
Abstract
BACKGROUND:
Collagen receptors are characterized by binding to and being activated by collagens. We know little about the molecular mechanism by which the integrins and discoidin domains (DDRs) recognize collagen.
OBJECTIVE:
The aim of this study was to investigate the expression of two main collagen receptor subfamilies, integrins and DDRs, during osteogenic and chondrogenic differentiation of human mesenehymal stem cells (hMSCs).
METHODS:
Using qRT-PCR, Western blots and FACS, the levels of DDR1, DDR2, integrin subunits β1, α1, α2, α10 and α11 receptors on hMSCs, were assessed upon activation by collagen type I, as well as during osteogenic and chondrogenic differentiation.
RESULTS:
The expression of DDR2 and integrin α11β1 was altered compared with other receptors when the cells were cultured under undifferentiated conditions. During osteogenic and chondrogenetic differentiation, DDR2 and α11 were up-regulated during early stages (6 day) of osteogenesis and chondrogenesis, respectively. The expression and activation of DDR2 was concomitant with another receptor integrin subunit β1 during osteogenetic differentiation.
CONCLUSIONS:
The results suggested that DDR2 was more specific for osteogenesis than chondrogenesis, while integrin α11β1 was more specific in chondrogenesis. DDR2 and α11 may play a role in the regulation of osteogenesis and chondrogenesis based on the differential expression of these receptors during lineage-dependent changes.
Introduction
The most widely expressed mammalian receptors for collagen are four members of the β1 integrin family and the discoidin domain receptors (DDRs). Integrins are a major class of the extracellular matrix (ECM) receptors for cell adhesion [1]. They are heterodimeric transmembrane glycoproteins that are composed of non-covalently associated α and β subunits. There are four collagen-binding integrins in mammalian cells: α1β1, α2β1, α10β1 and α11β1 [2–4]. The α subunit of these collagen receptors have an inserted (I) domain in their extracellular region that contains the collagen-binding site. The most widely distributed collagen-binding integrins are α1β1 (predominantly found in mesenchymal cells) and α2β1 (predominantly found in epithelial cells). α1β1 was expressed at the surface of mouse chondrocytes [5]. DDRs are receptors tyrosine kinases that recognize collagens as their ligands. DDRs constitute a unique subfamily of receptor tyrosine kinases containing a discoidin homology region in their extracellular domain [6]. There are two related DDRs: DDR1 and DDR2 are specifically expressed in epithelium and mesenchyme, respectively [6,7]. DDR2 was expressed and induced in the mouse osteoblasts and chondrocytes [8].
Both DDR and collagen-binding integrins bind a number of collagens with distinct specificity and recognize specific amino acid motifs within the collagen triple helix. α1β1 integrin is able to bind collagen types I, II, III, and V [3,9–11], and favor other subtypes such as type IV [10], while α2β1 integrin is the major receptor for type I collagen [9,11]. Currently, less is known about ligand binding preferences of the novel α10β1 and α11β1 integrins. DDR1 is primarily activated by type I–IV and VIII [12,13], whereas DDR2 responds to collagen I and to a lesser extent to type II, III, and V [12–14].
The interactions of collagens with integrin collagen receptors have been characterized, in particular for integrin α1β1 and α2β1. In addition to their importance as anchoring molecules, these heterodimeric receptors transmit bidirectional signals that regulate many important aspects of cell behavior including proliferation, differentiation, and survival [2,15,16]. Collagen-induced DDR signaling controls cell proliferation, adhesion and migration, as well as re-modeling of extracellular matrices [17–20]. For example, DDR receptors regulated the migration of primary human lung fibroblasts through collagen matrices and DDR2 was shown to be involved in fibroblast proliferation [21]. But very little is known about the collagen receptors and their expression, or the involvement during cell differentiation.
Interactions between proteins within the ECM and their receptors on the surface of mesenchymal cells might provide environmental signals that contribute to the progression of differentiation. Previous literature has shown that ECM proteins influence differentiation on hMSCs derived from bone marrow in in vitro cultures [22–25]. Type I collagen, the main ECM protein, was able to induce the osteoblastic differentiation of bone marrow cells, and furthermore promoted mineralization and up-regulated the osteogenic genes such as transcription factor Cbfa-1 and osteocalcin [26]. On the other hand, collagen receptors that bind to these ECM proteins promoted and inhibited differentiation of hMSCs. Previous studies showed that interactions of α2β1 integrin with collagen type I is required for osteoblastic differentiation of hMSCs [26,27] and the expression of osteogenic genes [28]. However, detailed studies of the expression of collagen receptors during the differentiation of hMSCs has not been described, particularly with regard to the interactions between collagen type I and collagen receptors during differentiation. The goal of the present study was to explore the expression of these collagen receptors during the chondrogenic and osteogenic differentiation of hMSCs, as well as to assess their roles in regulating collagen I-induced differentiation.
Materials and methods
Cell cultures
hMSCs were obtained from commercially available bone marrow aspirates from one male donor ≤25 years of age (Clonetics-Poietics, Walkersville, MD, USA) using methods previously reported [29]. Whole bone marrow aspirates were plated at 8–10 μl aspirate/cm2 on 185 cm2 tissue culture plates and cultivated until confluency (12–14 days) in 40 ml of ex vivo expansion medium consisting of Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum (FCS), 100 U/ml penicillin, 100 μg/ml streptomycin, 0.1 mm nonessential amino acids, and 1 ng/ml of basic fibroblast growth factor (bFGF) (Life Technologies, Rockville, MD, USA). hMSCs were maintained in a humidified tissue culture incubator at 37 °C with 5% CO2. hMSCs were selected based on their ability to adhere to the tissue culture plastic; nonadherent hematopoietic cells were removed during medium replacement after approximately 5 days in culture. Medium was changed twice per week thereafter. hMSCs were subsequently detached using 0.25% trypsin/1 mM EDTA, replated at 5 × 103 cells/cm2, and cultured to 90% confluency to passage 1 cells.
Osteogenic and chondrogenic differentiation
Osteogenic differentiation medium included standard hMSC culture medium as listed above with the exception of 1 ng/mL bFGF. In addition, 0.05 mM L-ascorbic acid 2-phosphate (Sigma, USA), 1000 nM dexamethasone (Sigma, USA), 10 mM glycerol 2-phosphate disodium salt hydrate (Sigma, USA), and 100 ng/mL bone morphogenetic protein (BMP)-2 was added (Promocell, Germany). Passage 1 hMSCs were seeded at a density of 3 × 104 cells/cm2. To induce chondrogenic differentiation, cells were plated at a 1 × 105 cells per well in a 96-well U-bottom suspension culture plate. Chondrogenesis was induced by culturing hMSCs in the presence of a defined hMSC medium that included 100 nM dexamethasone and 10 ng/ml transforming growth factor-beta(3) (TGF-beta(3)) (Promocell, Germany). Differentiation medium was changed two times per week.
Fluorescence-activated cell sorting (FACS)
FACS analysis was used to determine surface expression of DDR2 and integrin α11 in hMSCs. Membrane staining was performed with fluorescently labelled antibodies: anti-human DDR2 (R&D Systems (#290804); Abcam (ab98702), USA), and anti-human integrin α11 (Acris Antibodies Inc. (AP52249PU-N and R1255F), USA). Passage 1 cells were detached with 0.25% trypsin–EDTA, pelleted and resuspended in 300 μl DMEM. Cell suspensions were split into three aliquots; a control group without staining, anti-DDR2 or α11, and anti-human CD29 (known as integrin β1, Biolegend (303015), USA) conjugated with Alexa Fluor® 488 as positive control. After addition of antibodies the samples were mixed and incubated for 30 min in the dark at 4 °C. The cells were washed with 1.5 ml DPBS and centrifuged for 5 min at 1500 rpm, 4 °C. The supernatant was removed, the cell pellet was suspended in 1 ml DPBS in polystyrene tubes (Falcon) and subjected to FACS analysis. Data was analyzed using FlowJo FACS analysis software 7.6 (Treestar Inc., USA).
mRNA expression analysis
Total RNA was extracted from the cells using an RNeasy Mini Kit (Qiagen, USA), following the supplier’s instructions. Extracted total RNA was used to synthesize cDNA using a High-Capacity cDNA Archive kit (Applied Biosystems, USA), following the supplier’s instructions. mRNA levels were determined either by quantitative real time PCR or by semi-quantitative reverse-transcriptase PCR (RT-PCR). Real time RT-PCR reactions were conducted and monitored using an Mx3000P (Stratagene®). TaqMan Gene Expression System (Applied Biosystems, USA) with predesigned primer/probe sets were used to determine transcript levels of human genes DDR1, DDR2, integrin α1, α2, α10, α11 and β1 subunits, alkaline phosphatase (ALP), aggrecan and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The amplification program was as follows: 50 °C for 2 mins; 95 °C for 10 mins, followed by 40 amplification cycles of 95 °C for 15 seconds; and 60 °C for 1 min. Amplification specificity was validated by melting curve analysis. The mRNA expression changes (fold difference) were calculated after normalizing with GAPDH as an endogenous control. For RT-PCR, the number of amplification cycles was set at 25 to ensure that the amplifications were analyzed within an exponential phase. All PCR reactions from each cDNA sample were run in triplicates.
Western blots
Cells were harvested and lysed in radioimmunoprecipitation assay (RIPA) buffer (Invitrogen, USA). Protein concentrations were measured using the BCA protein assay (Thermo Fisher Scientific Inc., USA). The protein extract were resolved by 10% sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to Hybond-ECL nitrocellulose membranes (Invitrogen, USA). After blocking in Tris-buffered saline containing 5% BSA and 0.1% Tween-20, membranes were probed with primary antibodies against human DDR2 (R&D Systems (AF2538), USA) and integrin α11 (Acris Antibodies Inc. (AP52249PU-N), USA). Beta-actin was used as an internal positive control. For detection, the membranes were incubated with the appropriate secondary antibodies conjugated to horseradish peroxidase and the blots were visualized with enhanced chemiluminescence.
Statistical analysis
All studies were performed in triplicate, and all experiments were repeated at least three times (n = 3). Results are presented as mean ± SD.
Results
Expression profile of collagen receptors in hMSCs
First, we investigated collagen receptor expression on both TCP and collagen type I substrates under undifferentiated conditions to profile the expression of collagen receptors in hMSCs. After 6 days of culture, mRNA expression was detected for collagen receptors DDR1, DDR2 and integrin subunits β1, α1, α2, α10 and α11. Integrin α11β1 and DDR2 showed a moderate to high level mRNA expression compared with the expression level of other collagen receptors, and both receptors had a higher mRNA level expression on collagen type I than on TCP. α1, α2, and α10 had a low expression. In contrast, the expression of DDR1 was not affected in hMSCs (Fig. 1).

hMSCs were cultured on TCP or TCP coated with type I collagen (Invitrogen, USA) for 6 days and mRNA was extracted to detect expression profiles of collagen receptors in hMSCs. RT-PCR was carried out with the indicated primers DDR1, DDR2, integrin subunits α1, α2, α10, α11 and β1 (A). The results are representative of mean fold increases ± S.D. of three independent experiments done in triplicate. For semi-quantitative RT-PCR, the number of amplification cycles was set at 25 to ensure that the amplification was analyzed within the exponential phase. PCR products were run on 2% agarose gels (B).
To test whether the expression level of DDR2 and integrin α11 subunit was expressed in hMSCs, we analyzed cell surface receptors level using FACS, and integrin β1 was used as positive control. As shown in Fig. 2, cell surface receptor expression levels of α11 was consistent with the mRNA expression level, while DDR2 showed a decrease when compared with the mRNA results.
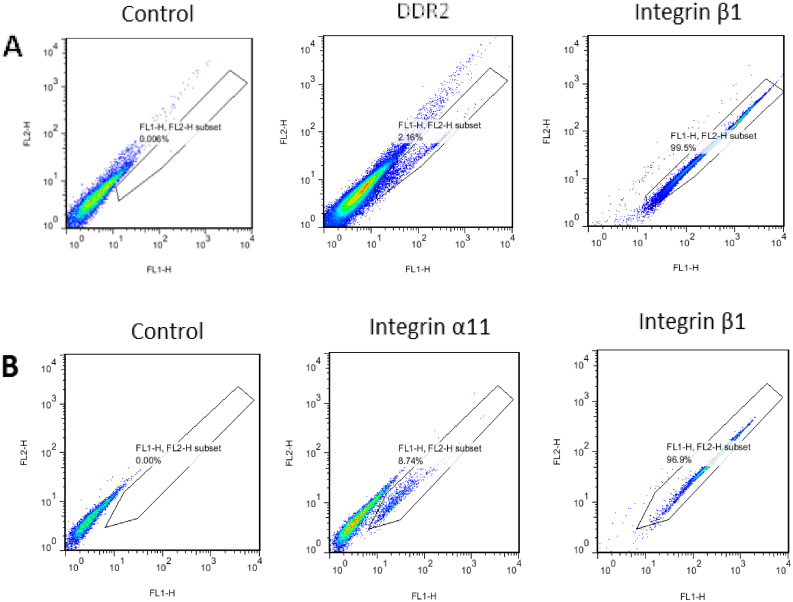
hMSCs were collected after 6 day culture. The cells were incubated and stained with anti-DDR2, anti-integrin α11 and anti-integrin β1 antibodies. Flow cytometry was performed using FACSCalibur TM System (BD, USA). The frequency of positive results in the selected population is shown. DDR2 and α11 subunit showed 2.56% (A) and 8.72% (B) positive labeling, respectively, compared with negative and positive β1 subunit controls. The results are representative of mean ± S.D. of two independent experiments.
We further examined activation dynamics of collagen receptors during differentiation to see whether collagen receptor expression was altered by the differentiation commitment of hMSCs. As shown in Fig. 3, hMSC were harvested after culturing in osteogenic and chondrogenic medium for 6 days. The mRNA expression of collagen receptors DDR1, DDR2 and integrin subunits α1, α2, α10, α11 and β1 were examined separately by RT-PCR. Cells exhibited an osteoblastic phenotype at 6 days, which was determined by the increase in osteogenic marker expression. The chondrogenic commitment was assessed by chondrogenic marker expression. The expression of markers indicated early osteogenic and chondrogenic activities separately.
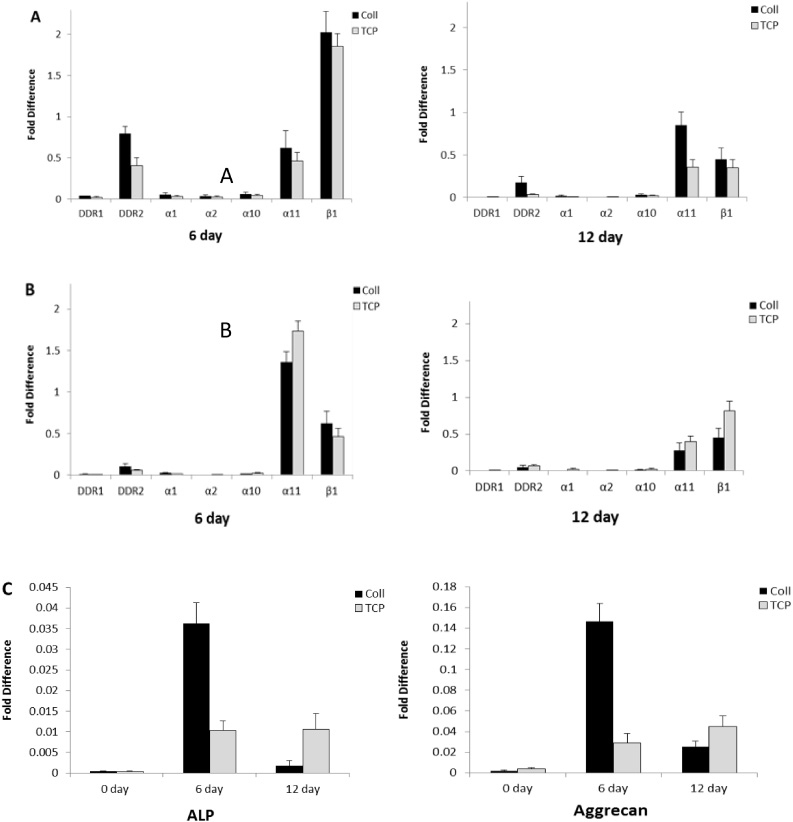
hMSCs were cultured on TCP or TCP coated with type I collagen in osteogenic and chondrogenic medium for 6 and 12 days and were harvested for total RNA isolation. RT-PCR was carried out with the indicated primers DDR1, DDR2, integrin α1, α2, α10, α11 and β1 subunits. The relative mRNA expression levels of collagen receptors was detected at different day points during osteogenic (A) and chondrogenic (B) differentiation. mRNA levels of ALP and aggrecan, osteogenic and chondrogenic markers, was detected at 0, 6 and 12 day (C). The results are representative of mean fold increases ± S.D. of three independent experiments done in triplicate.
mRNA expression of DDR2 and integrin α11β1 were peaked at day 6, followed by downregulation at 12 day in osteogenesis. Integrin subunit β1 showed a higher increase than α11 at 6 day, both subunits α11 and β1 decreased at 12 day. All collagen receptors showed higher mRNA expression when hMSCs were cultured on type I collagen substrate than on TCP during osteogenic differentiation. In chondrogenic differentiation, mRNA expression level of subunit α11 showed an increase at day 6 compared with the expression level of DDR2 that showed a decrease. α11 showed the same decrease as DDR2 at 12 days. α11 showed less mRNA expression on type I collagen substrate than on TCP at 6 and 12 days. The results suggested that integrin α11β1 was more active than DDR2 in chondrogenesis.
Western blots were carried out to measure DDR2 and α11 protein expression as indicated in Fig. 4. In osteogenesis, we focused on the role of DDR2 in osteogenic differentiation. DDR2 protein abundance showed an increase at day 6 and tended to decrease at 12 day with higher protein expression on type I collagen than on TCP, which was consistent with the changes in mRNA. These results suggest that the differentiation-dependent fluctuation of DDR2 expression level is a common feature of hMSCs. In chondrogenic differentiation, the α11 protein expression showed the same fluctuation tendency as the DDR2 expression, while α11 showed the same or a slight increase in protein expression on TCP than as on type I collagen during osteogenesis, which was different from the DDR2 protein expression during osteogenesis. These results suggested that the expression of integrin α11β1 was different from DDR2 by activation of type I collagen during chondrogenesis.
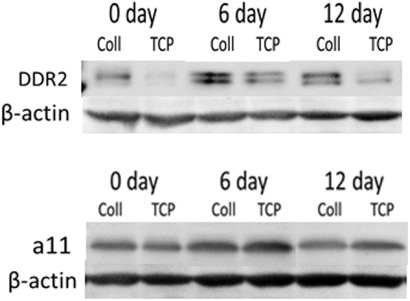
hMSCs were cultured on TCP or TCP coated with type I collagen in osteogenic and chondrogenic medium. Total protein was isolated after cells were lysed, and subjected to SDS-PAGE followed by anti-DDR2 and anti-α11 immunoblotting. Anti-β-actin was used as loading control. Protein expression of DDR2 during osteogenesis at different days and substrates was immunoblotted (A). The same commitment for integrin subunit α11 was performed in chondrogenesis (B). The results are representative of two independent experiments.
mRNA expression of both DDR2 and integrin subunit β1 increased at day 6 and tended to decrease at 12 day during osteogenic differentiations, and there was a higher expression level on type I collage than on TCP. The same association between DDR2 and subunit β1 was also found by activation of type I collagen during chondrogenesis, while this association was not found on TCP. These results suggested that mRNA expression of DDR2 was concomitant with the expression of subunit β1 by activation of type I collagen (Fig. 5).
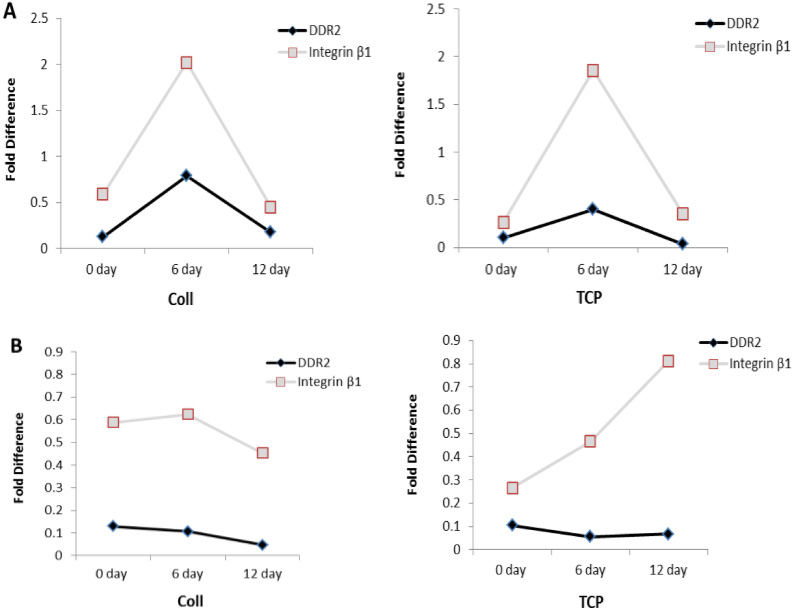
mRNA expression level of DDR2 and integrin subunit β1 at 0, 6 and 12 days on type I collagen and TCP substrates during osteogenic (A) and chondrogenic (B) differentiation. The results are representative of two independent experiments.
The expression and activation of two main collagen receptor subfamilies in hMSCs were initially profiled and DDR2 and integrin α11β1 were significantly altered compared with other receptors α1, α2, and α10. We further investigated the expression and activation of collagen receptors in hMSCs during osteogenic and chondrogenetic commitment. DDR2 and α11 were up-regulated during early stages of osteogenesis and chondrogenesis separately. The phenotypic similarities between DDR2- and collagen-deficient mice support in vitro evidence for DDR2 regulation of collagen synthesis in osteoblasts and chondrocytes [30–34]. A recent study showed that DDR2 played an important role in osteoblast differentiation and chondrocyte maturation via modulation of runx2 activation and also played an essential role in chondrocyte hypertrophy [8]. Notably, a recent report described a mutation in the human DDR2 gene that caused spondylometaepiphyseal dysplasia (SMED), a rare autosomal recessive disease [35].
We examined dynamic changes in DDR2 expression and collagen I-induced expression during osteogenic differentiation and found that DDR2 activation by type I collagen was paralleled by the changes in DDR2 expression on TCP. Maximal DDR2 expression was observed in mature osteoblasts at early stages of hMSC differentiation, suggesting that the differentiation-induced ECM microenvironment is required for full DDR2 expression. The unresponsiveness of hMSCs to DDDR2 upregulation under undifferentiated conditions is likely due to the lack of sufficient differentiation stimulation to DDR2. However, we did not find DDR2 upregulation of expression and activation during chondrogenic differentiation, unlike from previous research where DDR2 was active in both osteogenesis and chondrogenesis [8,35]. Thus these data suggest that DDR2 functions in osteogenic differentiation correlated with its activation by type I collagen.
Integrin α11β1 constitutes a recent addition to the integrin family [36]. α11 expression was present in various mesenchymal cells around cartilage anlage during human embryonic development, suggesting that α11 might be associated with the chondrogenic differentiation of mesenchymal cells [36]. We found upregulation expression of α11 during chondrogenic differentiation, and that its collagen I-induced expression was not paralleled by changes in α11 expression. Although both DDR2 and α11 showed a binding preference to type I collagen, they showed opposing expression by collagen I-induced expression during osteogenetic and chondrogenic differentiation, implying that differentiations has different effects on the expression and activation of DDR2 and α11 in hMSCs. The present results suggest that a11 plays a different role in chondrogenic differentiation unrelated to its activation by type I collagen.
The interplay between the DDRs and collagen-binding integrins has been observed in some cellular systems [37]. A recent study suggested that DDRs promoted α1β1- and α2β1- integrin mediated cell adhesion to collagen by enhancing integrin activation [37]. Interestingly, in our study, we found that the expression and collagen I-induced expression of DDR2 may be concomitant with another receptor integrin subunit β1 during osteogenetic differentiation. Little is known about DDR2 and β1 has been shown to be both required for cell adhesion on collagen in some cell types, and they may have mutual effects on the differentiation of mesenchymal cells.
Conclusion
In conclusion, the results presented in the present study suggest that DDR2 and integrin α11β1 play an important role in the differentiation of mesenchymal stem cells. DDR2 regulated the osteogenetic differentiation of hMSCS by activation by type I collagen, and for the first time we found that α11 played an important role in chondrogenic differentiation. DDR2 and integrin α11β1 functioned differently in osteogenesis and chondrogenesis. The data suggest that DDR2 was more specific in osteogenesis, while integrin a11b1 was more specific in chondrogenesis than in osteogenesis. DDR2 and α11 may play a role in the regulation of osteogenesis and chondrogenesis based on the differential expression of these receptors during lineage-dependent changes. The expression and activation of DDR2 may be concomitant with another receptor integrin subunit β1 during osteogenetic differentiation. Unraveling the importance of DDR2 and integrin α11β1-involved activity in mesenchymal cell differentiation represents an important direction related to control of stem cell fate and tissue regeneration.
Footnotes
Acknowledgements
The authors thank Bo An, Birgit Leitinge, Barbara Brodsky and David L. Kaplan for their contributions to this study.
Conflict of interest
None to report.