
Abstract
BACKGROUND:
Calcium phosphates including β-tricalcium phosphate (β-TCP) and hydroxyapatite (HAp) have been widely used for bone regeneration application because of their high osteoconductive activities. In addition, various kinds of inorganic ions enhance differentiation, proliferation, and mineralization of osteoblasts. However, information about the effects of silver-doped β-TCP [β-TCP (Ag)] and HAp [HAp (Ag)] particles on osteogenic differentiation is not available yet.
OBJECTIVE:
We focused on the impact of β-TCP (Ag) and HAp (Ag) particles on the osteogenic differentiation of MC3T3-E1 osteoblast precursor cells.
METHODS:
MC3T3-E1 osteoblast precursor cells were pre-treated by β-TCP (Ag) or HAp (Ag). And then the medium was changed to differentiation medium. Subsequently, osteoblast differentiation-related markers were determined.
RESULTS:
We found that treatment with β-TCP (Ag) or HAp (Ag) particles increased alkaline phosphatase activity in MC3T3-E1 cells. Expression of osteoblast differentiation-related genes also increased after treatment with β-TCP (Ag) or HAp (Ag) particles, a response thought to be regulated by zinc finger-containing transcription factor osterix. The ratio of the receptor activator of nuclear factor kappa-B ligand (RANKL) to osteoprotegerin (OPG) was decreased by β-TCP (Ag) and HAp (Ag) particles.
CONCLUSION:
Silver doping of β-TCP and HAp particles is effective for bone regeneration.
Introduction
Bone defects caused by injury, tumor, inflammation, or congenital disorders, are a significant problem all over the world, therefore, effective therapies for bone regeneration is demanded [1]. Currently, a variety of biomaterials based on inorganic salts and polymeric materials are being used for promoting bone regeneration and repair in the clinical field. These materials are required to show a favorable effect on bone formation by stimulating osteoblastic proliferation and differentiation [2]. Calcium phosphates (CaPs), mainly as β-tricalcium phosphate (β-TCP, Ca3(PO4)2) and hydroxyapatite (HAp, Ca10(PO4)6(OH)2), have been widely used for bone regeneration application because they possess high osteoconductive activities [3]. Calcium (Ca2+) and phosphate (PO
In addition to Ca2+ and PO
However, the osteoblastic differentiation-inducing activity of silver-doped calcium phosphate has not yet been evaluated. Silver-doped calcium phosphate has already been used as a bone material, but the main purpose is to prevent infectious diseases during clinical application. There are some reports on the antibacterial activity of silver-doped calcium phosphate [13–15]. Therefore, in this study, in addition to antibacterial activity, osteoblast differentiation-inducing activity was evaluated. Antibacterial activity and osteoblast induction promoting effect are advantages as a material for clinical use of this material.
The benefits of microparticles (MPs) or nanoparticles (NPs) are their high specific surface area, which increases the dissolution rate, and their small size, which allows them to be engulfed by the cells to deliver their therapeutic ions [16]. However, both in vivo and in vitro investigations have raised concern that MPs and NPs may be related to cytotoxicity and genotoxicity by oxidative stress induction and inducing pro-inflammatory responses [17,18]. In addition, particulate β-TCP and HAp increased the production of pro-inflammatory cytokines, such as tumor necrosis factor (TNF)-alpha, interleukin (IL)-1β, IL-6, and IL-8 [19–22], and induced local inflammatory responses and bone resorption [23]. In order to reduce the pro-inflammatory effects of these particles, researchers have been manipulating the immune reaction by controlling the release of bioactive ions. For example, Mg2+, Sr2+ and Zn2+ could effectively downregulate the pro-inflammatory responses caused by β-TCP and HAp exposure [24].
Therefore, in this study, we investigated the osteogenic effects of Ag-doped β-TCP [β-TCP (Ag)] and Ag-doped HAp [HAp (Ag)] particles on MC3T3-E1 cells.
Material and Methods
Characterization of particles
β-TCP was provided by Shiraishi Central Laboratories Co., Ltd. (Hyogo, Japan) and Ag-doped β-TCP [β-TCP (Ag)] and HAp particles were obtained from Sangi Co., Ltd. (Tokyo, Japan). Ag-doped HAp [HAp (Ag)] particles were obtained from Nanoshel LCC (Wilmington, DE, USA). Particle microstructures were observed by scanning electron microscopy (SEM, JSM-6330F; JEOL Ltd., Tokyo, Japan). Crystallographic properties of the particles were recorded by X-ray diffractometer (XRD, MultiFelx; Rigaku Corp., Tokyo, Japan). Median diameter was determined using a laser diffraction particle size analyzer (Master Seizer 3000; Malvern Panalytical, a division of Spectris Co. Ltd., Hyogo, Japan). Specific surface area was measured by the Brunauer–Emmet–Teller (BET) method using a Macsorb1208 (Mountech Co., Ltd., Tokyo, Japan). Silver concentration was analyzed using atomic absorption spectrometry (iCE 3300; Thermo Fisher Scientific K.K., Tokyo, Japan). Particle-medium suspensions were prepared as described previously [25]. In brief, the powder was suspended in 1% bovine serum albumin at a concentration of 10 mg/mL prior to use. The BSA-suspensions were sonicated for 30 min, subsequently, the suspensions were diluted in culture medium for cellular experiments.
Cell culture
Mouse calvaria derived preosteoblast MC3T3-E1 cells were purchased from the RIKEN BioResource Center (Ibaraki, Japan) and cultured in minimum essential medium alpha (MEM𝛼; ThermoFisher Scientific Inc., Waltham, MA, USA) containing 10% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich, Saint Louis, MO, USA), 100 U/mL penicillin, 100 μg/mL streptomycin, and 250 ng/mL amphotericin B (Nacalai Tesque Inc., Kyoto, Japan) at 37 °C in an atmosphere of 5% CO2. For experiments, the cells were seeded in different-sized multi-well plates (ThermoFisher Scientific Inc.) at a density of 5 × 104 cells/mL and incubated for 48 h. The culture medium was replaced with fresh medium in the presence or absence of β-TCP or HAp particles, and this was followed by incubation for 24 h. All cultures were washed with phosphate-buffered saline (PBS) and supplied with osteogenic induction medium containing 50 μg/mL ascorbic acid (AA) and 5 mM β-glycerophosphate (β-GP; Sigma-Aldrich). This medium change was referred to as differentiation day 0. The medium was replaced with fresh osteogenic induction medium every 2–3 days thereafter. All experiments were carried out using cells between passage 6 and 15.
Measurement of cell viability and cell membrane damage
Cell viability and cell membrane damage were determined by WST-1 assay and lactate dehydrogenase (LDH) release using a Premix WST-1 Cell Proliferation Assay System (TaKaRa Bio) and a Cytotoxicity Detection KitPLUS (LDH) (Roche Diagnostics GmbH, Mannheim, Germany) according to the manufacturer’s instructions, respectively.
Alkaline phosphatase activity assay
Intracellular alkaline phosphatase (ALP) activity was measured using a TRACP & ALP Assay Kit (TaKaRa Bio) according to the manufacturer’s instructions. The values of ALP activity were normalized with the total protein content which was determined using a Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Inc.).
Alizarin red staining
Mineralization of MC3T3-E1 cells was determined using Alizarin red staining. The cells were fixed with 70% ethanol at 4 °C for 1 h followed by washing with distilled water. 1% Alizarin red staining solution (Muto Pure Chemicals Co., Ltd., Tokyo, Japan) was then added to the cells, which were then incubated at room temperature for 1 min, followed by removal of stain and washing with PBS. For quantification, cells stained with Alizarin red were destained with 10% ethylpyridinium chloride (Tokyo Chemical Industry Co., Ltd., Tokyo, Japan), and then the absorbance at 562 nm was measured using Tecan Infinite M200 (Tecan Group Ltd., Mannedorf, Switzerland).
Isolation of total RNA and quantitative real-time PCR
Total RNA was extracted using a RNeasy Mini Kit (Qiagen, Valencia, CA, USA). First-strand cDNA was synthesized using a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). The mRNA levels of alkaline phosphatase (Alpl), bone sialoprotein (Ibsp), osteocalcin (Bglap), Runx2, osterix, RANKL, and OPG were analyzed using the TaqMan gene expression assay (IDs: Alpl, Mm00475834_m1; Ibsp, Mm00492555_m1; Bglap, Mm03413826_mH; Runx2, Mm00501584_m1; osterix, Mm00504574_m1; RANKL, Mm00441906; OPG, Mm00435454_m1). Beta-actin (β-actin) (ID: Mm02619580_g1) was used as an internal control. The target mRNA levels were measured using a Thermal Cycler Dice Real Time System III (TaKaRa Bio). The mRNA levels in each sample were normalized to β-actin, and then compared with AA and β-GP-treated cells.
Statistical analysis
All assays were conducted in at least triplicate. Data are expressed as mean with standard deviation (SD). Statistical analyses comprised the analysis of variance (ANOVA) using Dunnett tests for multiple comparisons.
Results and discussion
Characterization of β-TCP and HAp particles
The surface morphologies of the particles, as determined by SEM, are shown in Fig. 1A. The crystallographic properties of the particles, as recorded by XRD, are shown in Fig. 1B. Estimated contents of doped silver on β-TCP (Ag) and HAp (Ag) were 2.7 and 2.3%, respectively. Ag could not identify in XRD profile. Since the silver content is small, the peak may not be clear. The median diameter, specific surface area and silver concentration of particles are summarized in Table 1. The median diameters of β-TCP, β-TCP (Ag), HAp and HAp (Ag) were 150, 360, 100 and 60–70 nm, respectively. The diameters of these particles were not very different. Only median diameter of HAp(Ag) was in nano-scale (10–100 nm). The specific surface areas of β-TCP, β-TCP (Ag), HAp and HAp (Ag) were 7.1, 6.8, 45.8 and 37.4 m2/g, respectively. HAp particles had larger specific surface area than β-TCP particles.
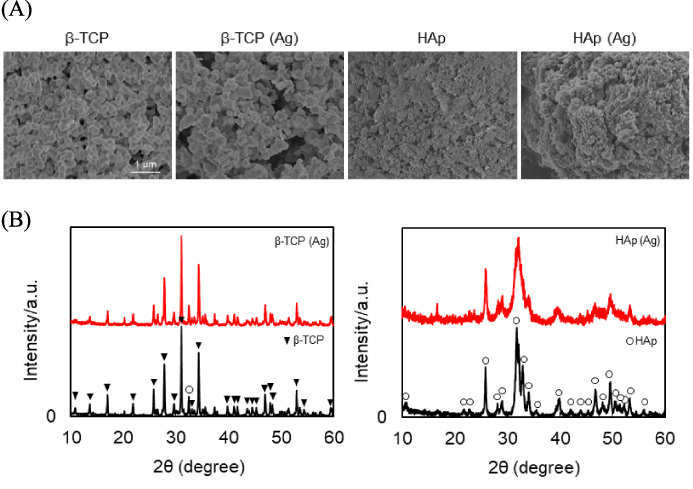
Characteristics of β-TCP and HAp particles used in this study. (A) shows SEM observations of β-TCP, Ag-doped β-TCP [β-TCP (Ag)], HAp and Ag-doped HAp [HAp (Ag)] particles. (B) shows X-ray diffraction (XRD) profiles for β-TCP, β-TCP (Ag), HAp and HAp (Ag) particles. β-TCP peaks are indicated by triangles (blacktriangledown) and HAp peaks by circle (circle).
Physical properties of particles used in this study
First, the cytotoxicity of β-TCP and HAp particles on MC3T3-E1 cells were examined. Cytotoxicity was assessed in terms of cell viability and cell membrane damage by WST-1 assay and LDH release, respectively (Fig. 2A and B). When the cells were treated with β-TCP or β-TCP (Ag) particles for 24 h, no cytotoxicity as assessed by WST-1 assay and LDH release was observed. On the other hand, treatment with HAp or HAp (Ag) particles resulted in a slight decrease in cell viability and an increase in cell membrane damage in a dose-dependent manner. These results were consistent with previous reports which stated that nano-scale HAp particles decreased cell viability and caused apoptotic cell death in certain cell types [26,27]. The elevation of cytosolic calcium ion concentration reportedly plays a crucial role in nano-scale HAp-mediated cytotoxicity [28]. Although it has been reported that particulate β-TCP and HAp enhanced the production of pro-inflammatory cytokines [16–19], the amounts of IL-6, TNF-𝛼 and IL-1β released from MC3T3-E1 cells treated with these particles in our study were below the detection limit (data not shown). Our results indicated that neither type of particle showed severe cytotoxicity against MC3T3-E1 cells up to 50 μg/mL.
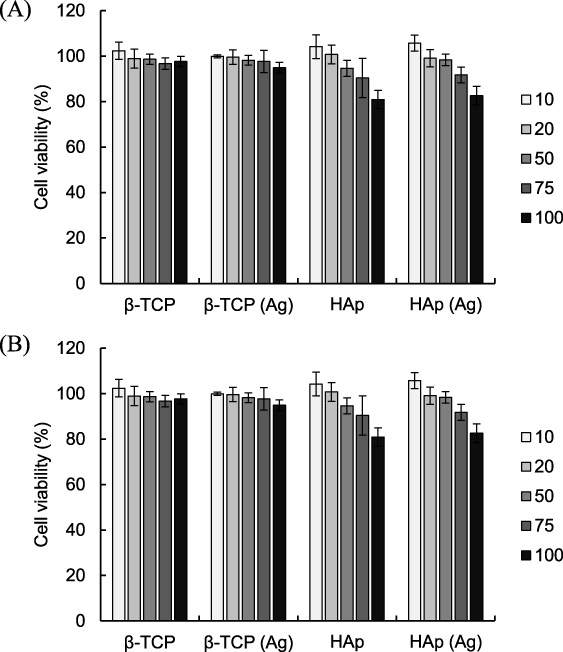
Effects of β-TCP and HAp particles on viability of MC3T3-E1 cells. (A) shows cell viability and (B) cell membrane damage as measured by WST-1 assay and LDH release assay respectively. Results are expressed as a percentage of untreated controls. Values are means ± SD (n = 3). ∗ P < 0.05, ∗∗ P < 0.01, compared with untreated controls.
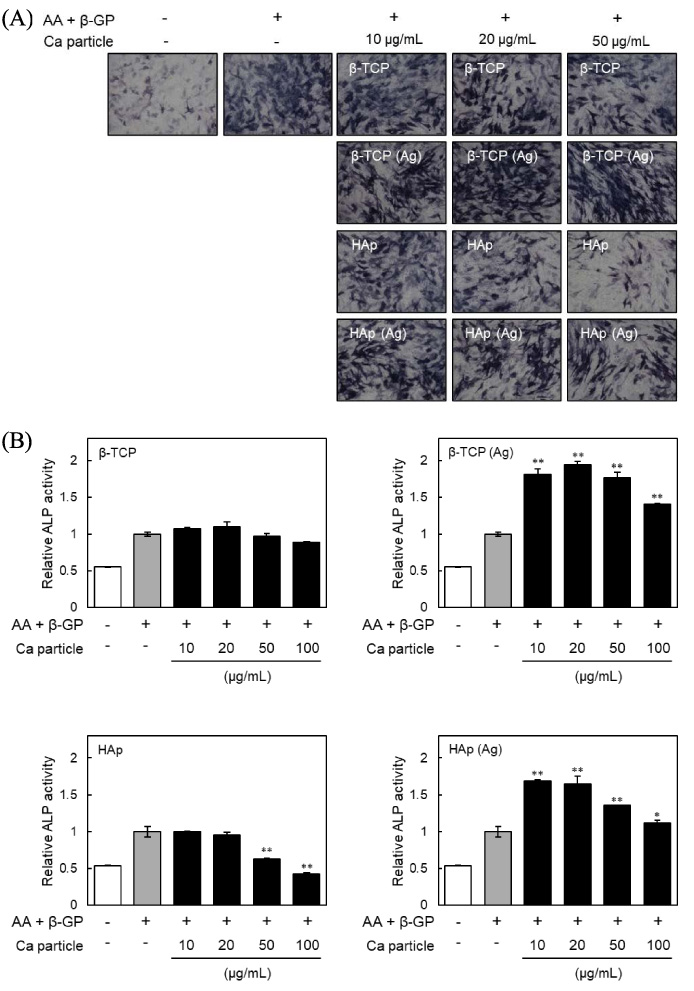
Effects of β-TCP and HAp particles on alkaline phosphatase (ALP) activity of MC3T3-E1 cells. (A) shows the ALP activity and (B) the ALP staining of MC3T3-E1 cells at day 7. Cells were incubated with β-TCP, Ag-doped β-TCP [β-TCP (Ag)], HAp and Ag-doped HAp [HAp (Ag)] particles for 24 h, washed with PBS three times to remove particles, and then cultured in the presence of ascorbic acid (AA) and β-glycerophosphate (β-GP) for an additional 7 days. The results are expressed as fold over the control cells, which were not treated with particles. Values are means ± SD (n = 3). ∗ P < 0.05, ∗∗ P < 0.01, compared with control cells.
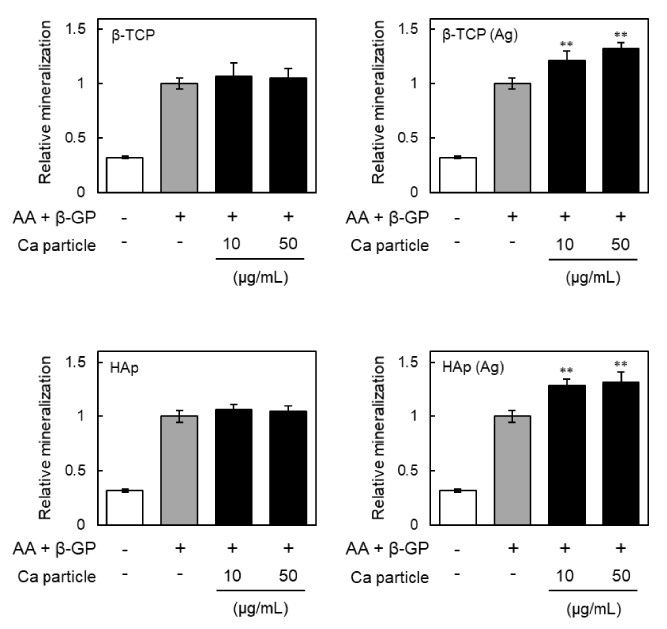
Effects of β-TCP and HAp particles on mineralization of MC3T3-E1 cells. Relative mineralization is shown for cells incubated with β-TCP, Ag-doped β-TCP [β-TCP (Ag)], HAp or Ag-doped HAp [HAp (Ag)] particles respectively for 24 h, washed with PBS three times to remove particles, and then cultured in the presence of ascorbic acid (AA) and β-glycerophosphate (β-GP) for an additional 28 days. The cells were stained with Alizarin red and the amount of dye was quantified by measuring at 562 nm. The results are expressed as fold over the control cells, which were not treated with particles. Values are means ± SD (n = 3). ∗∗ P < 0.01, compared with control cells.
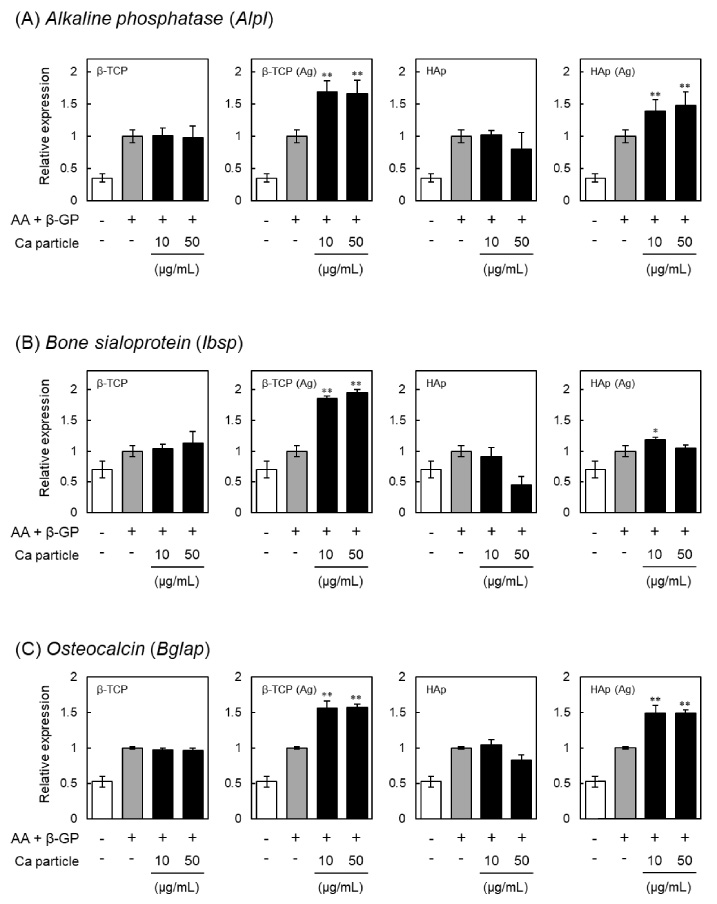
Effects of β-TCP and HAp particles on the expression of alkaline phosphatase (Alpl), bone sialoprotein (Ibsp) and osteocalcin (Bglap) in MC3T3-E1 cells. Relative expression of markers for osteogenesis is shown for cells incubated with β-TCP, Ag-doped β-TCP [β-TCP (Ag)], HAp or Ag-doped HAp [HAp (Ag)] particles respectively for 24 h, washed with PBS three times to remove particles, and then cultured in the presence of ascorbic acid (AA) and β-glycerophosphate (β-GP) for an additional 7 days. The mRNA levels shown in (A) Alpl, (B) Ibsp and (C) Bglap were measured using quantitative real-time PCR. Each mRNA level was normalized to the corresponding β-actin value and presented as fold over the control cells, which were not treated with particles. Values are means ± SD (n = 3). ∗ P < 0.05, ∗∗ P < 0.01, compared with control cells.
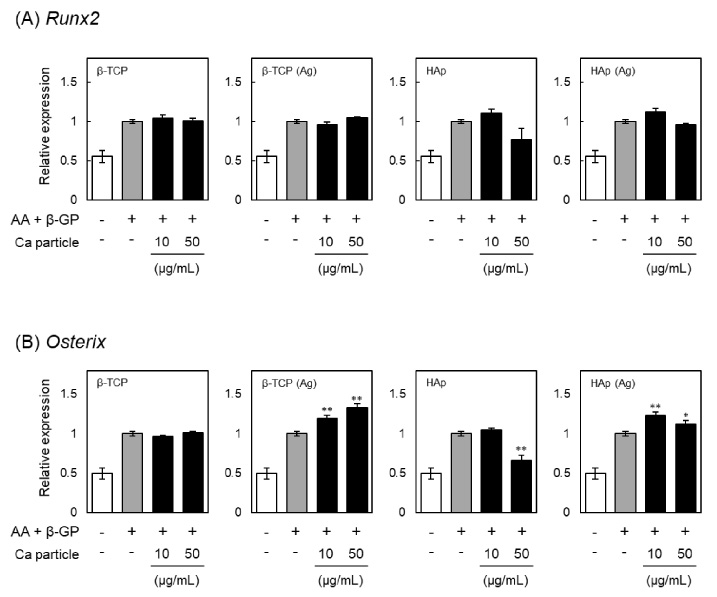
Effects of β-TCP and HAp particles on the expression of Runx2 and osterix in MC3T3-E1 cells. Relative expression of osteogenesis-related transcription factors is shown for cells incubated with β-TCP, Ag-doped β-TCP [β-TCP (Ag)], HAp or Ag-doped HAp [HAp (Ag)] particles respectively for 24 h, washed with PBS three times to remove particles, and then cultured in the presence of ascorbic acid (AA) and β-glycerophosphate (β-GP) for an additional 7 days. The mRNA levels of (A) Runx2 and (B) osterix were measured using quantitative real-time PCR. Each mRNA level was normalized to the corresponding β-actin value and presented as fold over the control cells, which were not treated with particles. Values are means ± SD (n = 3). ∗ P < 0.05, ∗∗ P < 0.01, compared with control cells.
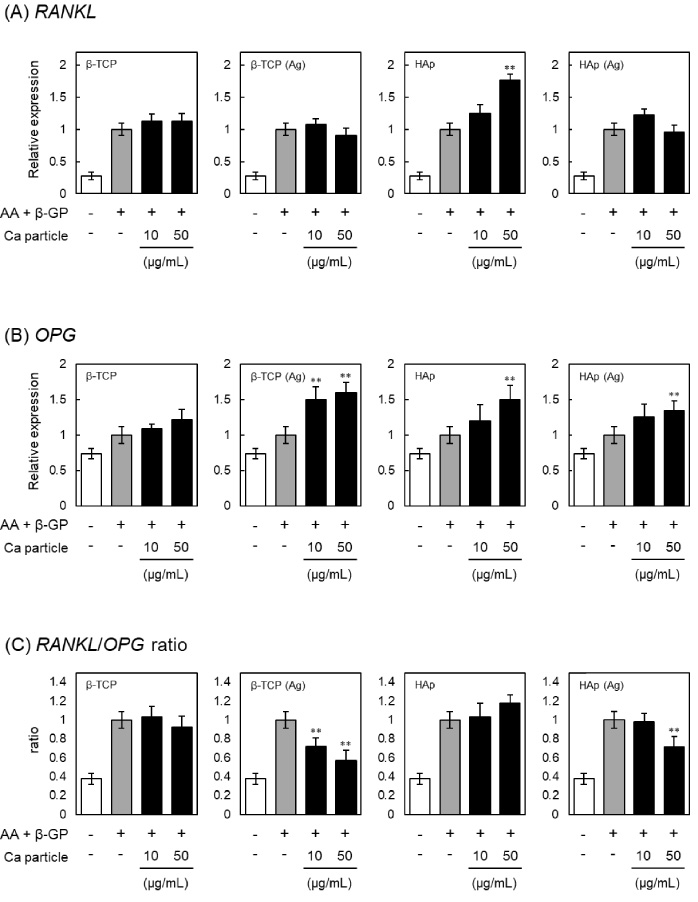
Effects of β-TCP and HAp particles on the expression of RANKL and OPG in MC3T3-E1 cells. Relative expression of osteoclast differentiation factor RANKL and its binding partner OPG is shown for cells incubated with β-TCP, Ag-doped β-TCP [β-TCP (Ag)], HAp or Ag-doped HAp [HAp (Ag)] particles respectively for 24 h, washed with PBS three times to remove particles, and then cultured in the presence of ascorbic acid (AA) and β-glycerophosphate (β-GP) for an additional 7 days. The mRNA levels of (A) RANKL and (B) OPG were measured using quantitative real-time PCR, and (C) the RANKL/OPG ratio calculated. Each mRNA level was normalized to the corresponding β-actin value and presented as fold over the control cells, which were not treated with particles. Values are means ± SD (n = 3). ∗∗ P < 0.01, compared with control cells.
For osteoblasts, ALP activity is considered as an early differentiation marker [29]. In addition, previous study showed that Ag nanoparticles significantly enhanced ALP activity and mineralization of MC3T3-E1 cells [10]. So to assess whether β-TCP (Ag) or HAp (Ag) particles are effective for the osteogenic differentiation of MC3T3-E1 cells, ALP activity in the cells after 7 days exposure to the particles was measured (Fig. 3). As shown in Fig. 3A, ALP staining to detect ALP activity revealed that treatment with β-TCP particles had no effect on the ALP activity, whereas treatment with HAp particles decreased the ALP activities of MC3T3-E1 cells. Previously, Ha et al. [30] demonstrated that nano-scale HAp particles stimulated suppression of ALP activity in a dose dependent manner, and that this effect was sustained for weeks. In contrast, the ALP activities of cells treated with β-TCP (Ag) or HAp (Ag) particles were significantly increased, indicating that Ag doping of β-TCP or HAp particles is effective in promoting osteogenic differentiation. As shown by the results of ALP staining, ALP activities were reduced by treatment with β-TCP or HAp particles, and were increased by treatment with β-TCP (Ag) or HAp (Ag) particles (Fig. 3B). As mineralization occurs during the latter stage of osteogenesis and is considered as a marker of osteogenic differentiation [31], we measured the mineralization of MC3T3-E1 cells after 28 days using Alizarin red staining. Mineralization by MC3T3-E1 cells treated with β-TCP (Ag) or HAp (Ag) particles was higher than for cells treated with β-TCP or HAp particles without Ag (Fig. 4). These results indicated that β-TCP (Ag) and HAp (Ag) particles increase ALP activity and promote the mineralization of MC3T3-E1 cells. The extent of silver elution from the particles may contribute to these activities. Therefore, the method of doping the calcium phosphate with silver can change its activity. However, the details of the relationship between the elution of silver ions and the manufacturing method of the particles are unknown at this time.
Expression levels of osteoblast differentiation-related genes
Bone sialoprotein is a non-collagenous component of the bone matrix and enhances the differentiation and mineralization of osteoblasts [32]. Osteocalcin is a calcium-binding protein found in bone extracellular matrix and expressed in osteoblasts [33]. Therefore, bone sialoprotein and osteocalcin are also considered as markers for an advanced stage of osteogenic differentiation [34]. We therefore investigated the expression levels of alkaline phosphatase (Alpl), bone sialoprotein (Ibsp) and osteocalcin (Bglap) in MC3T3-E1 cells treated with β-TCP or HAp particles, and the results are shown in Fig. 5. The gene expression levels of Alpl were significantly increased by treatment with β-TCP (Ag) or HAp (Ag) particles, as was also observed with regard to ALP activity (Fig. 2). Similarly, expression of Ibsp and Bglap was also induced by treatment with β-TCP (Ag) or HAp (Ag) particles. These results supported the view that β-TCP (Ag) and HAp (Ag) particles promote the osteogenic differentiation of MC3T3-E1 cells.
The genetic runt-related protein domain transcription factor-2 (Runx2) plays a pivotal role in the regulation of osteogenic differentiation. It has been reported that Runx2-deficient mice completely lack bone formation, which results in neonatal death [35]. In addition, zinc finger-containing transcription factor osterix is also required for osteogenesis and acts in the downstream of Runx2 [36]. Nakashima et al. [37] demonstrated that osterix-null mutant mice show normal cartilage development but a complete absence of bone formation. We therefore also investigated the expression levels of Runx2 and osterix in the MC3T3-E1 cells treated with β-TCP or HAp particles, and the results are shown in Fig. 6. Although the expression levels of Runx2 were not affected by any of the particles, those of osterix were increased by the treatment with β-TCP (Ag) or HAp (Ag) particles. Previously, it had been considered that osterix is a downstream target of Runx2 during osteoblast differentiation, because no osterix transcripts were detected in skeletal elements of Runx2-null mutant mice [37]. On the other hand, Lee et al. [38] reported that Runx2 is not involved in the induction of osterix. In addition, Matsubara et al. [39] showed that osterix is regulated via both Runx2-dependent and -independent mechanisms, and that it controls osteoblast differentiation by regulating gene expression. Considering these reports, our results suggested that the osteogenic differentiation promoted by β-TCP (Ag) or HAp (Ag) particles is mainly regulated by osterix-dependent but Runx2-independent pathways.
Effects of β-TCP (Ag) or HAp (Ag) particles on the RANKL/OPG pathway
RANKL and OPG secreted from osteoblasts are important molecules that participate in regulating bone remodeling [40]. RANKL stimulates osteoclastogenesis by binding to its receptor and is associated with the initiation of bone resorption [41]. OPG acts as a soluble receptor antagonist for RANKL and decreases osteoclastogenesis [42]. Finally, the mRNA levels of RANKL and OPG were further examined in the MC3T3-E1 cells treated with β-TCP or HAp. The mRNA levels of RANKL were increased by treatment with HAp particles, but not by treatment with β-TCP, β-TCP (Ag) and HAp (Ag) particles (Fig. 7A). On the other hand, OPG expression was increased by treatment with β-TCP (Ag), HAp and HAp (Ag) particles, but not by β-TCP particles (Fig. 7B). As a result, the ratio of mRNA expression was significantly decreased by treatment with β-TCP (Ag) or HAp (Ag) particles (Fig. 7C). The RANKL/OPG ratio in the bone microenvironmet is crucial for the differentiation and activity of osteoclasts, and down regulation of the ratio leads to a decrease in osteoclast maturation [43]. Therefore, our results suggested that β-TCP (Ag) or HAp (Ag) particles may have an indirect effect in regulating osteoclastogenesis via osteoblasts, through the RANKL/OPG pathway, predominately through their effect on OPG.
Conclusion
Particulate β-TCP and HAp doped with Ag promote osteogenic differentiation of MC3T3-E1 cells. In particular, the expression of osteoblast-related genes, Alpl, Ibsp and Bglap, was significantly increased by treatment with β-TCP (Ag) and HAp (Ag) particles. In addition, we found that treatment of MC3T3-E1 cells with β-TCP (Ag) and HAp (Ag) particles decreased the RANKL/OPG ratio, suggesting that Ag doping of β-TCP and HAp particles is effective for bone regeneration.
Footnotes
5.
None to report.