
Abstract
BACKGROUND:
Arterial function is regulated by estrogen, but no consistent pattern of arterial mechanical remodeling in response to depleted estrogen levels is available.
OBJECTIVE:
To examine long-term effects of ovariectomy (OVX) on the mechanical properties, morphology, and histological structure of the carotid artery in middle-aged rats and a potentially protective effect of Sideritis euboea extract (SID), commonly consumed as “mountain tea”.
METHODS:
10-month-old female Wistar rats were allocated into control (sham-operated), OVX,
RESULTS:
Remodeling in OVX rats was characterized by a decreased in situ axial extension ratio, along with increased opening angle, thickness, and area of the vessel wall and of its medial layer, but unchanged lumen diameter. Compositional changes involved increased elastin/collagen densities. Characterization by the “four-fiber” microstructure-motivated model revealed similar in situ biaxial response of carotid arteries in OVX and control rats.
CONCLUSIONS:
Carotid artery remodeling in OVX rats was largely consistent with hypertensive remodeling, despite the minor arterial pressure changes found, and was not altered by administration of SID, despite previous evidence of its osteo-protective effect.
Introduction
The lower incidence of cardiovascular disease in premenopausal women as compared to age-matched men and the sharp rise after menopause, as well as the beneficial effects of estrogen therapy in postmenopausal women indicate a protective role of estrogen on the cardiovascular system [1]. Several mechanisms have been put forth as the basis for these protective effects. Estrogen alters serum lipid concentrations in favor of high-density lipoproteins (systemic actions), while, as part of its direct actions on blood vessels, enables vasodilation (short-term) and inhibits the blood vessel response to injury and the development of atherosclerosis (long-term effects) [2–4].
Surprisingly, although it is generally thought that depleted estrogen levels in postmenopausal women are associated with greater age-related arterial stiffening compared to men and that increased arterial stiffness is an independent risk factor for cardiovascular disease, only few clinical studies have addressed the mechanical remodeling of arteries after menopause [5–7]. As regards the experimental (animal) studies, changes in arterial wall properties have been documented in ovariectomized rodents, serving as common models of estrogen deficiency, but there is clearly a lack of unanimity vis-à-vis the exact nature of the changes in wall composition, dimensions, and mechanical properties [8–14]. Even more importantly, the young age of the ovariectomized animals in all of the previous studies does not reflect the age of women at menopause and the possibility of distinct modes of arterial remodeling or even of impaired remodeling capability in middle-aged animals cannot be dismissed, given the known changes with aging in arterial phenotype and hemodynamics, i.e. the blood pressure and flow conditions that also regulate arterial remodeling.
Accordingly, it was the primary objective of the experiments described herein to address prior inconsistencies and to define changes in the mechanical properties of the carotid arteries associated with surgical ovariectomy in the middle-aged female rat. Our attention was focused on the carotid arteries, because their mechanical properties, along with those of the other central arteries, modulate cardiovascular homeostasis through the buffering of the pulsatile output of the heart and its translation into a steady flow in the periphery, constituting a prognostic factor in cardiovascular disease [15]. Our secondary objective was to examine the effect of Sideritis euboea (SID; Mediterranean mountain tea) extract administration to determine whether it produces a vasculo- and/or cardio-protective effect. Previous publications have demonstrated that the extract from this plant has bone mineralizing properties in vitro [16], as well as bone density and strength protective effects in vivo in the mature ovariectomized rat model that closely mimics postmenopausal osteoporosis in women [17]. This and several other plant extracts appear to be potential alternative options to pharmaceutical prevention of postmenopausal osteoporosis in women [18,19]. Third, detailed morphological and histological studies were conducted to measure the arterial geometrical and compositional changes, and to correlate them with the changes in the mechanical properties, issues that have not been investigated in this animal model to date.
Materials and methods
Animal groups, induction of ovariectomy, and administration of extracts
The study protocol was approved and licensed (permit no. K 5181/2006) by the General Veterinary Services and the animals’ care and use was according to Greek legislation (Presidential Decree 160/1991) in compliance to the Directive 86/609/EEC. Thirty-two mature (10-month-old) healthy female Wistar rats were obtained from the registered breeding facility of the Biomedical Research Foundation of the Academy of Athens (Athens, Greece). The rats were housed four to a cage under standard laboratory conditions, i.e. 19–22°C temperature, 55–65% relative humidity, 15 air changes/hour, and a light/dark cycle of 12/12 hours. The rats were allocated to four groups: control (
MALT was administered to the
Euthanasia and tissue collection
At the end of the 6 month postoperative administration period and following euthanasia by an overdose of anesthesia, an autopsy was conducted to inspect the rats for malignancies or other pathological findings, to substantiate the success of OVX, and for the resection of uteri and common carotid arteries. The success of OVX was substantiated by failure to detect ovarian tissue and by observing notable atrophy of the uterine horns. The uteri were excised and weighed. The left common carotid artery was exposed via a midline neck incision and two superficial surgical knots were tied immediately distal to the aortic arch and immediately proximal to the carotid bifurcation, taking care not to exert excessive loads. The axial in situ length of the left common carotid artery was determined by measuring the distances between the knots and loose connective tissues were cautiously trimmed. During measurements and excision, the head of the rats was held in a natural position, namely almost aligned with the body. The carotid artery including the knots was then excised, put in a cuvette with Krebs–Ringer solution at 37°C to stabilize for
Histology
Paraffin-embedded transverse sections of the carotid artery were stained with hematoxylin-eosin, Sirius red, and orcein, using standard techniques and reagents. All measurements were carried out using image-analysis software (Image-Pro Plus v4.5; Media Cybernetics Inc, Silver Spring, MD, USA) on images captured by a digital camera (Altra20; Soft Imaging System, Munster, Germany) connected to a light microscope (Olympus CX31; Olympus, Tokyo, Japan), as has been previously detailed [20–22]. Vessel dimensions were measured on hematoxylin-eosin stained sections under suitable magnification, specifically
Mechanical studies
Geometrical measurements
Prior to examining the inflation/extension characteristics of the vessel segments, thin (
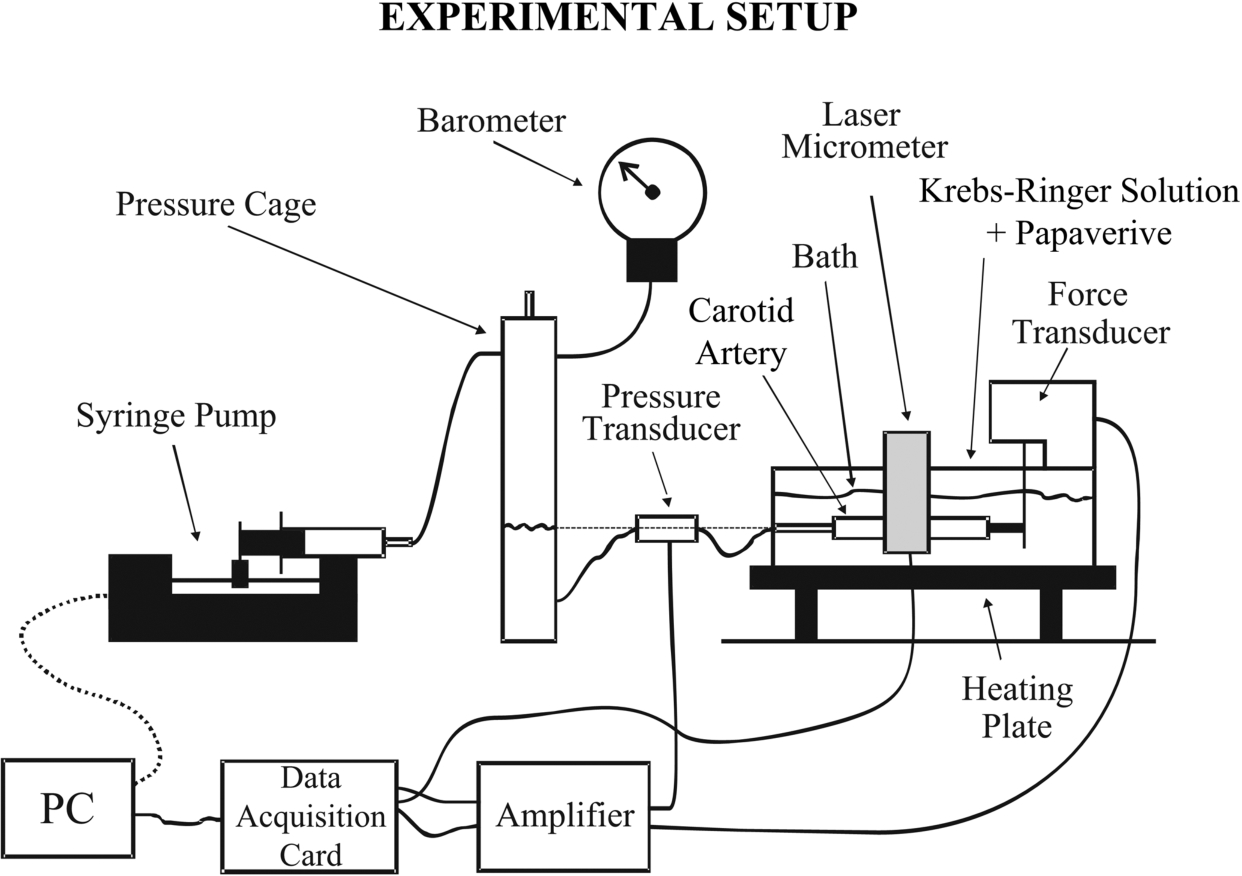
Schematic diagram of the experimental apparatus used for inflation/extension testing.
The residual carotid artery segments were mounted at their in situ length in the experimental apparatus, previously described in detail [20–22] and shown schematically in Fig. 1; with the implicit assumption that the axial in situ extension ratios of the segments used for mechanical testing were identical to those of the entire carotids harvested. The segments were immersed in the temperature-controlled bath of the apparatus and horizontally mounted on two adaptors, consisting of 20-gauge needles plugged on stainless steel cannulae. One of those was kept fixed whereas the second was connected to an isometric force transducer (Fort 100; World Precision Instruments, Hertfordshire, UK), measuring the axial force exerted to the segments. This adapter was suspended from a manifold on a screw-operated slide mechanism, used to regulate the length of vessels and their position in the bath with a micrometer (Tesa Technology, Renens, Switzerland). Length measurement was performed as the separation distance between the cannulae, justified by the large aspect ratio of the arterial segments (their average no-load length,
By use of a syringe pump (model sp100i2; World Precision Instruments), the arterial segments were submitted to a series of five inflation-deflation loops from 0 to 200 mmHg at a rate of 0.5 mmHg/s, serving to eliminate viscoelasticity and produce stable diameter-pressure-force loops. The inflating part of the last loop was used for the subsequent data analysis.
Material characterization
As is common in most material characterization studies (reference is made to the standard cardiovascular biomechanics textbook by Humphrey [23]), the carotid artery was considered to be a nonlinear, residually stressed, pseudoelastic, homogeneous, and orthotropic cylinder subjected to large isovolumetric deformations by lumen pressure and axial force. In the general 3D setting, circumferential and axial extension ratios
Inner radius
Compliance C and distensibility D to be used as structural measures were calculated from lumen area
Qualitative description of the stress-strain curves, together with distensibility and/or compliance indices have been used by most previous investigators to assess the effects of OVX on the in vitro mechanical properties of blood vessels [8–10,13]. These indices were also used herein (viz. Eq. (4)) for comparison purposes but they are limited in that they do not permit rigorous characterization of the multiaxial material properties, unlike current microstructure-motivated models that allow proper characterization with respect to tissue microstructure. As such, we used on our inflation/extension data the neo-Hookean and four-fiber family model, proposed by Baek et al. [24]. Note the far-reaching applicability of the four-fiber family model in that it has been tested on rabbit basilar arteries [24], on mouse carotid arteries from two common models of muscular dystrophy and wild-type control [25], and for biomechanical phenotyping of carotid arteries from various genetic, pharmacological, and surgical mouse models [26]. It has also been used by our group for material characterization of various soft tissue types, e.g. porcine and rabbit carotid artery [27], porcine jugular vein [28], rabbit esophageal [29], rat large intestinal [30], and rabbit ureteral tissue [31]:
Material parameters were calculated by fitting Eq. (6) to the pressure-force-radius testing data excluding those associated with negative forces, using the Nelder–Mead algorithm in MicroCal Origin (v8.5; OriginLab Corp., Northampton, MA, USA), as in our prior studies [27–31]. The parameters were restrained by inequalities of thermodynamic origin, ensuring that the material model was convex and the parameter values physically realistic. These inequalities prescribed a zero lower limit for all parameters and an upper limit of
Statistical methods
Results are presented as
One specimen from the control (sample #2), OVX (sample #6), and
Results
Animals’ body and uterine weights, and hemodynamic measurements
There were no adverse effects by the administration of SID and/or MALT, as witnessed by monitoring body weight, fluid, and food consumption, and no pathological findings/malignancies were observed at autopsy. Water and extract consumption did not vary among groups. Body weight was similar in the four groups at baseline, but the rats of OVX groups weighed much more compared to control at euthanasia, without differences among the OVX,
Body weight, uterine weight, and hemodynamic recordings and their statistical comparison in control, OVX,
, and
rats
Body weight, uterine weight, and hemodynamic recordings and their statistical comparison in control, OVX,
Values are
Figures 2 and 3 show that the overall carotid artery wall structure was preserved six months after OVX. In Fig. 2 the morphometric results obtained from computerized histology are also shown. A consistent finding was the increase in all OVX groups of the cross-sectional area occupied by the media, but not of that occupied by the adventitia, leading to an increase in total wall cross-sectional area. No differences were detected in the thickness and area of the various layers and the entire wall among the OVX groups, as well as in the inner and outer diameters of the carotid artery between all OVX groups and control.
As remarked in Fig. 3 summarizing the composition parameters measured, the relative area density of collagen in tunica media and adventitia, quantified in Sirius-red sections, was greater in all OVX groups than control. So was elastin area density in the media, quantified in orcein sections, while cell area density in the media and adventitia was decreased. As a result, elastin and collagen area densities in the entire wall were greater in OVX arteries than control, whereas cell area density was decreased. Unchanged was the number of lamellar units in the media and the number of nuclear profiles per unit surface area in all layers and the entire wall (data not shown). These data imply that arterial growth in response to OVX was characterized by collagen and elastin buildup.
Carotid artery mechanical properties
Geometrical parameters
Table 2 lists the morphological parameters measured from the digitized photographs of the carotid artery segments in the no-load and zero-stress state. In the no-load state, the inner diameter did not vary between control and OVX,
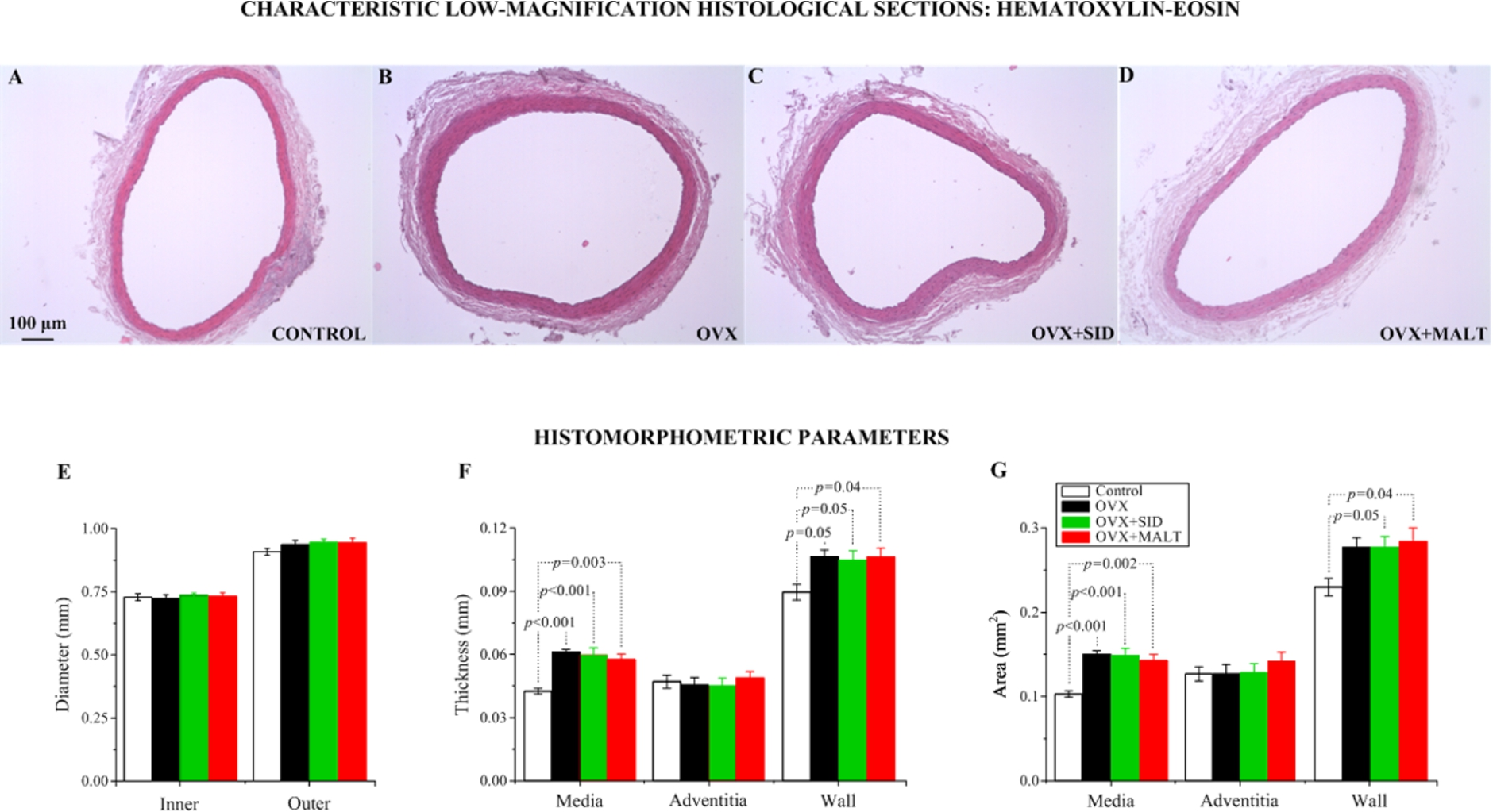
Characteristic low-magnification histological sections of the common carotid artery stained with hematoxylin-eosin from (A) a control, (B) an OVX, (C) an

High-magnification histological sections of the common carotid artery, adjacent to those of Fig. 1, stained with orcein for elastin ((A)–(D)) and sirius-red for collagen ((E)–(H)) differentiation from ((A), (E)) a control, an OVX ((B), (F)), an
The opening angle and residual extension ratio results at the intimal and adventitial surfaces of the carotid artery are depicted in Table 2. Opening angle was significantly higher in
Figures 4 and 5 illustrate the findings from inflation/extension testing on the common carotid arteries at the in situ extension ratio and under passive smooth muscle that permit direct comparison among groups at their individual in situ condition. All arteries exhibited a bi-phasic deformational response (Figs 4(A) and (C)), with a low-pressure phase (0–80 mmHg) referring to a great variation of outer radius and wall thickness, and a physiologic and supra-physiologic phase referring to radius and thickness locking. Note the quite similar curve shape of OVX and control arteries. Pressure inflation in the former led to a somewhat greater radius enlargement at all levels of pressure that was associated with an equally compliant and distensible wall than control, however; Figs 5(A) and (B). Quite similar were also the cumulative circumferential and axial stress-circumferential extension ratio data between the OVX and control carotids shown in Figs 5(C) and (D), noting that the data referred to a significantly smaller axial in situ extension ratio in OVX compared to control rats (Table 1). Note also the striking similarity in the curves of OVX,
Carotid artery material characterization
Pressure-radius-force data and curve-fits are shown in Fig. 6 for all the carotid arteries tested. The neo-Hookean and four-fiber family model provided a realistic characterization of the inflation-extension response (
Geometrical parameters in the no-load and zero-stress state of the common carotid artery and their statistical comparison in control, OVX,
, and
rats
Geometrical parameters in the no-load and zero-stress state of the common carotid artery and their statistical comparison in control, OVX,
Values are

(A) Outer radius, (B) axial force, and (C) wall thickness vs. lumen pressure data of control, OVX,
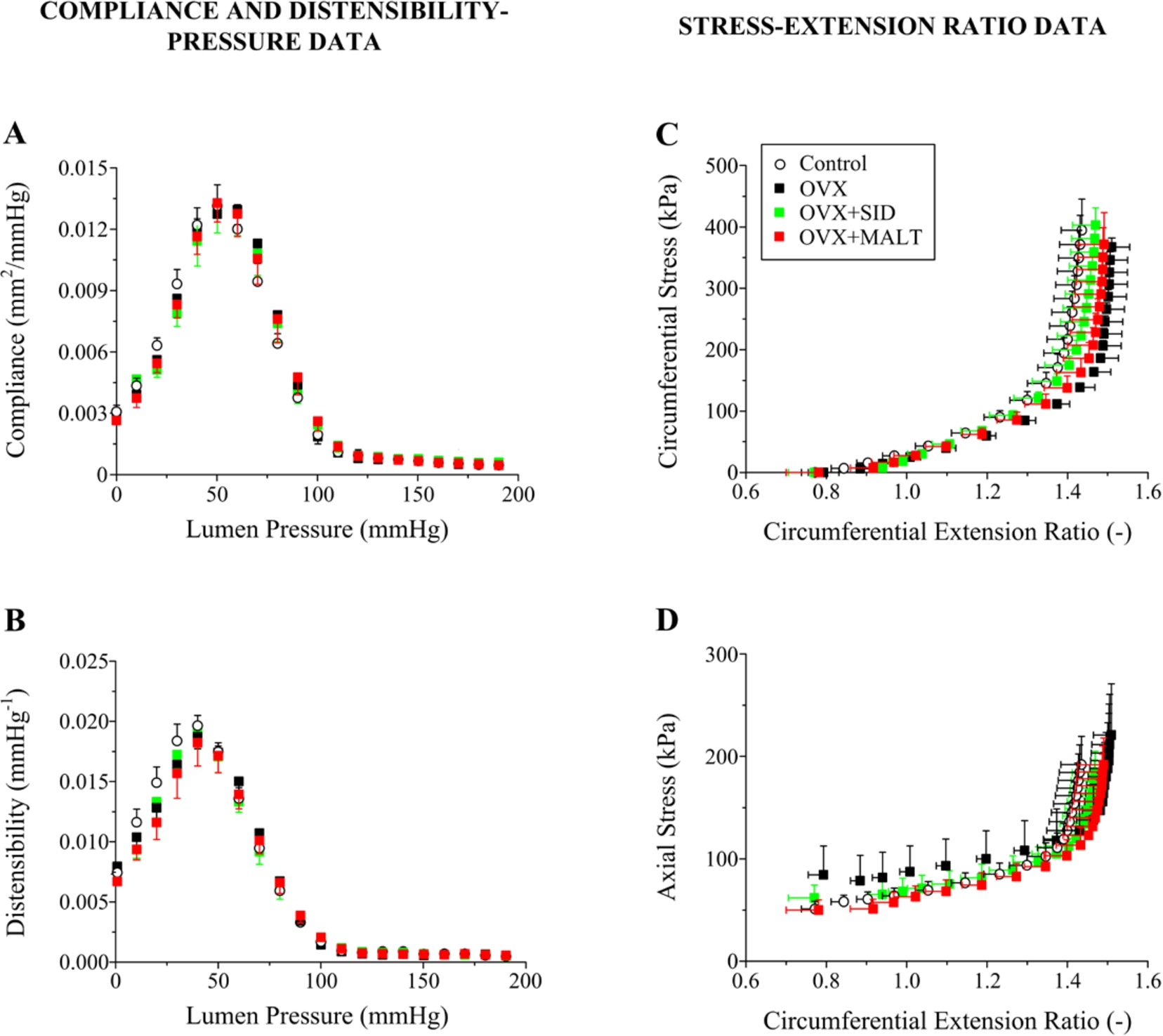
(A) Compliance and (B) distensibility vs. lumen pressure data, and (C) circumferential and (D) axial stress vs. circumferential extension ratio data of control, OVX,
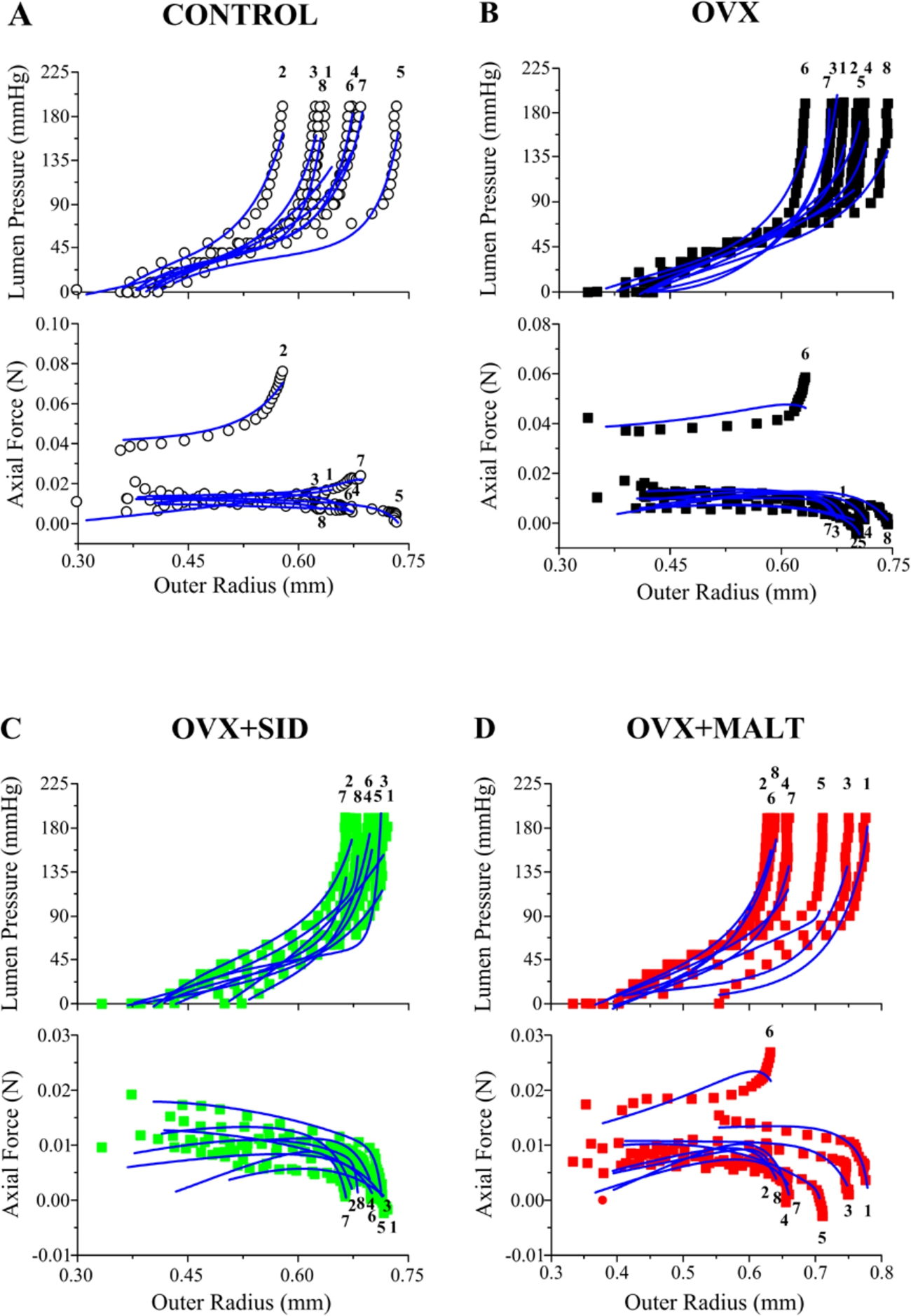
Experimental lumen pressure (upper panel) and axial force (lower panel) data as a function of outer radius for all carotid arteries tested and fits by the neo-Hookean and four-fiber family model for (A) the control, (B) OVX, (C)
Animal models and general considerations
A number of studies [8–14] have compared the mechanical behavior of arteries from sham-operated and OVX animals before and after hormone replacement therapy, but relatively young immature animals were only examined, even though the mechanical properties of arteries from common laboratory animals are modified with aging, as do the hemodynamic conditions, and most importantly the capacity of blood vessels to remodel may be impaired by age [32–34]. To overcome this shortcoming, OVX was performed in the present study to middle-aged rats, thus ascertaining whether the arterial remodeling is age-related. Particularly, we selected mature rats at the age of 10 months, since this is the age that they achieve peak bone mass and is considered most appropriate for modeling postmenopausal osteoporosis in women [35].
Inherent differences among the various studies may be responsible for the inconsistencies in the reported remodeling for young animals, such as lumen narrowing in response to depleted estrogen levels in some cases [9,10] and increased wall thickness in other cases [12], and stiffening [12,14] unlike the invariant [8–10,13] and decreased stiffness [11] disclosed by other authors. The reasons behind these discrepancies are uncertain but may be ascribed to differences in the duration of OVX (few weeks [8,10–12] vs. several months [9,13]) or in the blood vessels tested (large elastic [11–13] vs. small resistance arteries [8–10]). Other likely explanations relate to the methods employed to determine the mechanical properties (in vitro [8–10,13] vs. in vivo [11,12,14]) and dimensions (no-load [8] vs. loaded state [9–12]). Despite the rather incomplete data, Section 4.2–4.4 lay emphasis on interesting analogies between the present and past studies, while providing a comprehensive assessment of the mechanical, morphological, and structural aspects of remodeling.
Parameters and fit quality of the four-fiber family model for the common carotid artery and their statistical comparison in control, OVX,
, and
rats
Parameters and fit quality of the four-fiber family model for the common carotid artery and their statistical comparison in control, OVX,
Values are
The outer radius-lumen pressure curves of the carotid arteries from all three OVX groups were displaced to greater radii (Fig. 4(A)), caused by their greater wall thickness (Fig. 4(C)) and inner radius. This finding may be expected based on the earliest mechanics-based study by Cox and Fischer [36], who conducted pressure-radius measurements under inflation at in vivo length in the presence of smooth muscle inhibitors. The curves for carotid arteries treated with estradiol were shifted to smaller radii, consistent with the effect of estrogen in decelerating the weight gain of rats (Table 1), and those for testosterone-treated arteries to greater radii, with qualitatively similar results in castrated and non-castrated male rats. Three-week administration of estradiol decreased the incremental elastic modulus, a measure of material stiffness, whereas administration of testosterone increased it, albeit to a lesser extent, through the opposite effects of these steroids on the elastin/collagen accumulation in the arterial wall that was demonstrated in their companion study [37].
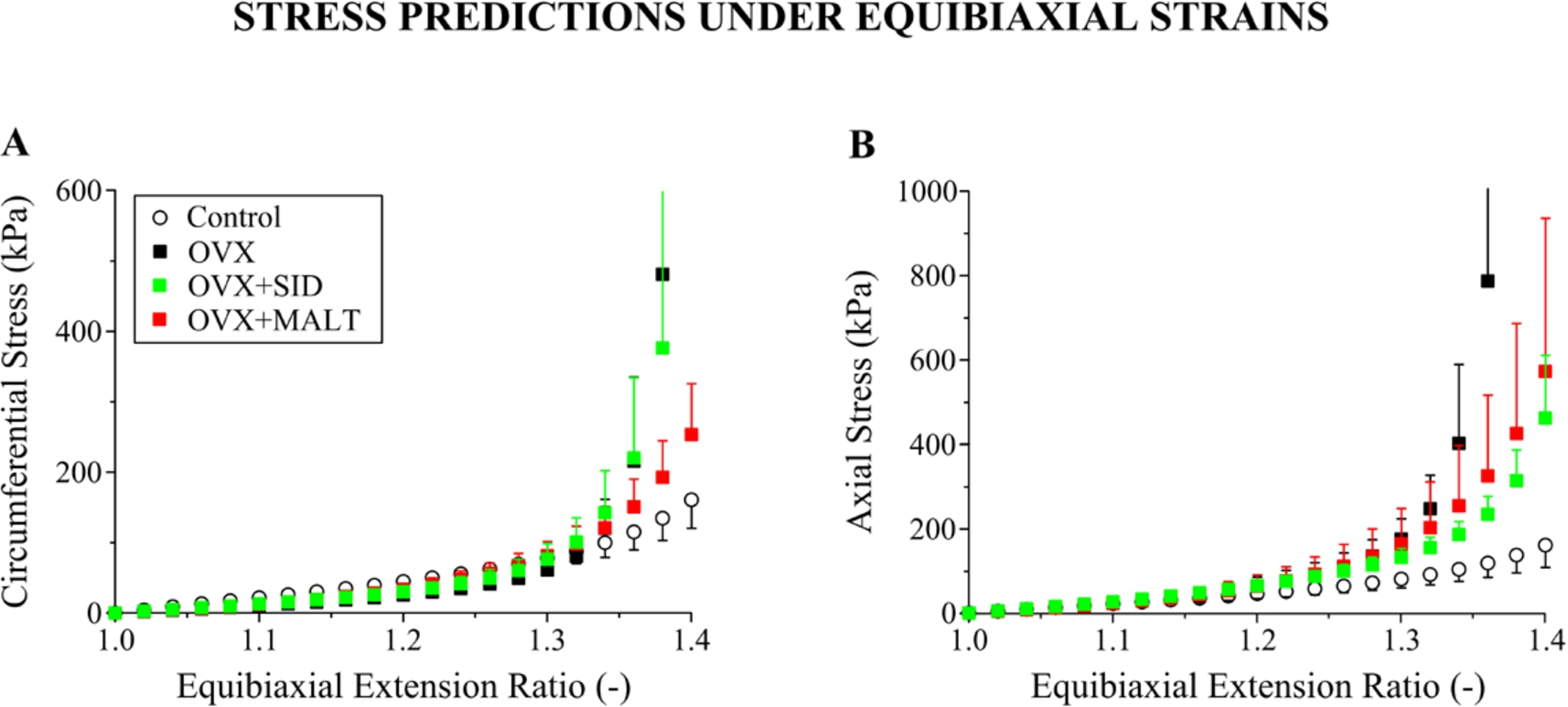
Carotid artery stiffness comparisons and anisotropy assessment via the prediction of (A) circumferential and (B) axial stresses at equibiaxial extension ratio states by the neo-Hookean and four-fiber family model for the control, OVX,
Acs et al. [9 ,10] conducted in vitro tests of saphenous arteries from OVX and control rats at in vivo length and potential modification in vasoactivity was appropriately considered. Contrary to our findings, 12-week pharmacological OVX shifted the passive pressure-radius curves to smaller inner radii that were restored by hormone replacement therapy, with unchanged vessel wall thickness. No significant changes in distensibility (structural) and incremental elastic modulus (material index) were found, in certain analogy to the invariant distensibility as well as circumferential and axial stress vs. circumferential extension ratio curves for the OVX and control rats reported herein (Fig. 5). More recently, Guo et al. [12] reported data from in vivo pressure-radius experiments on mouse aorta, and carotid and femoral arteries. Much like the present data, they disclosed that surgical OVX for 4 weeks increased wall thickness but not lumen radius and, although it did not affect the axial stiffness as we found, it substantially increased the circumferential stiffness. Unfortunately, axial force was not measured in any of the abovementioned studies, so that the pressure-radius data alone constitute in essence uniaxial data that do not suffice for multiaxial material characterization. Other important information that is missing regards the in vivo axial prestretches of vessels, and their zero-stress state was not examined except in [12] but no opening angle and/or residual strain data were presented.
Park et al. [13] reported that estrogen deficiency did not have an effect on aortic stiffness of rats at 12 weeks after surgical OVX, basing their conclusions on ring tests. Their finding of unchanged elastic modulus, and ultimate tensile stress and stretch were corroborated by their biochemical data, that is the absence of calcium deposition and elastin content change; the latter finding being at variance with our light densitometry data (Fig. 3). Note, however, that ring tests are less preferred for both experimental and theoretical reasons, as they provide uniaxial data with decoupled circumferential and axial loads as experienced physiologically, the unloaded state of tissue is considered as its zero-stress state, and bending during the initial testing phases is ignored. Similar reservations, therefore, hold for the ring studies by Hansen et al. [8] on rabbit cerebral and coronary arteries that disclosed unchanged elastic modulus at 6 weeks after surgical OVX, as well as unchanged collagen content and morphologic features, i.e. lumen diameter, wall thickness, and layer thicknesses.
Other parameters being alike in two available pulse wave velocity studies, i.e. unchanged wall thickness and lumen diameter (unlike our results in Table 2), the significant aortic pressure rise disclosed in [14] after 2 months may be responsible for the increased pulse wave velocity in OVX rats. Even adjusting for pressure, this finding contrasts with [11] showing a reduced pulse wave velocity and elastic modulus after a 3-week OVX period, despite the unchanged pressure. However, inferences of aortic stiffness from pulse wave velocity measurements are not straightforward given the complexity and number of their determinants, including wall dimensions, passive wall elements, smooth muscle tone, and the level of distending pressure.
The mechanical remodeling of the carotid artery in OVX rats was mathematically assessed by characterizing our inflation-extension data using a microstructure-motivated material model (Eq. (6)). As such, we employed the neo-Hookean and four-fiber family model that is extensively used in the literature and permitted realistic data characterization for both OVX and control arteries (Fig. 6). Although non-significant due to the wide scatter, the parameter values optimized exhibited marked differences between OVX and control arteries, i.e. a decrease in the neo-Hookean function parameter (associated with elastin and other non-collagenous elements) and an increase in several four-fiber family parameters (associated with collagen fibers; Table 3). At equibiaxial stretch states, the model predicted a significantly stiffer carotid artery wall in all OVX groups compared to control (Fig. 7), justified by the increased medial and adventitial collagen contents found that are circumferentially- and axially-aligned, respectively [27]. Note, however, that the material characterization results would have been more robust had the inflation testing been performed at several and not only at the in situ axial extension ratio that was unfortunately performed in the current experiments. Comprehensive testing protocols, including a large variety of loading scenarios, are necessary for accurate parameter identification of nonlinear material models. Consideration of pressure-radius-force data at additional axial extensions and of axial force-length test data at different pressures, as done in the early study for the rat carotid artery by Weizsäcker et al. [38], and more recently for the mouse carotid artery by Gleason et al. [25] and Bersi et al. [26], would have considerably strengthened the descriptive and predictive capabilities of the chosen material model.
Humphrey et al. [39] (see also [26]) suggested the determination of the in situ axial extension ratio from the force-length relationship, where various isobaric axial force-length curves intersect, or alternatively from the force-pressure relationship, where force remains nearly constant; especially when direct measurements are difficult, for instance with small animals since measurements may be affected by changes in body position. Our average and individual data for the control group in Figs 4(B) and 6 are consistent with such a definition, as we found that the axial force was nearly unchanged with pressure inflation at the directly-measured in situ value of axial extension ratio, while the somewhat negative slopes of the average force-pressure data for the OVX groups, and especially of individual data, indicated a testing condition below the in situ axial extension ratio as defined by a constant force-pressure relationship. Given also the difficulties in interpreting our data, due to the absence of testing data at multiple axial extensions, the presented comparisons among groups should be seen with caution.
Notwithstanding this limitation, the observation of reduced in situ axial extension ratio in OVX rats (Table 2) is valuable and suggests that the physiologic environment of carotid arteries changed in response to OVX, so that the circumferential and axial stress-extension ratio responses of the control and all OVX groups be quite similar when examined at their individual in situ axial extension ratios (Fig. 5). Humphrey et al. [39] examined the biomechanical adaptation in diverse conditions, i.e. modified cell-matrix interactions in muscular dystrophy, modified matrix composition in fibrillin-1 deficiency, modified pulse pressure in aortic banding, and modified axial extension in organ culture, and found that mouse common carotid arteries modified drastically their axial extension ratio to normalize the overall stress state toward the homeostatic condition. Our results corroborate these important observations and add confirmation to the statement made by those authors that “Despite these different types of perturbation from normal, the arterial wall compensates in each case via a reduction in axial stretch, that is, an increase in unloaded length.” [39].
Structural and zero-stress state changes
The carotid artery suffered thickening, without change in lumen diameter, leading to an increase in their ratio and in wall cross-sectional area (Table 2 and Fig. 2). These findings are in line with the observations in [37] that carotid arteries from estradiol-treated rats had a significantly smaller wall area; these authors also reported that arteries from castrated rats had a significantly greater ratio of wall thickness to lumen radius compared to control rats. We additionally observed that the different wall layers grew at dissimilar rates, with the carotid arteries of OVX rats exhibiting a thicker media but not adventitia relative to control rats (Fig. 2).
The geometrical remodeling of tunica media in OVX rats was accompanied by an increase in collagen and elastin; similar was the trend for collagen in the adventitia. This similarity together with the growth of media led to increased collagen and elastin total wall percentage contents in OVX rats (Fig. 3). In most respects, these findings are not particularly unexpected; they seem consistent with early studies showing that exogenous estradiol treatment reversed the aortic growth in OVX female rodents or other animal models caused by the accumulation of elastin, collagen [40–42], and glycosaminoglycans [43], irrespective of body weight changes. Analogous results were more recently found in a human cell culture study [44] examining the direct effects of sex steroids devoid of their hemodynamic effects that are present in animal models. On the other hand, the initial age of OVX animals in the abovementioned studies was much less than that of the animals in the present report that more closely reflects the age of postmenopausal women.
Dobrin et al. [45] proposed that the axial retractive force in large arteries is primarily exerted by elastin, which is the main view held today. Elastin content was not reduced or fragmented in OVX rats to justify the significantly reduced axial extension ratio found herein (Table 2). This may rather be ascribed to arterial growth effected in both the circumferential and axial directions, as proposed by Langille et al. [46]. The in situ axial extension ratio we report about the control rats was considerably smaller than the 1.7-value reported in the literature; reference is made to the data by Weizsäcker et al. [38] on 9-month old rats and our recent study [22] on 6-month old rats. However, female rats of very large age were presently examined, i.e. 16-month-old, so that the
As a measure of the zero-stress state, we found that the opening angle varied with the morphological and structural remodeling. Specifically, the increased opening angle of the carotid artery in response to OVX associates directly with the increased ratio of wall thickness to lumen diameter (Table 2). It also corroborates Fung’s hypothesis [48], that is of the inner wall layers undergoing greater growth than the outer, as we found the media of the carotid artery from OVX rats to be significantly thicker and to occupy a greater cross-sectional area than that of control rats, unlike the adventitia that was equally thick (Fig. 2); noting that a reliable thickness/area measurement of the adventitia with its highly irregular structure around the circumference was not easy, and hence more likely to be afflicted with errors. Opening angle changes may also be attributed to changes in material properties, e.g. [49]. The increased opening angle in OVX rats may thus be associated to the stiffer carotid artery wall (Fig. 7). The consideration of age-related differences between the present and our previous control data on younger rats adds confirmation to well-known geometrical changes occurring with aging in the vasculature. Particularly, it discloses an age-related increase in opening angle (cf.
Consideration of the effects of SID
It has been observed that patient compliance to some of the current available treatments for osteoporosis, such as hormone replacement therapy and bisphosphonates is low, due to actual or fear of adverse effects [50,51]. Many patients therefore, in search of alternative protective/therapeutic options, resort to natural substances or ethnic functional foods [52,53]. Studies of the plant extracts that are being consumed to this end have demonstrated increased isoflavone and other compound levels, which have an affinity for estrogen receptors [54,55] and may thereby explain their bone-sparing effects during estrogen deficiency.
The extract from the Mediterranean plant Sideritis euboea that was used in the present study has been shown to enhance osteoblast proliferation in vitro [16], as well as to have a protective effect on bone mineral density and strength of OVX rats [17]. Unfortunately, this study does not report a beneficial effect as regards the structure, morphology, and mechanical properties of a large elastic artery, which is evidenced by the non-significant differences in any of these properties among the OVX,
Consideration of the hemodynamic aspects of arterial remodeling
Estrogen has a number of effects on cardiovascular function and can regulate the mechanical properties of arteries either by direct effects on the vascular cells or indirectly by systemic effects, so that it is important to identify the stimulus for the observed changes; i.e. is it a systemic action on, e.g. hemodynamics, or a direct action on smooth muscle cells? Furthermore, are the effects of OVX on arterial wall mechanical properties, morphology, and structure mediated by a physical (i.e. change in pressure/flow) or a biochemical stimulus (i.e. change in NO production) with estrogen deficiency? Systolic, diastolic, and mean blood pressures measured in the tail artery of OVX rats, which need to be seen with caution, did not differ from control whereas the increased pulse pressure may not be sufficient to explain a consistent hypertensive effect per se as it’s magnitude was not in the hypertensive range [56,57] and not likely to be great enough to induce the observed remodeling in OVX rats. Although we have no data on cardiac output, it is unlikely that it was much depressed in the absence of bradycardia that is on account of a reduced stroke volume and a previous investigation disclosed no variations in hindquarters and carotid blood flow in OVX rats [11]. We could also not study pressure variations among all groups, because the data from the
Concluding remarks
Estrogen depletion in postmenopausal women can produce cardiovascular system changes that can range from hypertension, atherosclerosis, arrhythmias up to cardiac arrest or stroke [58,59]. Animal studies are needed to determine the detrimental effects of depleted estrogen levels in the cardiovascular system and as such the present communication reports morphological, histological, and mechanical findings for the common carotid artery from middle-aged control (sham-operated) and OVX rats. Additional studies are clearly needed to provide more robust inflation/extension data for material characterization and to better define the time course of arterial changes following OVX, thereby offering important insight into the effects of estrogen on vascular structure and function.