
Abstract
BACKGROUND:
Biodegradable implant coatings promote proliferation and expression of BMP-2, VEGF, and TGF-β2 genes and enhance BMP-2, VEGF, and TGF-β2 regulatory effects at different stages of reparative osteogenesis.
OBJECTIVE:
To study the topography and ratio of PCNA-, VEGF-, BMP-2-, and TGF-β2-immunoreactive cells in rat femoral bone after closed fracture and implantation of titanium implants with biodegradable calcium phosphate and hydroxyapatite coatings.
METHODS:
Standard titanium implant screws and similar implants with bioactive coatings were used. A total of 18 rats were randomly divided into three groups, two experimental and a control one. The rats in the first experimental group were implanted with implants without specific coating, while those in the second group, with implants with specific coatings. The control rats were subjected to the same fracture as the experimental ones without subsequent implantation. On days 7, 14, and 30 of experiment, the rats were sampled for histological examination. Histological sections were prepared and processed for PCNA, BMP-2, VEGF, and TGF-β2 immunoreactivity.
RESULTS:
In the regeneration zone, PCNA-immunoreactive cells substantially outnumbered other immunoreactive cell types. During the first two weeks after fracture, in the immediate vicinity of implant surface, the rate of VEGF production increased in osteoblast subpopulations and level of TGF-32 immunoreactivity decreased in chondroblasts. The level of TGF-32 was maximum on day 30 of experiment. BMP-2-immunoreactive osteocytes were found in the zone of external general plates. They accumulated at implants with calcium phosphate coating. Their number gradually increased by day 30 of experiment.
CONCLUSIONS:
The present data suggest that biodegradable implant coatings promote proliferation and expression of BMP-2, VEGF, and TGF-β2 genes and enhance BMP-2, VEGF, and TGF-β2 regulatory effects at different stages of reparative osteogenesis.
Keywords
Introduction
Bone tissue reparation after injury involves various morphogenetic factors, whose local effects ensure the efficiency of reparative processes [1–3]. Bone tissue reparation can be promoted using a variety of osteoinductive and osteoconductive composites releasing growth factors and acting as implantable osteogenic matrix [4,5]. Another promising direction of implant biomedicine is development of implants with biodegradable calcium phosphate coating providing biologically active ions [6–8]. These ions may facilitate intercellular signalling and provide environmental kues for osteogenic cells essential for their stable proliferation [6,9,10]. Bone morphogenetic protein-2 (BMP-2), vascular endothelial growth factor (VEGF), and transforming growth factor-β2 (TGF-β2) participate in stimulation and control of osteoblast differentiation in posttraumatic reparation [11–13]. Further research is necessary to elucidate the role of these factors in reparative osteogenesis at the surface of osteogenic composites.
We studied cell proliferation and expression of BMP-2, VEGF, and TGF-β2 using a rat model after experimental femoral bone fracture followed by implantation of titanium implants with bioactive calcium phosphate and hydroxyapatitecoatings.
Materials and method
Implant samples
Standard titanium implant screws (titanium grade BT-6, produced by scientific production associations Deost and Osteomed, Russia) and similar implants with bioactive calcium phosphate and hydroxyapatite coatings were used in the study. Titanium grade BT-6 is identical to medical ASTM F136 titanium Ti-6Al−4V alloy. Prior to coating, the implants were treated mechanically to the roughness level of 0.12 μm. They were then washed in distilled water and degreased in ethanol. Plasma electrolytic oxidation coatings were produced using bipolar current mode in electrolyte containing calcium glycerophosphate ((C3H7O6P)Ca⋅2H2O, 30 g/l) and calcium acetate (Ca(CH3COOO)2⋅H2O, 40 g/l). The coatings were produced at impulse duration of 100 μs, impulse frequency of 50 Hz, coating time of 10 min or less, impulse tension of 150–400 V.
Experimental animals and fracture modelling
200–250 g male rats were used in the study. All experiments were performed in strict accordance with the local experimental animal treatment regulations and approved by the local ethical committee. The ethical standards of experiments were compliant with the CPCSEA and World Medical Association Declaration of Helsinki Guidelines on Ethical Principles for Medical Research Involving Humans. The animals were kept in a vivarium meeting standard requirements to animal keeping and treatment under experimental clinical conditions. All surgery was performed under aseptic conditions and sevoflurane anaesthesia. Low-invasive closed osteosynthesis resulting in indirect bone healing was modeled.
The surgical site was disinfected. To reach the femur cavity, an inlet hole was formed in the intercondylar notch using a thin trocar. To ensure screw cap placing in the subchondral layer, the hole was widened using an end reamer. A 2-mm implant screw was placed into the hole so that its end protruded from the middle thrid of the shaft. The bone was manually fractured. Bone fragments were manually set and fixed. The wound was closed and antiseptically treated. The fracture and correct implant position were confirmed by radiovisiography using an X-ray apparatus Evolution X3000 2C Asepti. X-ray graphs were taken in the side projection at 0.08 s exposure, 40 kV tube voltage, and 25 mAs tube current-time product (Fig. 1).

Radiograph of a rat femoral bone (side view) after experimental fracture and implantation. The arrow points at the fracture site.
A total of 18 rats were used in the study. They were randomly arranged into three groups, two experimental and a control one, each of six rats. The rats in the first experimental group (EGI) were implanted with implants without specific coating, while those in the second group (EGII), with implants with calcium phosphate and hydroxyapatite coatings. The control rats were subjected to the same fracture as the experimental ones without subsequent implantation. Their fractures were allowed to heal under blood clot. On days 7, 14, and 30 of experiment, a total of six rats (two from each group) were sampled for histological examination. The animals were euthanatized under general anaesthesia by intraperitoneal injection of 3% sodium thiopental followed by decapitation.
The material was fixed in 10% neutral formaldehyde on 0.1 M phosphate buffer for 24 h, washed in seven changes of 0.1 M sodium phosphate buffer (pH 7.2), decalcified in 25% Trilon B solution (pH 7.0) using Orbital Shaker OS-20 Biosan at room temperature, washed, and paraffin embedded following standard procedures. Ten μm thick sections were mounted onto slides, deparaffinized, and incubated with mouse monoclonal primary antibodies against proliferating cell nuclear antigen (PCNA), BMP-2, VEGF (Abcam), and TGF-β2 (Swant) diluted 1:200 in phosphate buffer with Triton X-100 and normal goat serum overnight at room temperature. The sections were then washed and incubated with biotinylated secondary goat anti-mouse antibodies diluted 1:100 (Vector Laboratories) for one hour. After that, the sections were inclubated in avidin-peroxidase complex solution (Vectastain ABC Kit, Vector Laboratories), developed in 0.1 M phosphate buffer with 0.03 % 3,3′-diaminobenzidine and 0.01 hydrogen peroxide for 10–20 min, washed, dehydrated, and mounted in Canada balsam following standard protocols. Some sections were counterstained with hematoxylin. Control sections were treated as described above, but were not incubated with primary antibodies. No specific labeling was found in these sections.
The specimens were observed using an AxioScope A1 microscope (Carl Zeiss). Photomicrographs were taken by means of an AxioCam ICc3 digital camera. Numerical parameters were estimated using the AxioVision 4.8.1 package. The spatial density of PCNA-, BMP-2-, VEGF-, and TGF-β2-immunoreactive cells was measured as cells per 0.01 mm3 of bone matrix, corrected for section thickness and nucleus diameter. The significance of differences was assessed using Student's t-test at p < 0.05.
Results
In the fracture site, the number of osteoblasts, microvessels, and fiber structures increased and amorphous material was formed and subsequently calcified. These changes were observed in the area of periosteum and endosteum and between edges of bone fragments of all posttraumatic samples, but the degree of osteogenic processes varied between different rat groups.
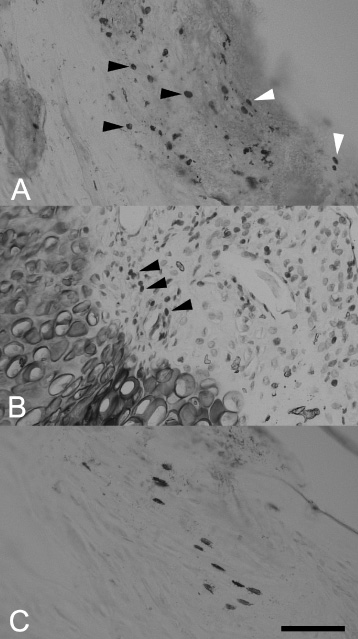
The PCNA reactivity in rat bone tissue after experimental fracture and implantation. (a). PCNA-positive osteoblasts (black arrows) and fibroblasts (white arrows) in the periosteum. Control group, posttraumatic day 7. (b) PCNA-positive osteoblasts (arrows) at the boundary of the new-formed bone and cartilage. Haematoxylin counterstaining. EGI, posttraumatic day 14. (c) PCNA-positive osteocytes in the zone of internal general plates. EGII, posttraumatic day 30. Parts (a) and (c) were combined from two focal planes each. Scalebar, 100 μm.
In the control group, on day seven after trauma, the number of osteoblasts increased accompanied by neoangiogenesis beneath the fibrous layer of the periosteum. Numerous PCNA-positive osteoblasts and fibroblasts were observed all across the periosteum (Fig. 2a). At the endosteum, groups of two to three PCNA-positive cells rarely occurred. These groups were irregularly distributed at the boundary of the new formed bone and cartilaginous tissue. The maximum PCNA reactivity was registered on day 14 of the experiment, when a hard callus was formed (Fig. 2b). Numerous joint lines were found here suggesting that bone matrix formation and resorption took place. On day 30, osteons were formed and penetrated with a plexus of fully developed vessels. PCNA positivity was observed in discrete subpopulations of osteocytes in the zone of external general plates (Fig. 2c). In both experimental groups, the intensity of osteogenesis was greater compared to the control as reflected in a significant increase in the number of PCNA-reactive cells at the vicinity of bioactive coating (Table 1).
Spatial density of immunoreactive cells (× 0.01 mm−3) in rat bone tissue after experimental fracture (mean ± SEM)

The BMP-2 reactivity in rat bone tissue after experimental fracture and implantation. (a) BMP-2-positive osteocytes (arrows) in the zone of external general plates. EGI, posttraumatic day 14. (b) BMP-2-reactive osteoblasts (arrows) in the periosteum. EGI, posttraumatic day 7. (c) BMP-2-positive osteoblasts (arrows) in the periosteum. EGI, posttraumatic day 30. Scalebar, 100 μm.
BMP-2-, VEGF-, and TGF-β2-immunoreactive cells were unevenly scattered in bone tissue. In the control group and EGI, BMP-2-positive osteocytes were mainly concentrated in the zone of external general plates (Fig. 3a). In EGII, the number of BMP-2-, VEGF-, and TGF-β2-immunoreactive osteoblasts in the periosteum significantly exceeded that in the control and EGI and gradually increased by day 30 of experiment (Fig. 3b, c).
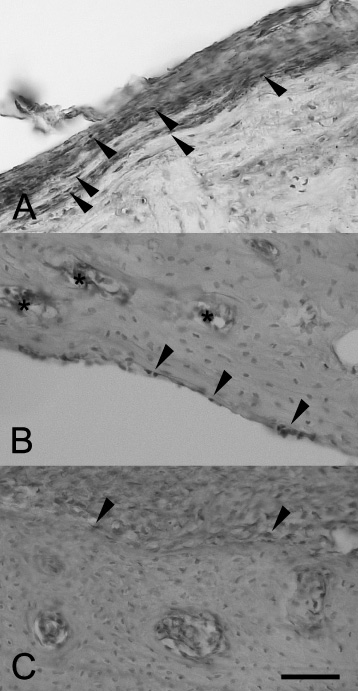
The VEGF reactivity in rat bone tissue after experimental fracture and implantation. (a) VEGF-positive osteoblasts (arrows) in the inner layer of the periosteum. Control group, posttraumatic day 7. (b) VEGF-reactive osteoblasts (arrows) in the endosteum and VEGF-positive endothelium cells (asterisks). EGI, posttraumatic day 14. (c) VEGF-positive osteoblasts (arrows) in the periosteum. EGII, posttraumatic day 30. Scalebar, 100 μm.
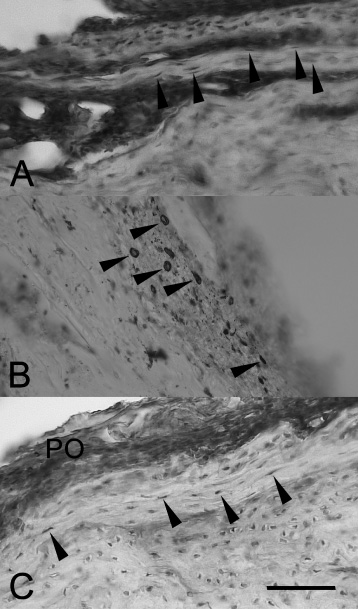
TGF-β2 immunoreactivity in rat bone tissue after experimental fracture and implantation. (a) Fibroblast-like cells (arrows) in the fracture zone. Control group, posttraumatic day 7. EGI, posttraumatic day 14. (b) TGF-β2-positive cells (arrows) in the periosteum. EGI, posttraumatic day 14. (c) TGF-β2-positive cells (arrows) in the periosteum (PO) and external general plates. EGII, posttraumatic day 30. (a), (c) - haematoxylin counterstaining. Scalebar, 100 μm.
In the control group and EGI, individual VEGF-positive osteoblasts were observed. They were diffusely scattered in the periosteum and at the endosteum, often in the vicinity of VEGF-positive microvessels (Fig. 4a, b). In EGII, VEGF-immunoreactive osteoblasts were concentrated at the surface of implants with calcium phosphate coatings (Fig. 4c), and the number of endotheliocytes significantly exceeded that in the control and EGI (Table 1). The number and distribution of TGF-β2-immunoreactive cells in the control and EGI were essentially the same. The TGF-β2 immunoreactivity in these groups was restricted to subpopulations of osteoblasts and/or fibroblast-like cells in the periosteum (Fig. 5a, b). The number of these cells gradually decreased from day seven to day 14 of experiment (Table 1). In EGII, the number of TGF-β2-immunoreactive cells and staining intensity increased substantially on day 30 of experiment (Fig. 5c).
The distribution of PCNA, BMP-2, VEGF, and TGF-β2 positivity suggests that the reparative osteogenesis occurred most actively in the vicinity of implants with bioactive coatings.
Implant integration and efficiency depend on osteogenesis and reparative processes in the surrounding tissues. The chemical activity of a coating, its resorbtion, and interaction with bone matrix are determined by several parameters, in particular, the amount and proportion of calcium and phosphorus and coating roughness and porosity [9,14]. Increasing coating roughness facilitates attachment and adhesion of stem cells and osteoblasts to the implant surface under variable mechanical stress and may promote osteogenesis. This is confirmed by the fact that poor, if any, osteogenesis was observed after implantation of implants with smooth hydroxyapatite coatings [11,15,16]. The roughness and other physical and chemical properties of the implant coatings presently tested correspond to those required for stimulating osteogenesis and implant integration after consolidated fracture [17,18].
Implant-tissue interface is a critical point in reparative osteogenesis. It is here where implant coatings interact with surrounding tissue elements stimulating a variety of morphogenetic events including cell death and regeneration. For instance, the proliferation of stromal stem cells and their differentiation to osteoblasts are regulated by BMP-2 and BMP-7 [14,19]. Stromal cell-derived factor-1 (SDF-1) is expressed in the fracture zone, especially, periosteum and interfragmentary space [13]. BMP-2 is a potent osteoinductive cytokine capable of induction of chondrogenesis and playing a key role in osteoblast differentiation and apoptosis [12,15].
Soon after trauma, an acute inflammation develops and macrophages begin to express inflammatory cytokines including tumor necrosis factor-α (TNF-α) and interleukins 1 and 6 [20]. TNF-α promotes the differentiation of stromal stem cells into osteogenic cells [20], while interleukins 1 and 6 stimulate angiogenesis by activating VEGF expression and differentiation of osteoblasts and osteoclasts [21]. VEGF is the only growth factor involved in osteogenesis at all stages beginning with interfragmentary hematoma that is formed during the first hours after fracture and ending with callus remodelling several months later [22]. VEGF facilitates primary vascularization of the callus suggesting that adventitial and endothelial cells are its primary targets in the periosteum. Excessive VEGF administration to the wound area speeds up fracture union, while blockade of VEGF receptors slows it down [2,21]. In the present study, VEGF was hyperexpressed in cells lying in close proximity to implant coating. We suggest that calcium phosphate coating may enhance VEGF expression.
An important prerequisite of successful bone remodeling and tissue replacement is sufficient blood supply of the fracture zone. The early hematoma is replaced by a soft callus formed by connective tissue rich in fibrin. Later on, intramembranous ossification begins in the subperiostal zone, and the soft callus is remodeled and ossified [3]. These processes are largely controlled by members of the TGF superfamily, especially, TGF-β2 and TGF-β3. These factors regulate collagen production, hondrogenesis, and enchondral ossification [23,24]. Depending on the amount of secreted intercellular matrix, TGF-β2 may inhibit its degradation in cartilage and, partially, bone. During bone tissue resorption, especiaaly, at early stages of fracture union, osteoclasts enhance TGF-β2 production [25]. TGF-β1 may block signal pathways mediated by BMP-2 and BMP-7, thereby inhibiting bone tissue reparation [12]. The present results are generally consistent with the known dynamics of TGF-β2 activity. Hydroxyapatites may stimulate TGF-β2 expression. An increase in the TGF-β2 level in EGII at late osteogenesis suggests that it participates in the formation of mature callus at late ossification. It is unclear what physical or chemical properites underly osteoinductive capacity of calcium phosphate coatings. We suggest that bioactive ions affect small molecules involved in intra- and intercellular signal pathways. Such osteoinductive effects may be induced by small bioactive molecules modulating the expression of bone morphogenic protein, mitogen activated protein (MAP) kinase, and canonical Wnt signaling [26].
Thus, the present data suggest that BMP-2, VEGF, and TGF-β2 are involved in osteogenesis activation by calcium phosphate bioactive coating. Studying molecular and cellular mechanisms of this process is an important step towards modeling specific and directed regeneration after implantation of metal implants with biodegradable coatings.
Footnotes
Funding
This study was supported by a grant from the Russian Science Foundation (RSF) under grant no. 14-33-00009.
Conflict of interest
The authors report no conflict of interest. The authors alone are responsible for the content and writing of the article.