
Abstract
The rheological properties of synovial fluid (SF) are largely attributed to the presence of high molecular weight hyaluronan (HA). However, rheological differences between SF and pure HA solutions suggest that SF proteins actively contribute towards the bulk viscoelasticity of this biological fluid. Due to various experimental challenges involved with the rheometry of low-viscosity biological fluids, the macromolecular interactions in SF and their relative rheological importance are still a matter of active discussion. Interestingly however, recent evidence suggests that the concentration and structure of proteoglycan 4 (PRG4, also known as lubricin) can directly modulate the viscoelastic properties of HA-PRG4 solutions. The objective of this review is to highlight recent rheological studies that examine the macromolecular interactions between HA and proteins in SF. First, a general overview of the chemical composition of SF and the molecular structure of its key constituents HA and PRG4 is provided. Subsequently, diverse rheological experimental techniques that have been developed to characterize HA solutions are discussed. Finally, rheological investigations of macromolecular interactions between HA, serum proteins, and PRG4 are examined. This review illustrates how diverse rheological techniques can expand our understanding of the composition–structure–function relationships in SF.
Keywords
Introduction
Synovial fluid rheology and joint lubrication
Synovial fluid (SF) is the clear and viscous fluid that fills all cavities within synovial joints, where it functions as the body’s natural lubricant that protects tissues from both damage and wear during motion. As a result of a combination of compressive loadings and sliding motions, there is a complex interplay between articular cartilage and SF that gives rise to various lubrication mechanisms that depend (to varying degrees) on both the biochemical properties of SF and its bulk rheological behavior. In an effort to better understand how changes in SF composition relate to its mechanical properties, and ultimate lubricating ability, more than half a century of rheological studies have extensively examined the viscosity and linear viscoelastic moduli of SF under physiologically-relevant conditions [1–11].
SF is highly viscoelastic and strongly shear-thinning [4]. In joints like the knee, it is estimated that it can be exposed to rates of shear strain up to 1000 s−1 [12], as well as frequency variations ranging from 1–9 rad/s [13]. SF exhibits a high viscosity at low shear rates and frequencies, which allows it to act as a load-bearer that stabilizes the articular capsule during very small movements at low speeds (e.g. shifts in position). In this regime, SF features a pseudo-Newtonian plateau characterized by zero-shear viscosities on the range of 10–100 Pa · s, with ∼10 Pa · s considered to be normal [10,12,14,15]. At higher shear rates and frequencies (characteristic of full joint movements like walking or running), the viscosity of SF decreases to ∼0.01 Pa · s [10]; this thinning contributes to the formation and stability of a fluid film that effectively separates cartilage surfaces during hydrodynamic lubrication. Recent studies have also found that SF has pronounced elastic and strain-hardening properties; these likely confer joints with shock-dampening characteristics during sudden compressive loads and impacts [10,15–17].
Macromolecular interactions and synovial fluid rheology
The viscoelastic properties of SF have been largely attributed to the presence of the high molecular weight polysaccharide hyaluronan (hyaluronic acid, HA) [8,9,18]. This linear polymer forms a loosely-entangled network that can undergo significant degrees of deformation and whose viscoelastic response is significantly affected by changes in its concentration and molecular weight [2,3,5,10,18–22]. Nevertheless, the physicochemical characteristics of the HA network alone are not able to fully explain the rheological properties of SF [6,9]. Studies have shown that protein constituents in SF can significantly contribute to its mechanical properties, particularly its bulk viscosity and elasticity [6,9]. This may reflect the existence of HA-protein complexes that form as a result of protein aggregation or due to the formation of a soluble complex between HA and the proteins [10]. Over the years, researchers have examined the role of SF proteins like albumin, γ-globulins, fibrinogen, and proteoglycan 4 (PRG4, also known as lubricin), and their relative importance in the rheology of SF [9–11,17,23,24]. However, their specific interactions with each other and with HA, as well as their ultimate effect in the bulk rheology of SF, is still a matter of active discussion [11]. Recent years have been witness to substantial advances in the field of rheology which have provided scientists with novel ways to examine these macromolecular interactions in SF and other complex biological fluids.
Objectives and organization
The objective of this review is to examine recent rheological studies that provide insight into HA-protein macromolecular interactions in relevance to joint lubrication. The remainder of this review is organized as follows: in Section 2, we describe the chemical makeup of healthy SF and describe HA and PRG4 in terms of their structural properties. In Section 3, we introduce the rheological techniques that have been recently used to characterize HA solutions before transitioning into studies of their macromolecular interactions with serum proteins and PRG4 in Section 4. Finally, Section 5 will provide a general outlook for future investigations of SF rheology.
Synovial fluid composition
Healthy SF is a clear, pale yellow viscous liquid that is slightly alkaline pH (7.3–7.64) and is mostly composed of water (960–988 g/kg) [25]. Biochemically, it is an ultrafiltrate of blood plasma that has retained most of its electrolyte and low molecular weight organic molecule composition; however, it has only a third of its original serum protein composition [25] and is enriched with locally-synthesized biomolecules like HA and diverse proteins [26] (Table 1).
Concentration of main constituents in normal synovial fluid
Concentration of main constituents in normal synovial fluid
HA was first isolated from the vitreous humor of cattle eyes [40] in the mid 1930s, but it was not until the 1950s that Ogston and Stanier [1] first attributed the viscoelastic and lubricant properties of SF to its presence. This naturally-occurring polymer is ubiquitously distributed throughout the extracellular matrix of vertebrate tissues, with the highest concentrations found in soft connective tissues [41,42]. In SF, HA is produced by synoviocytes [36] and, structurally, it is composed of disaccharides of alternating D-glucuronic acid and N-acetyl-D-glucosamine linked by β-
In aqueous solutions, HA takes up a loosely coiled wormlike configuration that may be stabilized by dynamically forming and breaking intramolecular hydrogen bonds parallel to the chain axis [21]. This configuration, combined with its high molecular weight and numerous mutually-repelling anionic groups, allows HA to adopt an expanded random coil structure. HA coils can have a radius of gyration of ∼300 nm which, due to their rapid motions, can be effectively described as highly hydrated spheres that occupy hydrodynamic volumes ∼1000 times larger than those occupied by the dry polymers themselves [44–47]. The hydrodynamic volume of individual HA coils increases with molecular weight (
Proteoglycan 4
Tribological studies have found that cartilage lubrication at near-zero sliding speeds (boundary lubrication) is dominated by macromolecular interactions between HA, PRG4, type II collagen fibrils, and phospholipids [52–58]. In the joints, PRG4 forms end-grafted brush structures on cartilage surfaces that lower the adhesion energy between the surfaces [59]. In particular, friction tests in latex-glass bearings [60] and in cartilage–cartilage in vitro friction tests [61] point towards specific synergistic interactions occurring between HA and the boundary lubricant PRG4 (also called lubricin). This large and complex glycoprotein was first purified from bovine SF (BSF) in the 1980s [62]. In SF, it is secreted by synovial fibroblasts and superficial/intermediate-zone chondrocytes [63–66]. Structurally, PRG4 has a protein core that is 1,404 amino acids long with globular N- and C-terminal domains and a central mucin domain (Fig. 1) [67]. The central mucin domain is extensively glycosylated via O-linked β(1-3)Gal-GalNAc oligosaccharide side chains [68,69] that have been shown to be predominantly sialylated [70]; this gives the mucin-like central domain a high negative charge that allows it to generate repulsive hydration forces and/or charge repulsions [71]. The globular amino- and carboxy-terminal domains allow for intra- and inter-molecular disulfide bonding of PRG4 subunits [59,72,73], which are known to affect its boundary lubricating ability [74] and may play an important role in PRG4’s stability and aggregation kinetics [75].
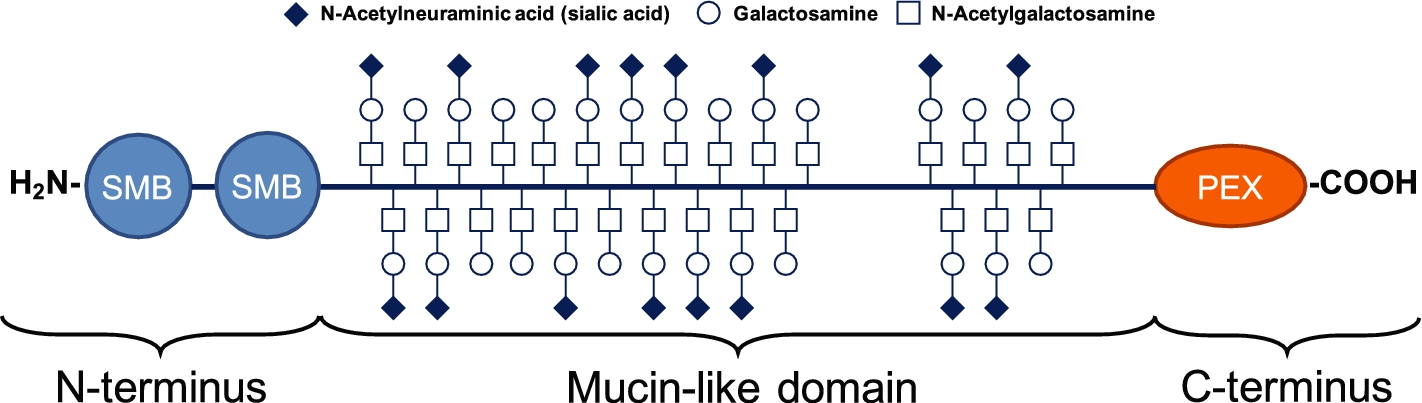
Structure of a PRG4 monomer. Schematic depicts the domain structure of PRG4, with its central region (mucinous and heavily glycosylated) being flanked by the amino-terminal at the left and by the carboxyl-terminal at the right. The amino-terminal contains 2 domains similar to the somatomedin B (SMB) domains of vitronectin. The carboxyl-terminal is a haemopexin (PEX) domain. The globular domains form disulfide bonds with other PRG4 subunits. Adapted from Rhee et al. [67] and Estrella et al. [76].
The mucinous nature of its central domain makes PRG4 different from other proteins present in SF. Mucin solutions are known to have a viscosity proportional to their concentration [77], with small differences in mucin concentration causing significant changes in mucus viscoelasticity [78]. Furthermore, disulfide bonded multimerization is necessary for gel behavior [79,80], therefore, the absence of specific disulfide bonds could lead to partially unfolded PRG4 structures that are prone for aggregation.
Bulk shear rheology
The viscoelastic behavior of HA solutions has been widely characterized in the past using steady and dynamic shear rheometry [51,81–83]. In general, HA solutions exhibit a rheological behavior typical of non-associating or loosely cross-linked polymers [51,84,85] which, unlike SF, do not exhibit a shear history dependence of viscosity [86,87]. This non-ideal behavior arises from mutual macromolecular crowding, in which the crowding of highly expanded HA chains forces individual domains to interpenetrate and entangle with each other [21,22,42,88]. As a result of the crowding and the resultant excluded volume effect, the effective concentration of HA molecules increases dramatically with increasing concentration and molecular mass. At the concentration and molecular mass of HA in semidilute solutions like SF, HA molecules confer solutions with a high viscosity at low shear rates and, as shear rates increase, these rigid molecules gradually line-up with the flow and offer less resistance to it, conferring solutions with a shear-thinning behavior.
In an effort to predict their rheological behavior, several authors have established structure–property relationships for HA solutions that correlate various molecular parameters to measured viscosities [43,81,89–97]. Using size exclusion chromatography and multi-angle light scattering in conjunction with capillary viscometry, Meyer et al. [43] established a relationship between the intrinsic viscosity
The behavior of HA solutions as a response to oscillating shear flows has been comprehensively examined in the past [98]. At low frequencies, the macromolecular network has a dominant viscous behavior (
Different arthropathies are known to decrease the concentration of HA in SF [29,39,101], shift its HA molecular weight distribution to lower ranges [102,103], and increase its total protein and lipid content [8]. In patients with inflammatory joint diseases (e.g. rheumatoid arthritis), SF features a reduced viscosity and a quasi-Newtonian bulk behavior [8], which reflects an increased degradation of HA molecules that leads to a decrease in high
Extensional rheology
Extensional flow refers to a type of flow deformation that involves the elongation, or stretching, of a fluid. These ideally shear-free flows are useful in polymer characterization because they can generate high molecular stretch/orientation flow fields that are very sensitive to the molecular structure of polymeric systems. Compared to shear flows, extensional flows have a stronger effect on flexible macromolecules because the latter unravels and stretches macromolecules uniaxially whereas the former superimposes rotation with stretching in all directions [16]. In general, the deformation of macromolecular structures occurs when the extensional strain rates (
When subjected to strong extensional flows (
Recent extensional studies provide evidence that HA solutions are significantly strain-hardening under elongation. Bingol et al. [10] examined low- and high-
To overcome these limitations, extensive characterizations of HA solutions under planar elongational flows have been done using microfluidic cross-slot devices that measure flow-induced birefringence in order to quantify macromolecular deformations [17,97,110]. These cross-slot devices impose a planar extensional flow field with a stagnation point that elongates fluid elements in either the y or x directions during each cycle, with enough residence time given in each direction to ensure fully-stretched macromolecules (see Fig. 2). Using an shape-optimized version of these devices, Haward [97] characterized the extensional viscosities and extensional relaxation times of dilute HA-PBS solutions over a range of concentrations (0.1–1.0 mg/ml) and molecular weights (0.9–4.8 MDa). In this study, the author compared shear and extensional strains by using the dimensionless Trouton number
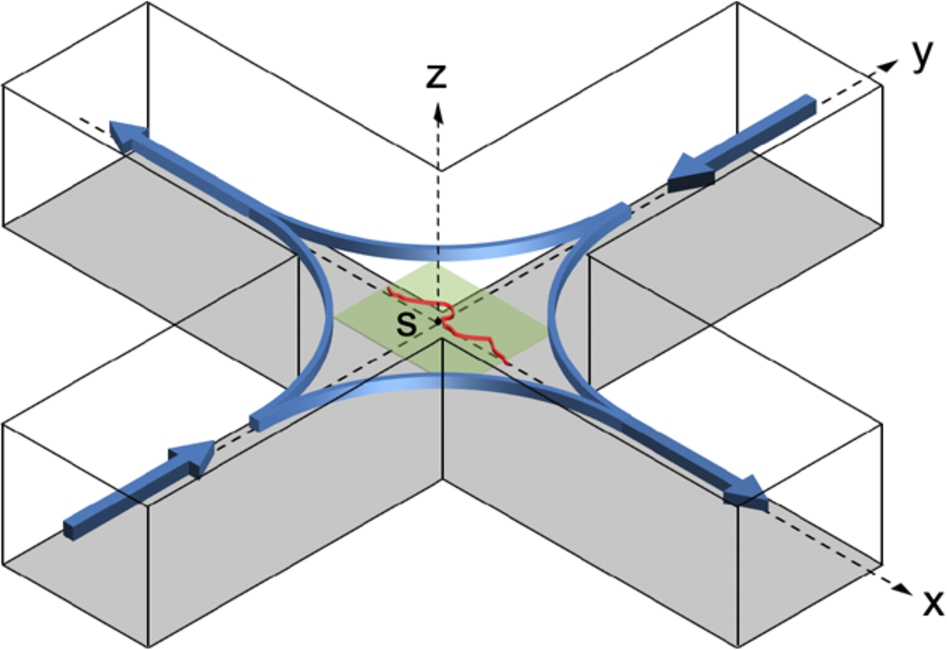
Microfluidic cross-slot device for producing planar elongational flow with stagnation point. Schematic indicates the coordinate system with the origin at the stagnation point (marked with an “s”) and the flow direction indicated with arrows (blue). A polymer molecule (red) is stretched close to the stagnation point and extended along the x-axis. Extensional rheological data is determined from flow-induced birefringence measurements at the vicinity of the stagnation point (shaded green plane) by using the stress-optical rule. Adapted from Haward [97].
Shear flows have been used throughout the years to determine the dynamic viscosity and linear viscoelastic rheological properties of diverse complex fluids like SF. Usually, a small amplitude sinusoidal strain/stress wave is applied to a fluid and, by examining the frequency phase shift of the fluid’s response shear/strain wave, one is able to decompose it into its elastic and viscous properties. However, the analysis of linear viscoelastic data is strongly limited to a very small range of deformations; outside this regime, complex fluids exhibit nonlinear responses that cannot be as easily decomposed into their elastic and viscous components. Given the wide range of shear strain deformations and frequencies to which SF is exposed in vivo, a full rheological characterization requires well-defined nonlinear test protocols to examine its nonlinear viscoelastic behavior [112]. In the case of HA solutions, only a few studies have examined their nonlinear viscoelastic response under large amplitude oscillatory shear (LAOS) flow [113,114].
Zhang and Christopher [114] used a double wall cylinder geometry (rotor inner diameter 32 mm, rotor outer diameter 35 mm, and rotor height 53 mm) to impose LAOS-strain deformations to physiologically-relevant HA-PBS solutions (3.4 mg/ml; 1.6 MDa) at 25°C. Their experimental window encompassed strain amplitudes ranging from 1–300% at frequencies ranging from 1–8 rad/s. A qualitative analysis of the nonlinear viscoelastic response (using Bowditch–Lissajous curves) revealed that HA solutions are predominantly viscous within the tested ranges but retained a small elastic response even at high frequencies and strain amplitudes. Further analysis of LAOS frequency spectra showed that HA transitions from a linear to nonlinear behavior at low strain amplitudes, making nonlinear effects of SF predominant throughout most of the knee motion. Interestingly, the authors used a physical processes methodology to examine the time-dependent loss and storage moduli and found that their magnitude increases with frequency and their variation during the cycle increases with strain amplitude (Fig. 3(A)). Furthermore, they noticed that peaks in elastic moduli aligned with peaks in imposed strain acceleration (max imposed strain) during the oscillation while peaks in the loss moduli aligned with the imposed strain rate during the oscillation (Fig. 3(B)). The authors concluded that during peak strain/acceleration (where
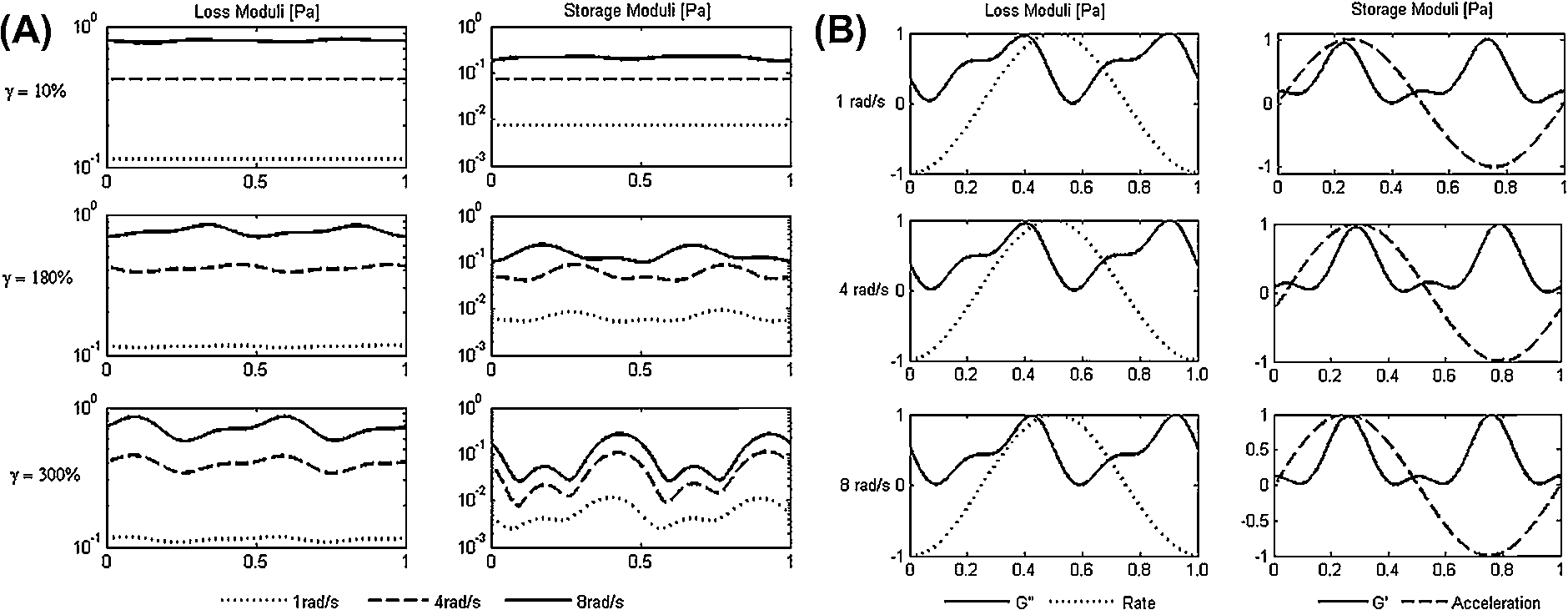
Nonlinear rheology of HA solutions in PBS. (A) Instantaneous time-dependent dynamic moduli. Loss moduli
Early studies of SF suggested that the protein content of SF contributes to its bulk viscosity and elasticity, especially in the low shear rate and frequency regions. Tirtaatmadja et al. [6] showed the direct effect of protein content in SF rheology by first separating the HA and protein content from pooled bovine SF and then examining their individual viscoelastic properties. The authors found that HA solutions exhibited a significant drop in both their viscosity and the storage modulus (
Serum proteins
Macromolecular interactions between HA and serum proteins have been widely examined due to the abundance of the latter in SF and also because early studies struggled to completely purify HA, giving rise to the assumption that these formed a stable HA-protein complex [116,117]. Early studies by Laurent and Ogston [118] found that HA-albumin mixtures had osmotic pressures greater than the sum of colloid osmotic pressures of the individual HA and albumin solutions at the same concentrations.
To examine the rheological contribution of serum proteins, and also due to the restrictive cost and low availability of SF samples, several authors have examined the rheological response of “model SF”, which are physiologically-relevant analogues of SF composed of HA, albumin, and γ-globulin [11,17,86,87,114]. The steady shear rheology of BSF, pure albumin solutions, and model SF solutions comprised of HA (1.5 MDa), albumin, and γ-globulin in PBS (at 3, 11, and 7 mg/ml concentrations, respectively) has been examined extensively [86,87]. By using a concentric cylinder geometry (outer cup diameter 12.0 mm, inner bob diameter 11.1 mm), the authors found that both albumin and model SF solutions exhibit a strong viscoelasticity at low shear rates (⩽1 s−1); this trend has also been reported in model SF (1.6 MDa HA) using a 40 mm diameter 2° cone-and-plate geometry [17]. In addition to viscoelastic enhancements, Oates et al. [87] also reported a rheopectic behavior (i.e., stress increased with time during steady shear) in albumin, model SF and BSF solutions at very low shear rates (0.05 and 0.08 s−1). Since rheopexy reflects the formation of a macromolecular structure under shear, and since the binding between HA and albumin is highly unlikely to occur at pH > 5 [119,120], the authors attributed this behavior to the aggregation of albumin in the bulk, which they proposed formed a weak polymeric network that entangled with HA filaments.
The apparent rheopexy of BSF and albumin solutions was later re-examined by Castellanos et al. [121] using different concentrations of albumin and using sensitive rotational rheometry to measure the individual contributions of interfacial protein adsorption and bulk protein aggregation at various shear rates. The authors found that the apparent viscosity of albumin solutions significantly increased over time (especially at low shear rates) and that these increases were more pronounced for lower protein concentrations. The authors attributed the presence of an apparent yield stress in the measurements to the age-dependent viscoelasticity of a protein film adsorbing at the air–water interface. The authors concluded that, instead of reflecting a continuous aggregation of albumin in the bulk forming a weak network, the rheopectic nature of albumin solutions reflects the consolidation of an air–water protein film. The study did find evidence for limited albumin aggregation in solution, but its contribution to the bulk viscosity at low shear rates was overshadowed by that of its interfacial viscoelastic contribution.
Increasing evidence supports the notion that the age-dependent, viscoelastic enhancements observed at low shear rates in model SF solutions (and pure albumin solutions) reflects systematic rheological measurement errors. Unlike HA molecules in solution, which do not adsorb at air–water interfaces [122], albumin and other globular proteins actively adsorb at the air–liquid interface of the rheometer and introduce artifacts to bulk rheological measurements [11,121,123–125]. In a comparative rheological study, Zhang et al. [11] used three different geometries on a rotational rheometer to examine the apparent viscosity of model SF and found widely varying results, suggesting that measurement artifacts were being captured at low shear rates. In the same study, the authors used a double wall ring geometry to quantify the interfacial viscoelasticity of HA, albumin, and model SF solutions and found that the interfacial rheological response of model SF is probably a combination of the low shear rate behavior of albumin and the high shear rate behavior of HA. Moreover, the interfacial viscoelasticity of model SF also exhibited elastic properties similar to those of albumin and viscous properties similar to those of HA solutions.
Once interfacial effects are abrogated, studies have been unable to find significant bulk rheological contributions stemming from the addition of serum proteins to HA solutions. Zhang et al. [11] used a combination of sensitive low shear rheometry (double gap Couette cell) and applied the surfactant sodium dodecyl sulfate (SDS) to the surface of model SF in order to displace albumin molecules and thus reduce interfacial viscoelasticity. By doing so, the authors found that the bulk rheology of model SF was dominated solely by HA over a wide range of shear rates, strains, and frequencies (Fig. 4). However, oscillatory experiments still showed slight increases in elasticity in model SF at low strains and frequencies, which can be clearly seen in the phase insets of Fig. 4(B), (C); the authors attributed these to protein interactions or aggregation in the bulk.
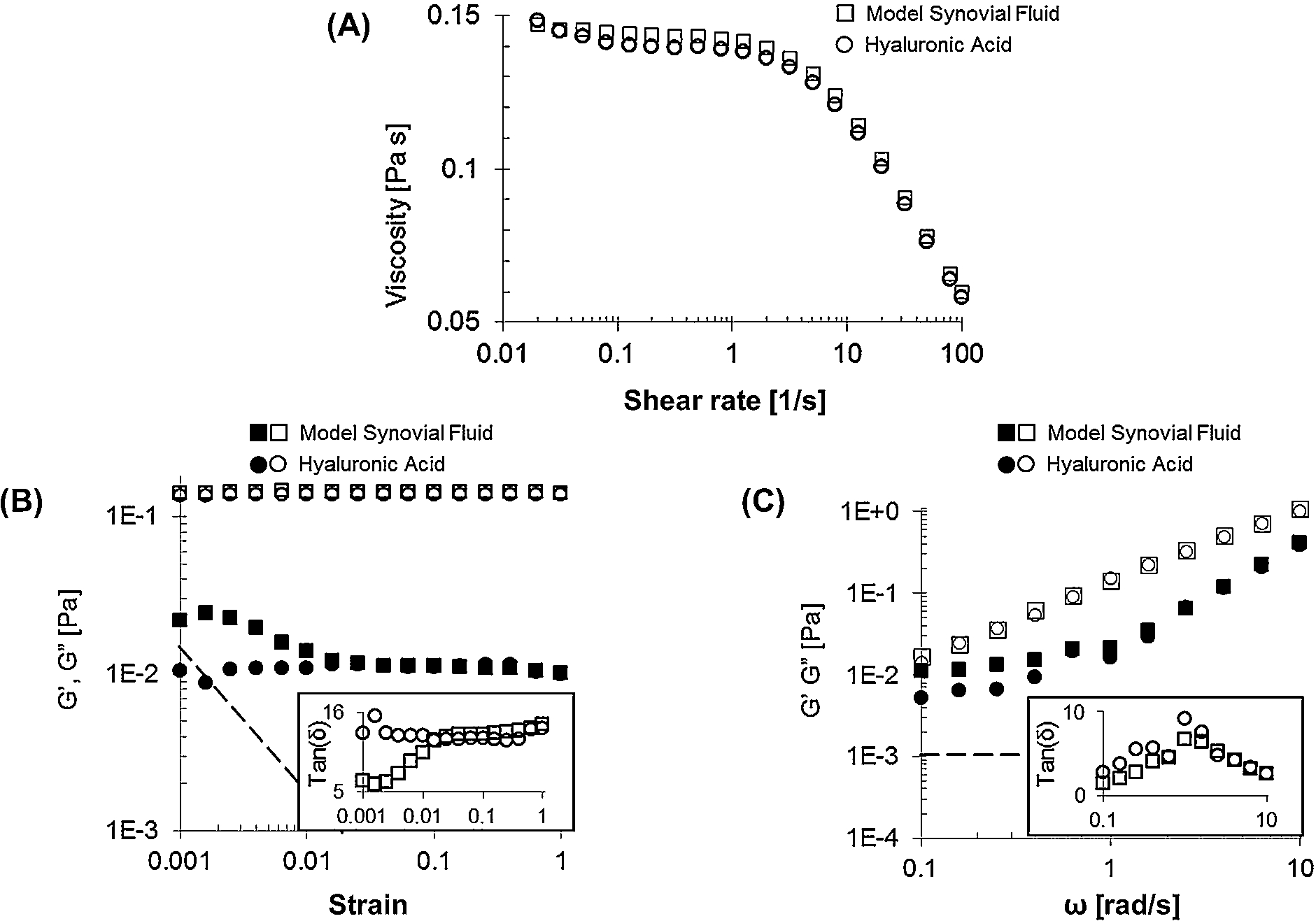
Bulk shear rheological properties of HA and model SF solutions. (A) Viscosity curve, (B) strain amplitude sweep (at 1 rad s−1), and (C) frequency sweep (at a strain of 0.01) for model SF (□) and HA (◯), with
In extensional flows, the rheological influence of serum proteins is still somewhat noticeable. Using their CaBER device, Bingol et al. [10] compared model SF [86,87] to HA solutions (1.7 and 4.6 MDa) at a range of concentrations (0.9 mg/ml–8.8 mg/ml) and observed that the extensional thickening, extensional viscosity, and relaxation times of model SF were only slightly greater than those of pure HA solutions. Tensiometer measurements led them to attribute these effects to a reduction in the surface tension of model SF driven by the serum proteins; this is reasonable since the CaBER device has a large surface area exposed to air that can draw hydrophobic molecules towards the air–water interface and introduce surface tension artifacts into the measurements. Using an optimized interface-free cross-slot device, Haward et al. [17] found no significant differences between the extensional viscosities of HA and model SF (Fig. 5(A), (B)). In a follow-up study, Haward [110] examined diluted porcine SF samples in a microfluidic cross-slot device and also found no conclusive evidence of HA-protein interactions or protein aggregation. However, the author argued that the high strain rate ranges at which measurements were being conducted, combined with the low concentrations of protein, might have dismantled the weak protein network. Finally, the LAOS-strain tests conducted by Zhang and Christopher [114] found no qualitatively large difference between the nonlinear stress output of HA and model SF solutions (Fig. 5(C)). Taken together, the results of all these studies strongly indicate that the functional viscoelastic properties of model SF are mostly conferred by HA molecules in solution.
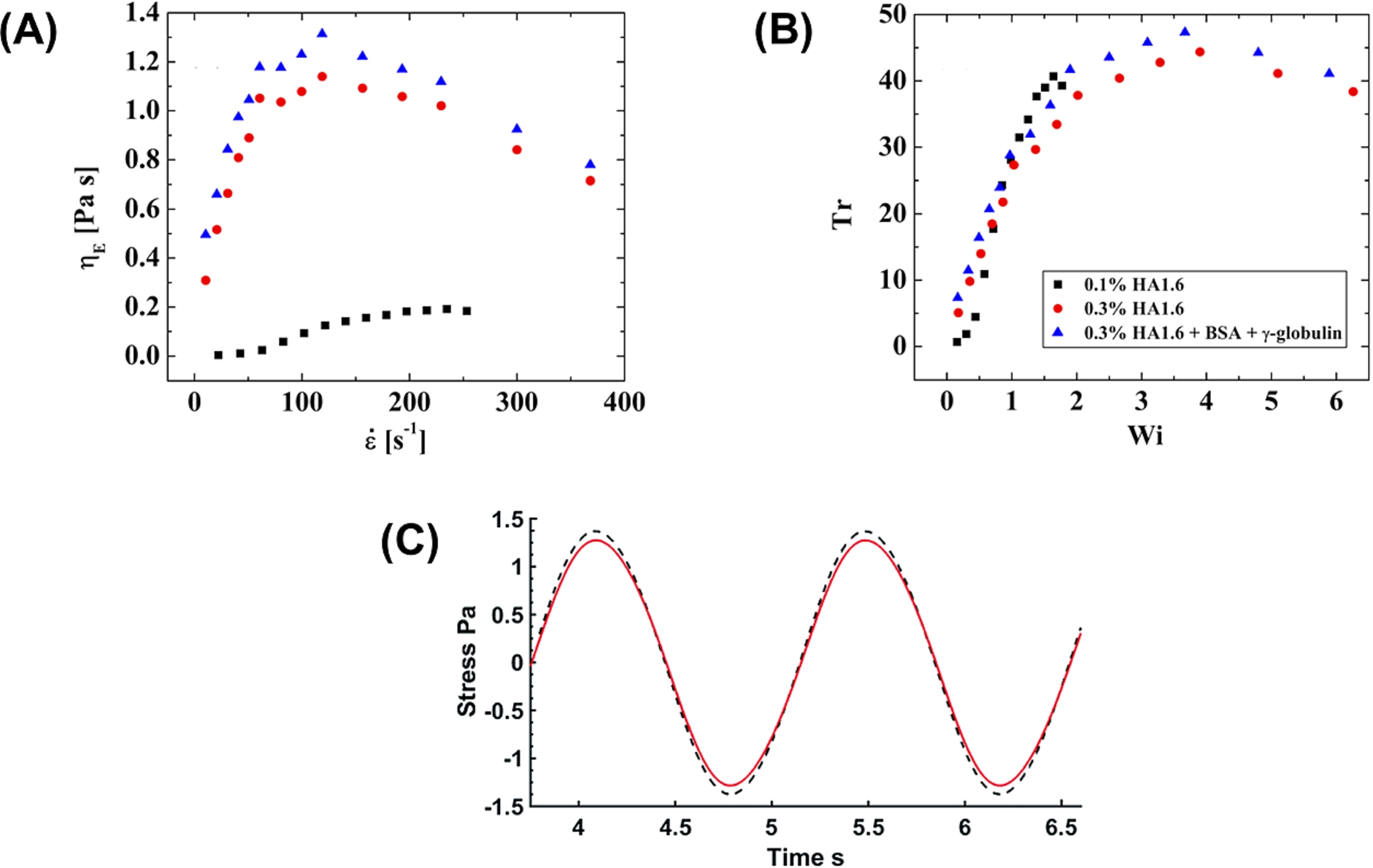
Extensional and nonlinear rheology of HA and model SF solutions. (A) Extensional viscosity (
Aside from tribological evidence of synergism, recent rheological studies suggest that HA-PRG4 interactions could play a significant role in the bulk rheological behavior of SF. Jay et al. [60] observed that purified synovial lubricating factor (PSLF, i.e., PRG4) at a concentration of 50 µg/ml in the presence of 4 mg/ml purified umbilical hyaluronate (UHA-P) resulted in a significant decrease in viscosity whereas the addition of 70 µg/ml of PSLF in 4 mg/ml of normal UHA increased it. This apparently anomalous finding was also exhibited in the interaction between the HA-binding region of a rat chondrosarcoma proteoglycan and UHA. The authors suggested that PSLF probably decreased the axial ratio of UHA-P and thus forced it to assume a more ellipsoidal molecular shape; however, this effect could also be explained by a reduction in the hydrodynamic volume of HA.
In a more recent study, Jay et al. [9] used a combination of bulk shear rheometry and multiple-particle-tracking micro-rheology to examine the viscoelastic properties of various HA, PRG4, HA-PRG4 and SF samples/solutions (see Fig. 6). Steady shear tests using a 40 mm diameter 0° cone-and-plate geometry showed that the zero-shear viscosity of HA solutions decreased with the addition of 0.3 mg/ml of PRG4. Also, both the trypsin-digested and the PRG4-deficient SF (from patients with the camptodactyly-arthropathy-coxa vara-pericarditis syndrome who lack the ability to express functional PRG4) exhibited an increased zero-shear viscosity, a shifted shear-thinning region, and a higher network diffusivity when compared to normal BSF. These results suggest that PRG4 molecules force HA to adopt a more coiled and less extended conformation. The authors concluded that PRG4 may play a crucial role in the structural organization of the SF network by increasing the elastic absorption and thus facilitating the slow dissipation of strain energy during loading cycles. PRG4 may increase the HA network elasticity by organizing and mediating the entanglements, which results in longer relaxation times and reduced viscosities. The authors suggested that, by controlling the diffusivity of the network, PRG4 could play a biological role by controlling the transit of biomolecules through the cartilage matrix [9].
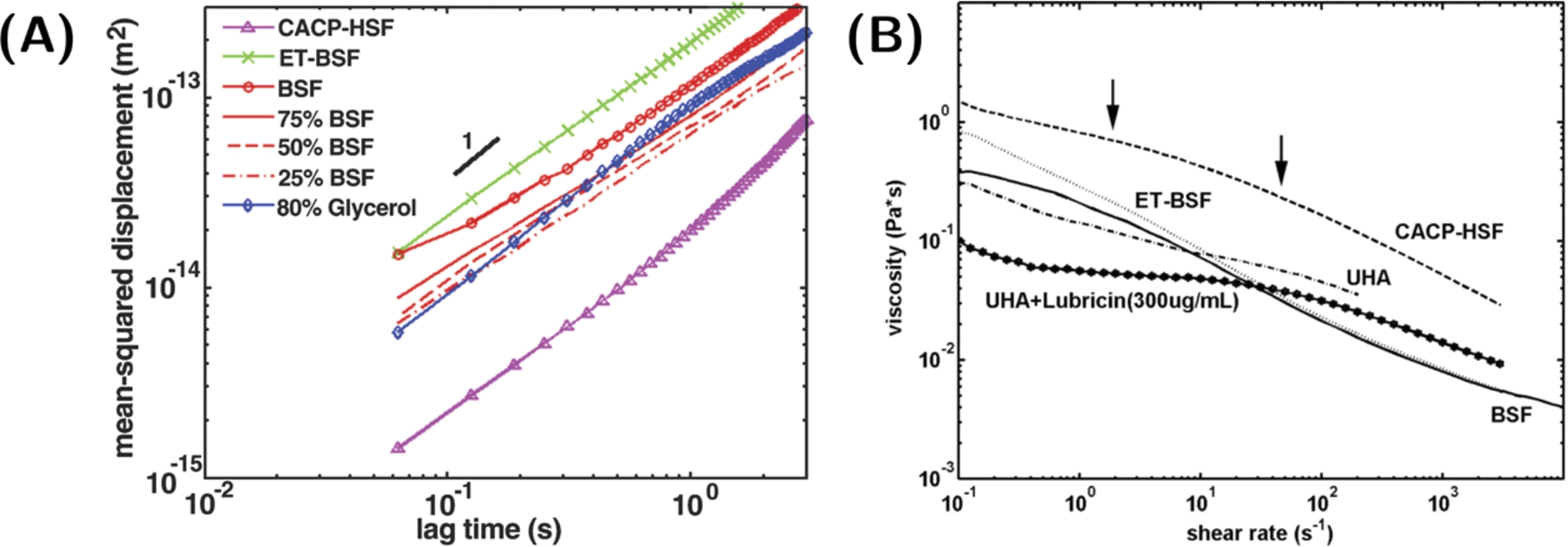
Viscoelasticity of diverse SF samples and HA-PRG4 mixtures. (A) Diffusive behavior of 200-nm particles in bovine SF (BSF), glycerol, enzyme-digested BSF (ET-BSF), PRG4-deficient human SF (CACP-HSF), and BSF diluted with 80% glycerol. Newtonian fluids exhibit a linear relationship between the mean-squared displacement and the lag time (slope of unity) whereas non-Newtonian fluids exhibit a nonlinear relationship (slope lower than unity). (B) Shear rate dependent viscosity of CACP-HSF, BSF, ET-BSF, umbilical cord HA (UHA), and UHA-PRG4 mixture. The onset of power-law behavior (between arrows) is indicated. Shear measurements were made using a 40 mm diameter 0° cone-and-plate geometry. Reproduced from [9] © 2007 The National Academy of Sciences.
In a recent study, Ludwig et al. [24] compared the shear viscosities of bovine PRG4 (with and without disulfide bonds) and recombinant human PRG4 (rhPRG4, [126]) solutions, as well as their interactions with HA. The authors demonstrated that, unlike rhPRG4, PRG4 can self-associate and increase both the bulk viscosity and shear-thinning properties of PRG4 solutions in a manner dependent upon concentration and the presence of disulfide bonds. The authors suggested that these rheological differences might stem from other proteins surviving after PRG4 purification or, more likely, from differences in the O-linked glycosylations, which are known to affect biological boundary lubrication [69]. Furthermore, when either PRG4 and rhPRG4 were combined with HA at 3.3 mg/ml, there was a decrease in both the viscosity and shear-thinning behavior of the solutions; the authors suggested that a PRG4-induced complexation with HA could organize the macromolecular network of the solution and therefore reduce the hydrodynamic volume of the HA molecules.
At lower HA concentrations however (0.3 and 1.0 mg/ml), the authors found that the addition of PRG4 increased the viscosity and shear-thinning strength, which suggests a minimum PRG4 concentration required before noticeable interactions with HA occur. Furthermore, these rheological effects were not seen in HA solutions containing PRG4 with disrupted disulfide bonds. The authors suggested that the interactions between PRG4 and HA are mediated by intra and inter-molecular disulfide bonds and, therefore, the tertiary and quaternary protein structures of PRG4 are of relevance in the rheological behavior of the macromolecular network of SF. The authors proposed that low concentrations of HA trigger PRG4 to self-associate via cooperative assemblage that increases overall solution viscosity; conversely, high concentrations of HA in HA-rhPRG4 solutions decrease the hydrodynamic volume of HA molecules and thus results in decreased overall viscosity. The authors posited that PRG4 may have the biological role of counteracting decreases in HA concentration in pathological SF by complexing and thus compensating for the loss of viscosity.
As with serum proteins, there is a possibility that SF-specific proteins like PRG4 (or other SF-specific glycosaminoglycans) readily adsorb onto the air–water interface and/or onto the surfaces of the rheometers themselves, thus introducing rheological artifacts into the measurements. Given that SF retains a third of the plasma protein content, its surface adsorption kinetics may resemble those of blood and will also be dictated by competitive adsorption. It could be the case that the adsorption of surface-active proteins like PRG4 onto the surface of rheometers could introduce time-dependent artifacts into the measurements, especially if the bound PRG4 tethers suspended HA molecules; this would both reduce the concentration of HA in the SF matrix and, given HA’s water-imbibing ability, it could sequester a liquid phase at the surfaces of the rheometer. Any adsorption–desorption differences between blood and SF would depend on several factors, including (but not limited to) the surface hydrophobicity of the rheometer, the mobility and binding affinity of lubricin, and the time allowed for the consolidation of the protein film. Quantitative surface characterization studies like quartz crystal microbalance or ellipsometry should be used to evaluate the adsorption kinetics of interfacially active proteins and biomolecules in SF.
In this review, we have presented an overview of SF composition, the chemical structure of its main components, and how inter- and intra-molecular interactions in the SF matrix influence its bulk rheological properties. Aside from conventional steady and dynamic shear tests, this review has illustrated the value of examining SF and HA-protein solutions under extensional and nonlinear flows; a full characterization of these is needed before the macromolecular interactions taking place in vivo can be elucidated. Due to the small viscoelastic contributions directly provided by serum proteins, future studies will most likely focus on examining HA-PRG4 solutions and how PRG4’s degree of multimerization and glycosylation directly affect its extent and mode of interaction with HA. A full characterization of the macromolecular interactions between SF constituents and how these influence the rheological properties of SF is vital for the development and optimization of novel SF substitutes that can treat diverse arthropathies, serve as viscosupplements or periprosthetic fluids, or be used as rheological analogues for testing the wear and damage of orthopedic implants.
Footnotes
Acknowledgement
We thank Dr. Mary K. Cowman (Biomatrix Research Center, Department of Chemical and Biomolecular Engineering, Tandon School of Engineering, New York University, New York, NY) for useful discussion in preparing this review.