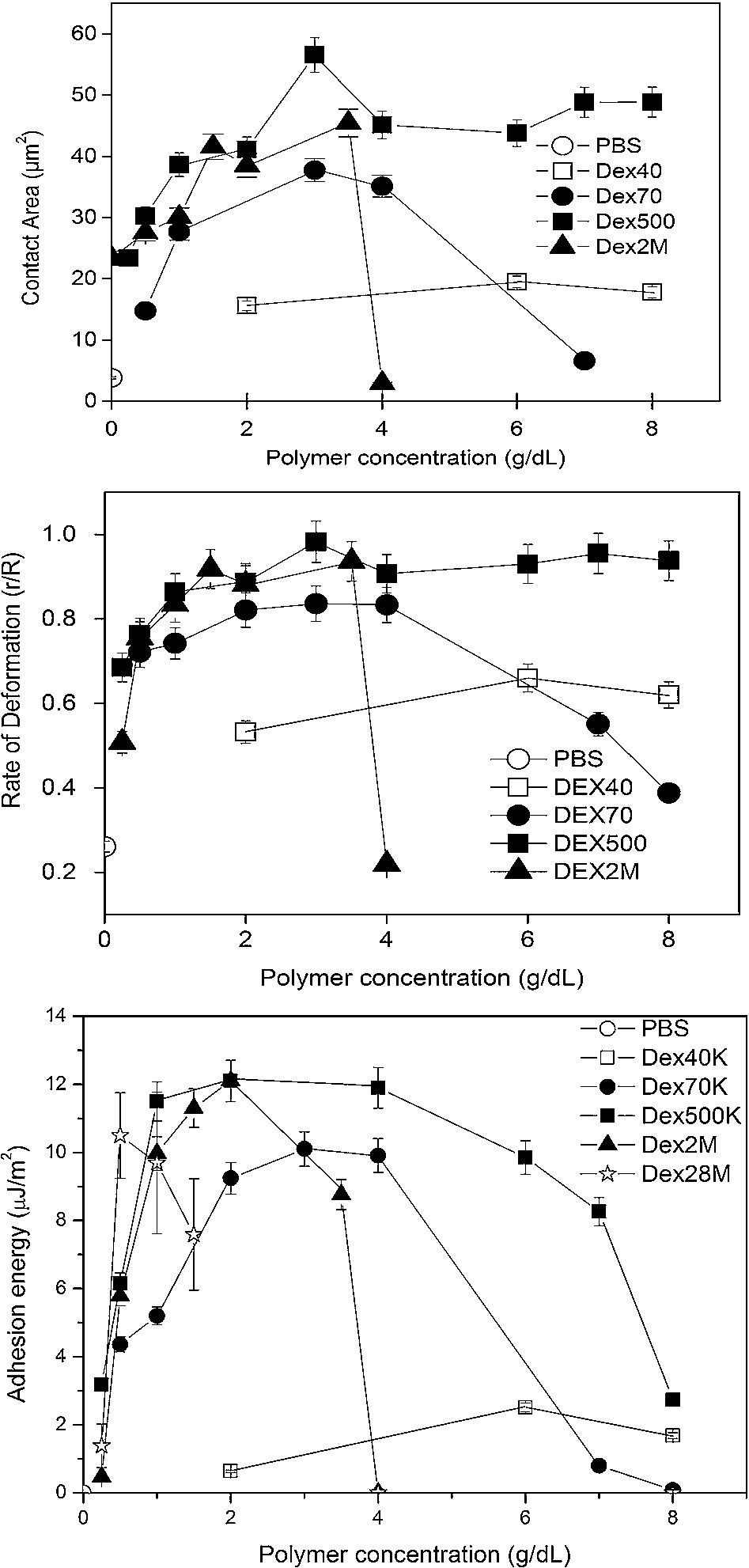
Experimental findings for RBC suspended in various PBS-dextran solutions as a function of polymer molecular mass and concentration: (top) contact area; (middle) degree of deformation; (bottom) adhesion energy. Data are mean ± SD,
Figure 5 presents RBC contact area, deformation and adhesion energy data for cells in PBS-dextran solutions of various dextran fractions (40, 70, 500 kDa and 2 MDa) interacting with BSA-coated surfaces. The contact area results (Fig. 5(top)) clearly show that both molecular mass and concentration have a marked impact on the adhesion of RBC: (a) the contact area for 40 kDa was low and essentially independent of concentration; (b) 70 kDa had a higher contact area and a greater sensitivity to concentration such that for concentrations greater than ∼3 g/dl, the contact area decreased from 37 to 5 µm2; (c) as concentration increased, contact area for cells in 500 kDa increased to a maximum then decreased and plateaued; (d) contact area–concentration relations for 2 M kDa were similar to 70 and 500 kDa below 4 g/dl but decreased to near zero at a concentration of 4 g/dl with only a few adherent RBC detected from both phase contrast and C-IRM images.
Figure 5(middle) shows the average extent of deformation as indexed by the
Figure 5(bottom) presents adhesion energy W values calculated from the truncated sphere model as a function of bulk polymer concentration for PBS only as well as PBS containing 40 kDa, 70 kDa, 500 kDa, 2 MDa or 28 MDa dextran fractions. The calculated values range from 0 to 12 µJ/m2. Inspection of these results indicates: (1) polymer concentration–adhesion energy relations are concave to the concentration axis; (2) maximal values of W increase with increasing molecular mass, although 500 kDa and 2 M kDa have almost the same maximal energy; (3) for larger dextrans (e.g. 70 or 500 kDa) at higher concentrations, W-concentration relations are fairly wide with a somewhat greater width for 500 kDa and a stronger concentration dependence for 70 kDa; (4) adhesion energies for 2 MDa dextran sharply decrease for concentrations greater than 2 g/dl.
The results above clearly indicate that RBC adhesion depends strongly on polymer molecular mass as well as concentration: (a) in the ascending region of the energy-polymer concentration region, total adhesion energy (
Lastly, it should be emphasized that abnormal increases of RBC aggregation have been observed in several diseases associated with vascular disorders (e.g., diabetes mellitus, hypertension) [18,19], and it is known that RBC aggregation is primarily determined by the plasma levels of pro-aggregating macromolecules or proteins and by RBC aggregability [20]. Therefore, it can be concluded that our results may have potential implications for pathological conditions that have been linked to vascular problems. Use of C-IRM in this study not only strongly supports the concept of depletion mediated RBC interactions, but also shows that this method is a simple and unique microscopic technique allowing visual examination of cell/surface contacts and separation in real time; our experience indicates that it is an excellent tool for studying depletion mediated cell adhesion to artificial surfaces [21].
Footnotes
Acknowledgements
We thank Prof. Vincent Chan for providing us with the special chamber and Beiyi Chen for providing us with a computer program to calculate the adhesion energies. This work was supported by BMRC-NMRC joint grant.