
Abstract
Introduction
Sepsis is a leading cause of death worldwide and its mortality rate could not be improved significantly, despite intense efforts to introduce novel treatment strategies [1]. Clinical studies provided many novel insights into the pathomechanism of sepsis and several potentially useful biomarkers have been suggested [2–4]. In septicemia, the activation of complement system as part of the inflammatory response together with the activation of coagulation system result in some predictive parameters for assessing the risk of lethal outcome [5]. A disadvantage of human studies is the large variability in the underlying clinical conditions as well as the effect of applied therapy on laboratory parameters.
The severity of the disease has been linked to several laboratory parameters in human studies, but the kinetics of these changes can better be followed when sepsis is elicited under controlled conditions in an animal experiment. A relatively well standardized way to study sepsis is to use an animal model that mostly replicates severe human sepsis. Unlike rodents, porcine sepsis models are indeed relatively close to human diseases and have been used in the past decade either by administering LPS or live bacteria to pigs [6, 7].
We studied hypoxia elicited markers after the administration of live Escherichia coli bacteria primarily investigating bone marrow responses in this lethal septic porcine model. Immature forms of all three lineages, red blood cells, platelets and leukocytes were studied. Furhermore, we hypothesized that mitochondrial membrane potential and an the antioxidant uric acid may be a suitable early marker for identifying hypoxemia induced changes during massive bacteremia. We suggest that the porcine septic model provides a crisp insight into the physiopathological changes during severe sepsis and may be suitable to determine the applicability of these tests in the course of human septic shockdevelopment.
Materials and methods
Porcine sepsis model
The experiments were carried in accordance with the European Community Guidelines and State Regulations with the approval of the University of Debrecen Committee of Animal Welfare (reg. Nr.: 21/2013. DEMAB). Seventeen juvenile female Hungahib pigs (bodyweight: 19.5±1.6 kg) were subjected into septic (n = 10) and control groups (n = 7). Under general anaesthesia (15 mg/kg ketamine, i.m. + 1 mg/kg xylazine i.m.) tracheostomy was performed for assisted ventilation, and the left external jugular vein and the femoral artery have been cannulated for sampling and hemodynamic measurements. The animals did not receive anticoagulant or any medication.
In the E.coli-treated group, sepsis was induced by Escherichia coli suspension (2.5×105/mL; strain: ATCC 25922, Department of Medical Microbiology, University of Debrecen), which was intravenously administered in a continuously increasing manner: 2 mL in the first 30 minutes, then 4 mL in 30 minutes and afterwards 16 mL/hour for 2 hours. A total amount of 9.5×106 E. coli was infused within 3 hours [8, 9]. In the control group the similar volume of isotonic saline solution was administered by the same protocol as in the septic group and no other intervention was applied. The anaesthesia was maintained for 4 hours and at the end of the experiment animals were euthanized.
Core temperature, hemodynamic parameters and blood gas monitoring
The core temperature was measured via the central venous cannula by PiCCO Monitoring Kit (Pulsion Medical Systems SE, Germany) connected to Philips IntelliVue monitor (Koninklijke Philips N.V., The Netherlands). Heart rate and mean arterial pressure were also monitored invasively by the PiCCO device. The modified shock index (MSI) − which is a descriptive parameter for the circulation status and a clinically significant predictor of mortality in emergency patients − was calculated as the ratio of heart rate and mean arterial pressure values [10]. The partial pressure of oxygen in arterial blood (paO2) and lactate concentration were recorded by GEM 3500 blood gas analyser (Instrumentation Laboratory, Bedford, USA).
Blood drawing and sample preparation
Immediately before E. coli administration blood samples were collected from the cannulated vein directly into blood collection tubes (Becton Dickinson, San Jose, CA, USA) containing 0.105M sodium-citrate as anticoagulant and the proportion of blood to the liquid anticoagulant volume was 9:1. Beside the baseline blood drawing, further blood samples were collected 2 and 4 hours after the start point of the infusion containing E. coli. All samples were transported to the laboratory at 37°C and the measurements were started within 20 minutes after blood drawing. Citrated whole blood was used for automated hematology analysis and blood smear examinations. During flow cytometric measurements, platelet rich plasma (PRP) was used which was obtained by centrifugation at 170 × g for 10 minutes at room temperature, then platelet count was determined by Sysmex XP 300 hematology analyser (Sysmex, Kobe, Japan) and it was adjusted to 250 G/L by platelet poor plasma (PPP). Clinical chemical tests were measured in PPP which was obtained by centrifugation at 1500 × g for 15 minutes at room temperature.
Hematology analysis and light microscopy
Blood cell count was performed according to our routine clinical laboratory procedure on ADVIA 120 hematology analyser (Siemens, Forcheim, Germany) using its special software suitable for porcine blood cell count. Nucleated red blood cells (NRBCs) were counted and platelet morphology was studied on peripherial blood smears using May-Grünwald and Giemsa staining. The microscopic examinations were carried out with Zeiss Axiostar Plus light microscope (Zeiss, Jena, Germany). The NRBC count (G/L) was calculated based on the NRBCs refered to 1000 white blood cells and the WBC count of the samples. Giant and vermiform platelets were quantified as the number of these platelets referred to 200 single platelets.
Flow cytometric measurements
Platelet mitochondrium membrane depolarisation was studied by FC500 flow cytometer (Beckman Coulter, Brea, CA, USA) based on a method [11] adapted to porcine platelets. The principle of this method is that platelets are stained with a fluorescent dye 3,3’-dihexil-oxocarbocianin-jodide (DiOC6 (3) ; Sigma-Aldrich, Budapest, Hungary), and when the mitochondrium membrane is depolarised, DiOC6 (3) accumulation in platelet mitochondria becomes impaired and a decreased mean fluorescence intensity (MFI) of the platelet population is measured. PRP aliquots (5μL) diluted 1:20 with phosphate buffered saline (PBS) were stained with 20μL of 1.5μmol/L DiOC6 (3) . Samples were incubated for 20 minutes at room temperature in the dark and were diluted to 560μL with PBS. Platelets were identified based on light scattering properties, and FL1 histograms were analysed. Mitochondrium membrane depolarisation was quantified as a decrease of MFI observed on FL1 histograms.
Clinical chemical tests
Uric acid and creatinine concentrations along with creatine kinase (CK) and glutamate pyruvate transaminase (GPT) activities were determined by COBAS 6000 analyser (Roche, Basel, Switzerland) according to routine clinical laboratory procedure.
Statistical analysis
GraphPad Prism 6.0 program was used for the statistical analysis. Data distribution was evaluated by Shapiro-Wilk test. The statistical significance of the differences between septic and control groups was determined by unpaired Student’s t-test in case of Gaussian distribution, and by Mann-Whitney test in case of non-Gaussian distribution. The matched septic animals’ data and the same of control data were analysed by ordinary one-way ANOVA using Bonferroni correction, or by Friedman test with Dunn’s correction, as appropriate. Differences were considered significant when p < 0.05. P values less than 0.05 are summarized with one asterisk, p≤0.01: **, p≤0.001: *** and p≤0.0001: ****. Data are expressed as median, interquartile range, minimum and maximum values of 10 septic animals and 7 controls.
Results
In the E. coli-treated group by 4 hours after the administration of bacteria, both the core temperature and the modified shock index parameters were increased significantly compared to the baseline data, while absolute lymphocyte counts and the lobularity index of the white blood cells decreased significantly (Fig. 1). In untreated control animals all of these parameters remained constant during the experiments.
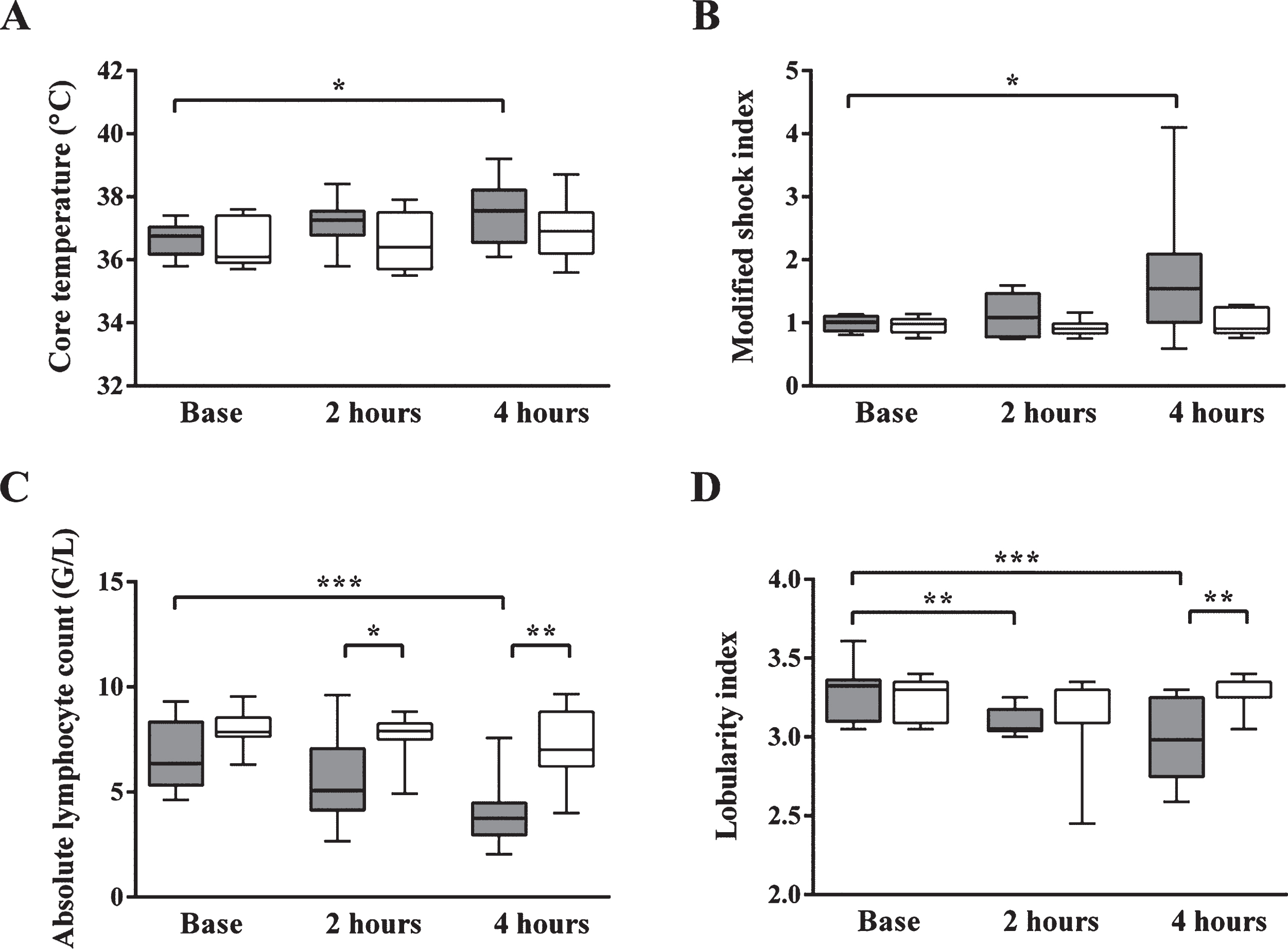
Core temperature (A), modified shock index (B), absolute lymphocyte count (C) and lobularity index (D) in case of septic animals (dark grey box) and controls (white box) before E. coli or physiological solution infusion, as well as 2 and 4 hours later. In the septic group by 4 hours after E. coli administration both the core temperature and the modified shock index parameters were increased significantly compared to the baseline data, while absolute lymphocyte counts and lobularity indices significantly decreased indicating the development of septic shock. In untreated control animals all parameters remained constant during the experiments. Data are expressed as median, interquartile range, minimum and maximum values of 10 septic animals and 7 controls. *p < 0.05, **p≤0.01, ***p≤0.001, ****p≤0.0001.
The surgical procedure (tracheostomy, assisted ventilation, arterial and venous cannulation) resulted in an acute phase response and muscle damage in both treated and untreated animals as reflected by the elevated neutrophil counts and CK activity values (Fig. 2). After E. coli administration, the absolute neutrophil count showed a mild, non-significant increase compared to the baseline data, the high maximum and low minimum values correspond to a considerable variance among septic animals. A significant increase was noted in control animals and the CK activities became elevated by the end of the experiments in both treated and untreated animals.

Trauma and stress elicited changes in animals. The surgical procedure resulted in an acute phase response and muscle damage in both treated and untreated animals as reflected by the elevated neutrophil counts (A) and CK activity values (B).
To investigate bone marrow reaction in sepsis, reticulocyte and platelet counts were measured by a hematology analyser, while NRBCs were counted and platelet morphology was studied on peripherial blood smears. The reticulocyte count and the NRBC count of the septic group were increased significantly compared to the control group and to the baseline values of septic animals (Fig. 3). These changes were significant as early as 2 hours after sepsis induction indicating a rapid bone marrow reaction in severe sepsis.
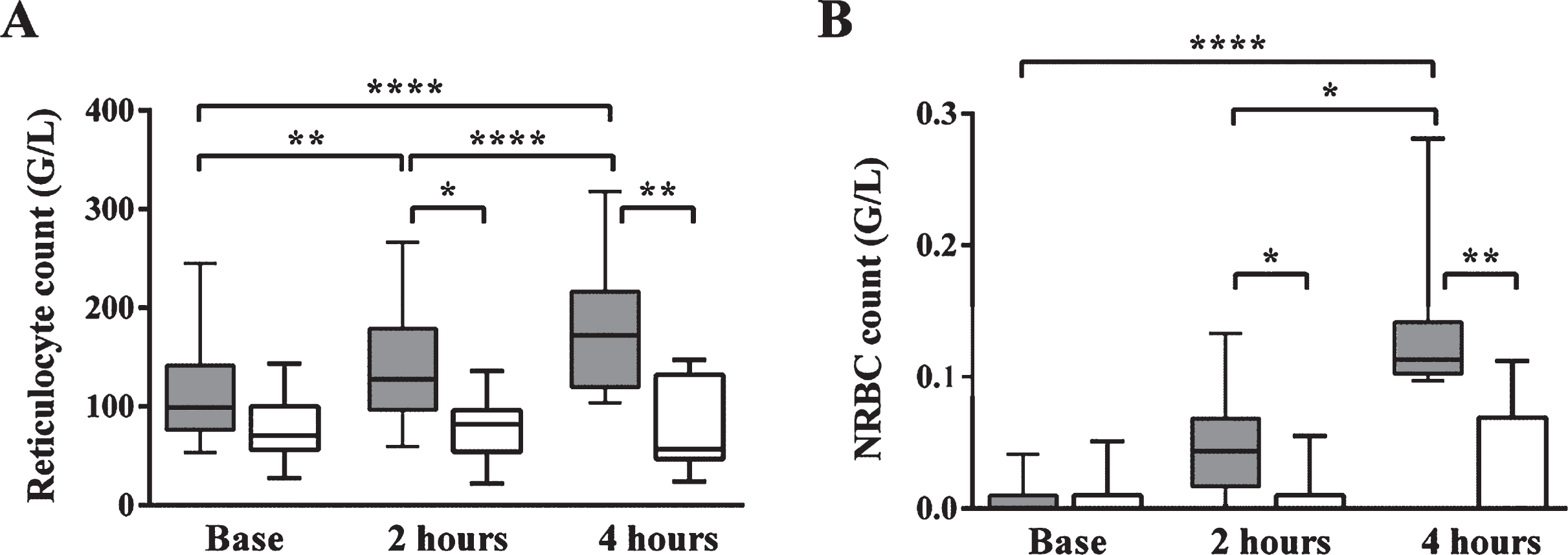
Absolute reticulocyte count (A) and nucleated red blood cell count (NRBC; B). The reticulocyte count and the nucleated red blood cell count of the septic group were increased significantly compared to the control group and to the septic animals’ baseline values. These changes were significant as early as 2 hours after sepsis induction.
Platelet count decreased significantly by 4 hours in both the septic and control group. This reduction was evident in septic animals already by 2 hours and further decreased up to 4 hours, corresponding to DIC after E. coli administration (data not shown). In controls a moderate platelet count decrease was detected. The platelet morphology in sepsis showed two types of alterations: heterogeneity in platelets possibly due to release from bone marrow (platelet anisocytosis, giant platelets and vermiform platelets), and platelet activation (platelet aggregates). In the E. coli-treated group an early and significant elevation of the number of giant and vermiform platelets was found compared to the control group and to the treated group’s baseline data (Fig. 4).
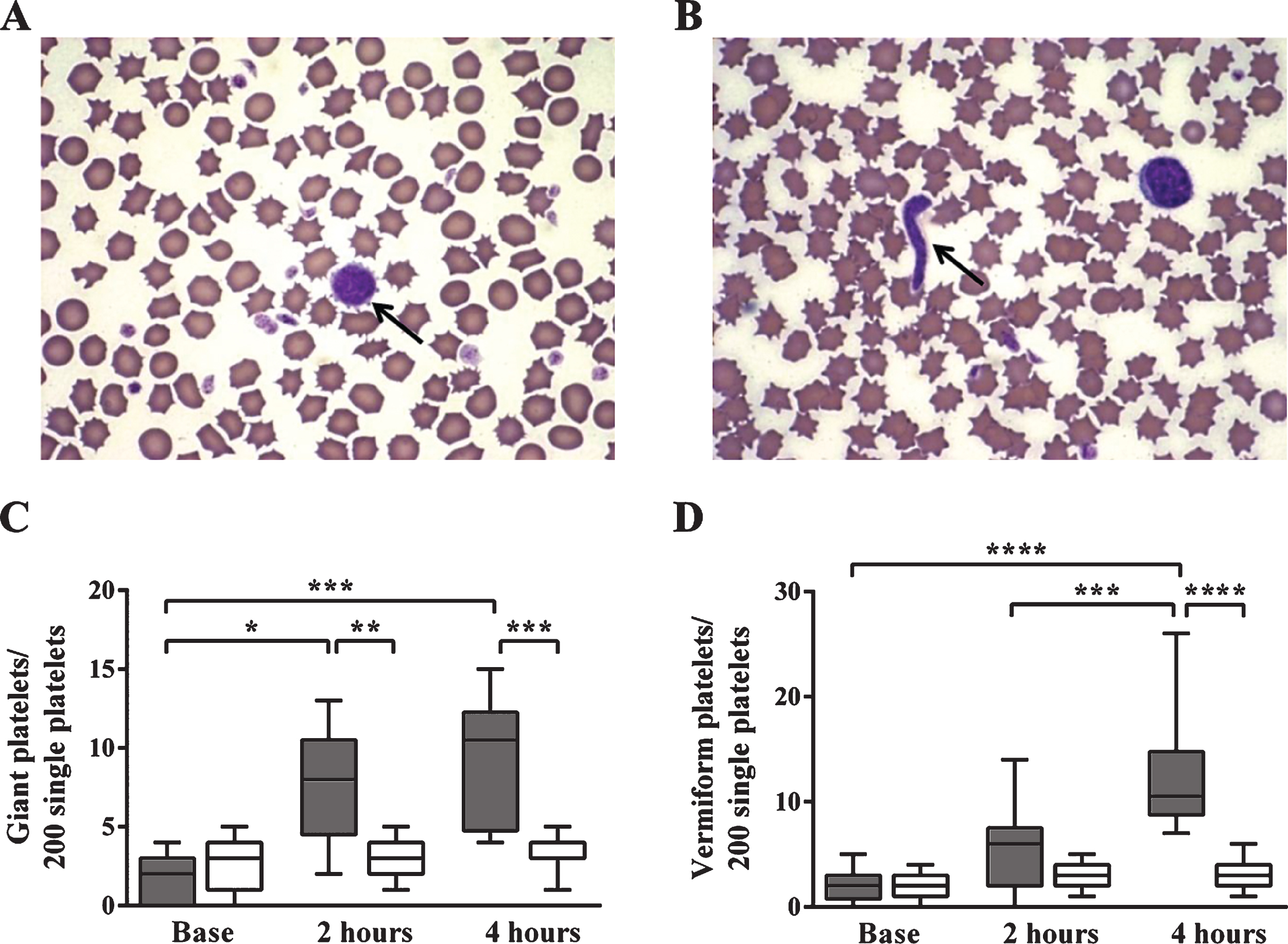
Platelet morphology in sepsis: giant (A, C) and vermiform (B, D) platelets. Platelet morphology in sepsis showed two types of alterations: heterogeneity in platelets possibly due to release from bone marrow (platelet anisocytosis, giant platelets and vermiform platelets), and platelet activation (platelet aggregates). Giant platelets (A) and vermiform platelets (B) are indicated by an arrow. The number of giant (C) and vermiform platelets (D) was shown referred to 200 single platelets. In septic animals an early and significant elevation of their number was found compared to the control group and to the baseline data of the septic group.
To follow the fate of platelets in sepsis, membrane depolarisation of platelet mitochondria was also determined. This was quantified as a decrease of MFI observed on FL1 histograms where the fluorescence signal for a mitochondria accumulated dye was detectable. The only significant change in the membrane depolarisation of platelet mitochondria was observed in E. coli-treated animals by 2 hours (Fig. 5). This result indicates enhanced mitochondrium membrane depolarisation and potentially platelet apoptosis in sepsis which occurs as early as 2 hours after sepsis induction.

Mitochondrium membrane depolarisation. Mitochondrium membrane depolarisation was quantified as a decrease of mean fluorescence intensity (MFI) observed on FL1 histograms. The only significant decrease in the membrane depolarisation of platelet mitochondria was observed in E. coli-treated animals by 2 hours.
The E. coli-treated animals developed marked hypoxia by 4 hours after sepsis inducing the corresponding appearance of NRBCs (Fig. 6). As a result of hypoxia, lactate acidosis occurred and the lactate concentration became significantly elevated by 4 hours into the range that in humans is regarded as severe sepsis. To monitor the response to oxidative stress, uric acid concentration was measured and was found to be continuously elevated. In untreated control animals all of these parameters remained almost constant during the experiments. These results indicate that uric acid concentration may be a more sensitive marker of oxidative stress than lactate concentration.
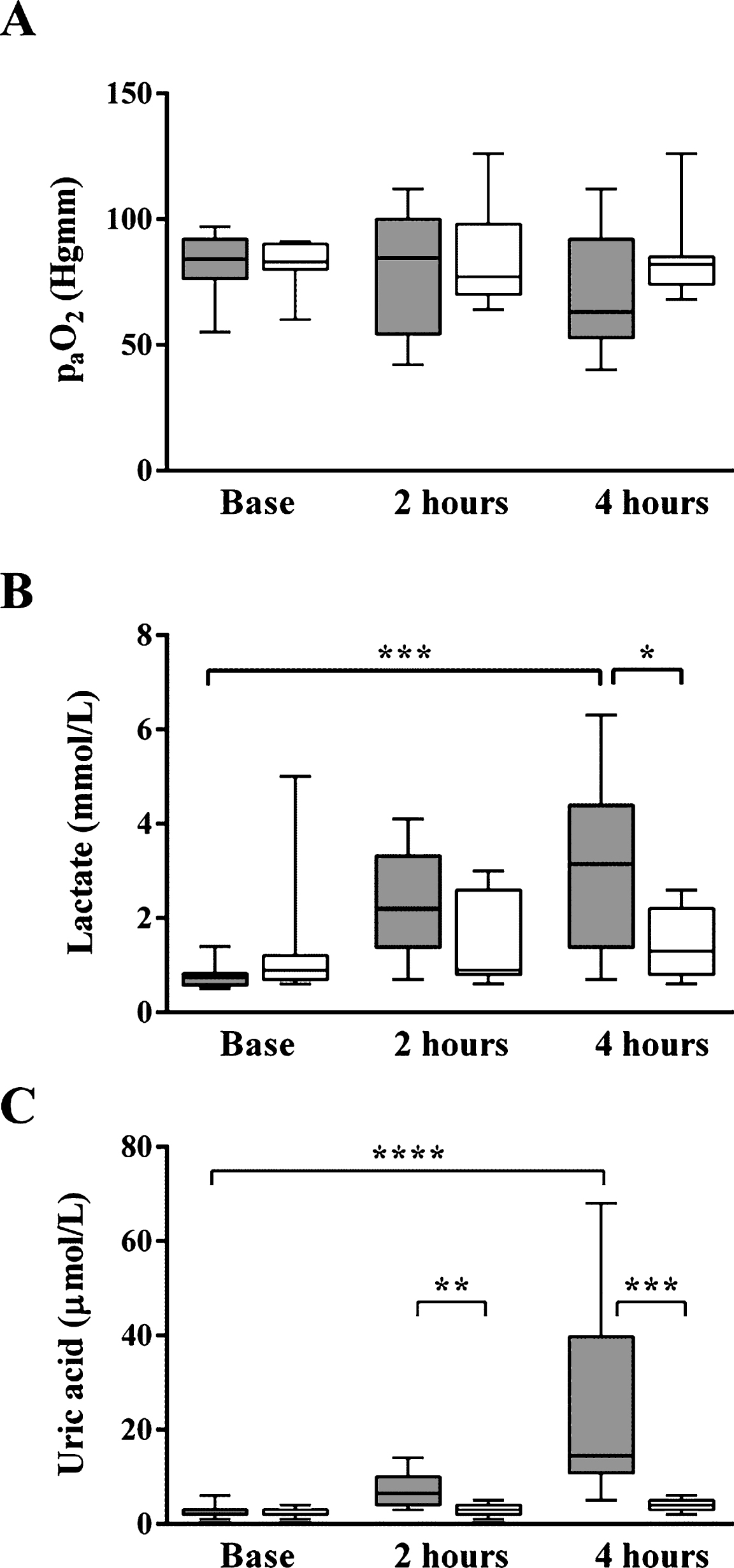
Oxidative stress markers. The E. coli-treated animals developed marked hypoxia by 4 hours after sepsis induction (A). As a result, lactate acidosis occurred and the lactate concentration became significantly elevated by 4 hours (B). To monitor the response to oxidative stress, uric acid concentration was measured and was found to be continuously elevated unlike in untreated animals (C).
To study potential renal and liver damage during the experiments creatinine values and GPT activities were measured (Fig. 7). Only creatinine values became significantly elevated by the end of the experiments, but this elevation was much milder than the changes observed in case of uric acid concentrations.

Markers for tissue damage. To study potential renal and liver damage during the experiments creatinine (A) and GPT values were measured (B). Only creatinine values became significantly elevated by the end of the experiments.
To characterize hypoxia elicited responses in septic patients standardized patient groups and controlled conditions for blood drawing are required. This is an uneasy task in a clinical setting due to the heterogeneity of the underlying disorder that may lead to severe sepsis. The majority of these patients develop multi-organ failure in the course of their disease that further complicates the picture. Finally, since nearly all patients receive antibiotics, anticoagulants, oxygen and many other drugs, objective observation of early changes during their disease development is hampered.
Bone marrow is known to react to intense bacteremia by releasing immature cells. In our septic cohort we could verify the decrease of lymphocyte count in septic animals versus controls by 2 hours as a part of sepsis criteria and the simultaneous appearance of left shift in neutrophils that is mirrored by the decrease of lobularity index. Both values kept progressing until 4 hours and these changes were not observed in the untreated animals.
In a previous report [9] we investigated the changes in microcirculation by laser Doppler tissue flowmetry and found that in septic animals after 1 hour over 40% reduction in blood flow was noticed that further progressed by 4 hours, while controls displayed only an insignificant decrease in flow by 1 hour that normalized in subsequent samples. We hypothesized that this blood flow impairment and resultant hypoxemia induce bone marrow reactions. Indeed, after 2 hours the absolute reticulocyte count increased and nucleated red blood cells appeared, mostly as polychromatic normoblasts with some orthochromatic cells. NRBC has been suggested in human studies previously as a marker that indicates mortality [12–14]. These authors concluded that NRBC may be considered as a parameter that sums up hypoxic and inflammatory changes during sepsis as in this condition hypoxemia occurs together with the increase of known stimulators of the bone marrow like erythropoietin, interleukin-3 and interleukin-6. Thus, the appearance of NRBCs in our study confirms previous reports and also identifies NRBC as an early marker of severe sepsis. Although reticulocyte count also increased as could be anticipated there the values of control and septic animals showed an overlap, while NRBC elevation was more specific for E. coli-treated animals.
Similarly to immature red blood cells, immature platelets were also released by the bone marrow. Previously the immature platelet fraction (IPF) was found to predict sepsis [3] and it was also found that the IPF is useful for discriminating septic patients from non-septic patients, but not for the determination of sepsis severity [15]. We investigated the morphological alterations in platelets that in our case were associated with the enhanced bone marrow response. We rarely observed giant platelets and vermiform platelets in smears of baseline samples and these two dysplastic forms increased several-fold only in the septic group while it did not change at all in the control group. The platelet is a very quickly reacting cell type in its standard function i.e. in hemostatic processes but it has been described that the platelet is also inflammatory cell [16]. Thrombocytopenia was identified as an independent variable significantly associated with increased mortality in a large prospective multicenter observational study [17]. The severity of thrombocytopenia parallels with the severity of inflammation and subsequent mortality [18]. The thrombocytopenia that accompanies the bacteremia also induces the release of young platelets. We thought to look at the mitochondrial function of these platelets that is important from the hypoxemia point of view. Mitochondrial alterations in pathological states can be harmful as well as beneficial for the host [19]. The consensus on the term ’mitochondrial dysfunction’ is still missing. Most studies that investigate this phenomenon require tissue homogenates and isolated mitochondria and the classical method is the measurement of oxygen consumption. Platelets are easily accessible cells and we determined the mitochondrial membrane depolarisation by a flow cytometric method that detects membrane leakage of platelet mitochondria measured by decline in the signal of a fluorescent dye. Mitochondrial membrane potential reflects mitochondrial function and is an indicator of mitochondrial energy status. Besides the production of energy, mitochondria play an important role in cellular survival and apoptotic death. Therefore, mitochondrial membrane depolarisation may reflect not only the abnormality of aerobic metabolism but it can be associated with the mitochondrial pathway of apoptosis. Although during our experiments a slow non-significant decrease was observed also in control animals, only the septic group displayed a significant drop in mitochondrial function by 2 hours indicating mitochondrial dysfunction and potentially platelet apoptosis in sepsis. The same technique was applied previously in humans and was found to correlate with the severity of the systemic inflammatory response [20]. Puskarich and coworkers have also published early alterations in platelet mitochondrial function in patients with septic shock which were associated with survival and organ failure [21]. Other apoptotic markers were investigated and was found to be useful in human studies. In severe sepsis in an observational clinical study, the serum caspase 3 activity was found to correlate with early mortality [22]. The mitochondrial dysfunction is known to be reversible, but this process is very sensitive even to mild hypoxia, as in our septic group the partial oxygen pressure was decreasing gradually but non-significantly during the experiment and the increase in the lactate values in our model became significant only after 4 hours.
Because of the different species and naturally dietary differences, uric acid may be an even better candidate for following hypoxemia induced changes in the septic pig model compared to human septic cases. Uric acid values are very high even in healthy humans the values are close to the solubility cut-off of this substance, but urate is exceedingly low in pigs. Thus, in human studies that utilized uric acid as a marker patients could only be dichotomized with ’normal’ or ’elevated’ urate values compared to sex matched reference ranges [23]. Unlike in humans, in our septic porcine model there is a huge difference by 4 hours in the uric acid levels of septic and control animals resulting in non-overlapping urate values. This increase is largely unrelated to kidney failure as the creatinine values of E. coli-treated animals only increase at 2–4 hours after the start of bacterial inoculation. It is also possible that uric acid itself can cause an acute kidney injury due to direct tubular toxicity, or crystal induced injury, or an indirect injury secondary to the release of vasoactive mediators and oxidative stress. During this sepsis model only a borderline creatinine elevation was observed with no change in the GPT activity i.e. the liver specific transaminase marker. Thus, it is important to emphasize, that urate elevation, mitochondrial membrane depolarisation and the bone marrow associated changes in red blood cells and platelets during this fulminant sepsis, all occur prior to the occurence of evident multi-organ failure.
The study has some limitations, as the number of investigated animals in each group is relatively small, but all animals were females and of similar age and the observed differences seemed convincing. Histopathological evaluation of tissues has also not been carried out in these animals, as we focused on the kinetics of biomarkers that may help to detect early changes during fulminant sepsis. We think that further studies are required to confirm the usefulness of these data in severe human sepsis.
Conflict of interest
The authors declare that there is no conflict of interest.
Author contributions
BF, NN and JK designed and coordinated the study, participated in the interpretation of the data. JT and IBD made the laboratory measurements, analysed the data and participated in the interpretation of results. JT performed the statistical analysis. MB, EH, ÁD and KP participated in anaesthesia and sepsis induction of the animals, point-of-care testing and blood drawing for laboratory measurements. JSz provided Escherichia coli for the experiments. JT and JK drafted the manuscript.
All authors revised the manuscript critically for important intellectual content and made the final approval of the version to be published.
Footnotes
Acknowledgments
Authors are grateful to the technical and laboratory staff of the Department of Laboratory Medicine and the Department of Operative Techniques and Surgical Research at University of Debrecen. The project was supported by the Hungarian Brain Research Program - Grant 510 No. KTIA 13NAP A II/5.