
Abstract
In cancer therapy, a number of drugs with different mechanisms of action are in clinical use, which act directly after administration without metabolism, while others only become active in the metabolites produced in the liver. Such drugs/metabolites – especially when administered parenterally – interact in high concentrations with the endothelium. Whether this induces adverse responses of the endothelial cells (EC) is barely studied for many medicaments.
This pilot in vitro study revealed that the addition of cyclophosphamide (CPA) to the culture medium (5 or 10 mM, respectively) showed a clear influence on EC compared to non-treated EC: The number of adherent human vein endothelial cells (HUVEC) decreased by the addition of CPA in a concentration-dependent manner compared to the untreated control, whereby the vitality of adherent cells was not affected. In addition, concomitant with activation of the adherent HUVEC, increased migratory activity occurred.
These results are in agreement with clinical events like thromboses in patients in compromised condition under therapy with CPA, as the detachment of EC might induce responses of circulating platelets leading to the adherence and aggregation with the risk of the formation of thrombi. Whether CPA acts directly or via toxic metabolites on EC will be examined in more detail in following studies.
Introduction
A lot of different anticancer drugs with different mechanisms of action are in clinical use. Some of these drugs act directly after administration without metabolism, while others only become active in the metabolites produced in the liver (first pass effect).
The alkylating cytostatic cyclophosphamide (CPA) is a chiral compound with two stereoisomers, the (+)-enantiomer and a (–)-enantiomer, because of the asymmetrically substituted phosphorus (Fig. 1). The anticancer drug CPA (RS) -2- [bis (2-chloroethyl) amino] -1,3,2-oxazaphosphorinane-2-oxide; (RS) -2- [bis (2-chloroethyl) amino] -2H-1,3,2-oxazaphosphinane-2-oxide; cyclophosphamide) is described to become effective only after biotransformation in the liver. CPA is therefore a prodrug, which develops its cytotoxic properties only after activation in the liver [1, 2]. In liver cells, it is hydroxylated by cytochrome P450 oxidases to 4-hydroxycyclophosphamide-aldophosphamide (4-OHCP). The latter is cleaved to acrolein (a toxic ingredient also in food [3]) and becomes phosphoramide mustard, the active metabolite with the alkylating potency [2, 5]. Metabolism in the liver is carried out by various cytochrome P450 (CYP) enzymes, e.g. CYP2A6, CYP2B6, CYP3A4/5, CYP2C8, 2C9, 2C18 and 2C19 [1], according to Clarke & Waxman are these CYP2B1, CYP2C6 and CYP2Cll [6] and according to Schwab et al. CYP1B6 and CYP3A4/5 [7].
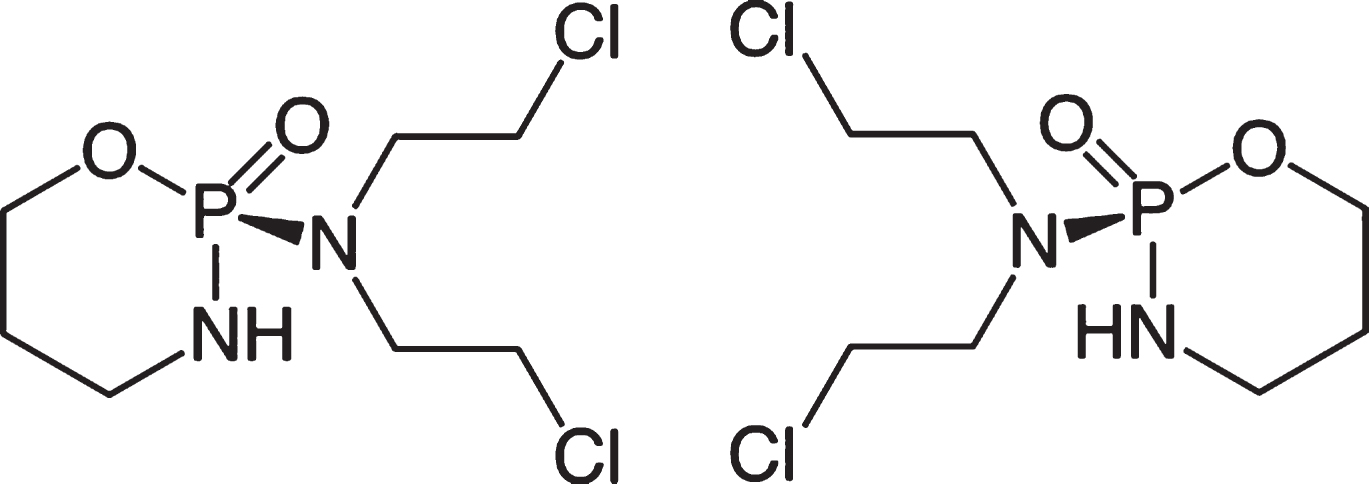
Structural formula of Cyclophosphamides (C7H15Cl2N2O2P·H2O).
Phosphoramide mustard causes alkylation of the DNA, which in turn leads to crosslinking of DNA strands and abnormal or abrogated base pairing [8]. This results in impaired cell division, which is well recognizable in the side effect profile of this substance (inter alia: inhibition of bone marrow function, deficiency of all blood cells (neutrophils, granulocytes, platelets, erythrocytes), inflammation, bleeding, fever, etc.).
CPA is administered orally as well as parenterally. Therefore, locally high concentrations in the vasculature can be achieved, possibly inducing damage to EC in the infusion area. Since endothelial cells can not metabolise CPA, in vitro testing in endothelial monoculture is an appropriate methological approach for exploring whether CPA as a prodrug exerts a negative influence on endothelial cells. This should be checked as part of a small pilot study.
Cultivation and treatment of HUVEC
Human umbilical vein endothelial cells (HUVEC, Lonza, Cologne, Germany) were cultivated in EBM-2 supplemented with EGM-2 singleQuots kit and 2 vol% FCS (Lonza, Cologne, Germany). Cells were used no longer than 3 passages [9]. For cell analysis cells were seeded in 24-well plates (TPP, Techno Plastic Products AG, Trasadingen, Switzerland) covered with glass slides with a cell density of 20,000 cells/well. Subsequently, CPA was added to the cell culture medium in concentrations of 5 mM or 10 mM. As (negative) control cells were cultivated in pure EGM-2 medium. As a further control HUVEC were used, which were treated with IL-1β (as supplement to EGM-2, leading to an activation of the endothelial cells). Every second day the cell culture medium was changed and all supplements were added freshly.
Experimental procedure
24 h after HUVEC seeding CPA was added in two concentrations directly to the cell culture medium. Then the cells were cultured in maximum for eight days, in order to investigate a broad range of the cell aging. The detailed procedure is presented in Table 1.
Design of the in vitro study
Design of the in vitro study
The density of adherent endothelial cells per mm2, the percentage of vital adherent endothelial cells (fluorescein diacetate / propidium iodide staining), mitochondrial activity by MTS assay, and the integrity of the cell membrane by lactate dehydrogenase (LDH) release were measured.
The analysis of the cell viability was performed using fluorescein diacetate (FDA, 25μg·mL–1, Invitrogen, Carlsbad, CA, USA) to stain vital cells in green and propium iodide (PI, 2μg·mL–1, Sigma, Taufkirchen, Germany) to stain dead cells in red. Subsequently, three pictures per sample (n = 3-4) were taken in 10-fold magnification (LSM 510 META, Zeiss, Oberkochen, Germany). HUVEC were stained 2, 6, and 9 days after seeding.
Measurement of cell membrane integrity
Cell membrane integrity was measured using the LDH Cytotoxicity Assay Kit II (LDH, Roche, Grenzch, Germany) according to the manufacturer’s instructions 2, 6, and 9 days after seeding.
Measurement of metabolic activity
The mitochondrial activity was measured using MTS cell Titer 96® Aqueous Non-radioactive Cell Proliferation Assay (MTS assay, Promega, Mannheim, Germany) according to the manufacturer’s instructions.
Statistics
Data are reported as arithmetic mean±standard deviation for continuous variables. For two-sample problems a two-tailed t-Test for paired samples was used, in the case of three sample problems an one-factorial ANOVA with post-hoc analyses. A p value of less than 0.05 was considered significant.
Results
Density of adherent HUVEC
While the density of the HUVEC increased as expected from 846±122 HUVEC / mm2 to 1094±103 HUVEC / mm2 8 days after cell seeding, the density of adherent HUVEC was significantly reduced by the addition of CPA compared to the untreated control (Fig. 2). HUVEC density decreased to 370±41 HUVEC / mm2 after addition of 5 mM CPA after 8 days of incubation, and to 274±8 HUVEC / mm2 after addition of 10 mM. The decrease after 10 mM CPA addition was greater at all times than after 5 mM CPA (after 2 days: p < 0.05, after 6 days: p < 0.05, after 8 days: p < 0.01).

Density of adherent HUVEC in [cells/mm2] 2, 6, and 8 days after adding CPA in comparison to control cells or cells activated with IL-1β (Arithmetic mean±standard deviation). *: p < 0.05 versus HUVEC, **: p < 0.01 versus HUVEC, ***: p < 0.001 versus HUVEC; +: p < 0.05 versus HUVEC+5 mM CPA, ++: p < 0.01 versus HUVEC+5 mM CPA.
Figure 3 shows the percentage of vital adherent HUVEC for the four groups (control cultures, cultures supplemented with 5 mM and 10 mM CPA, respectively, and cultures supplemented with IL-1β).
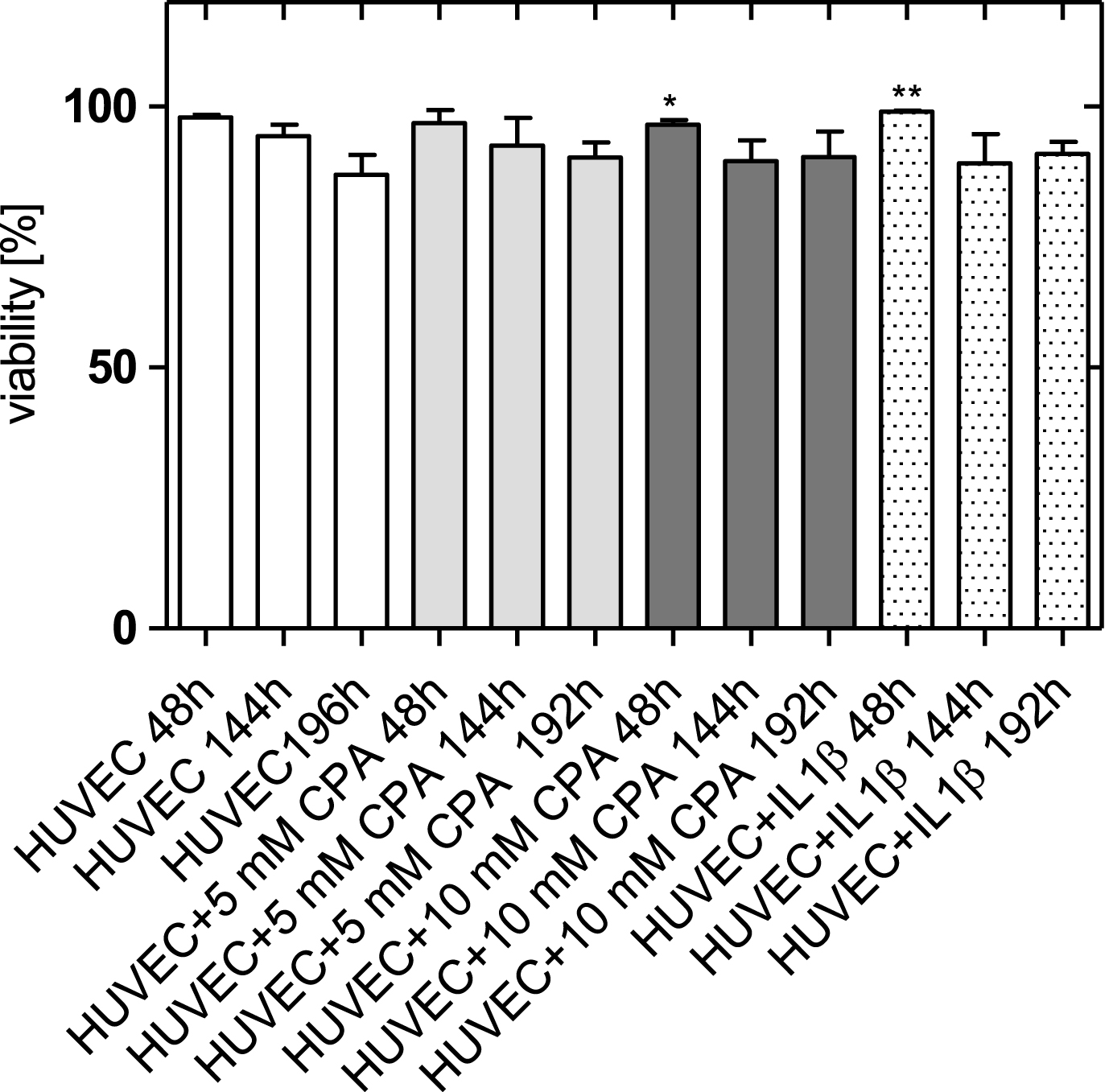
Viability of adherent HUVEC 2, 6, and 8 days after adding CPA (arithmetic mean±standard deviation) in comparison to control cells and cells activated with IL-1β. *: p < 0.05 versus HUVEC, **: p < 0.01 versus HUVEC.
The viability of the adherent HUVEC supplemented with CPA was nearly comparable to control cells. Two days after CPA addition, the number of adherent endothelial cells was slightly reduced after addition of 10 mM CPA (97±1 %) compared to the control (98±1 %).
Figure 4 shows the endothelial cell monolayer for the four groups 4 to 9 days after cell seeding. Vital endothelial cells are marked green, dead cells red. However, there are so few dead HUVEC that they only become visible at the highest post-enlargement. Clearly recognizable, the number of adherent endothelial cells remained significantly reduced after the addition of both concentrations of CPA until the ninth day. While almost all HUVEC in the control cultures on the ninth day had a round, spread shape, this is visibly not the case after CPA addition. Here, the cells are significantly elongated especially after 10 mM CPA addition presented with a high migratory activity. However, there were first islets of contacting cells with gradual spread. Overall, CPA seemed to lead to a pronounced activation of HUVEC in a more attenuated form and clearly 4 days compared to cells after stimulation with IL-1β.
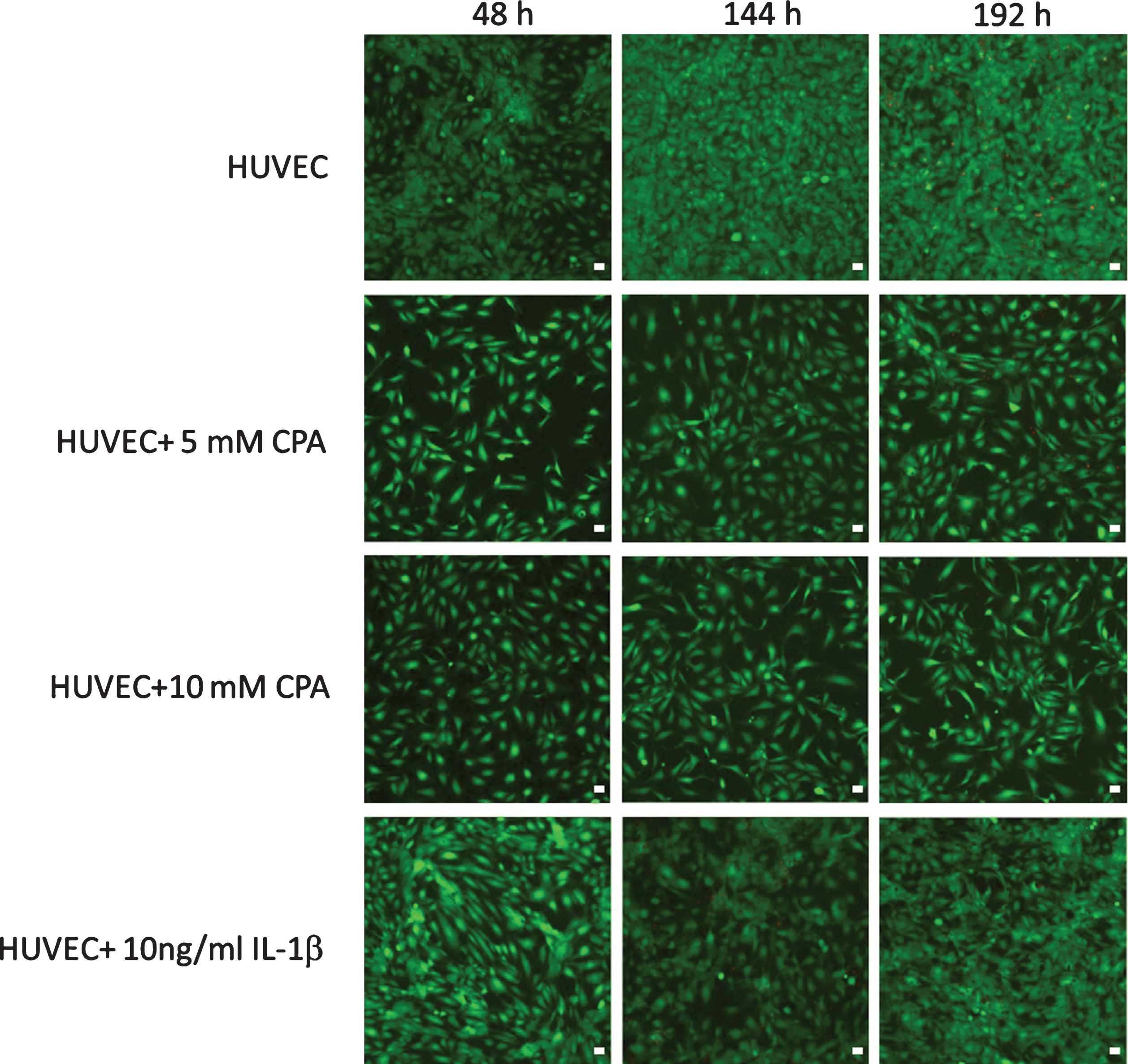
HUVEC monolayer 2, 6, and 8 days after adding CPA (5 mM or 10 mM, respectively) in comparison to control cells and cells activated with IL-1β. Viable cells are presented in green (fluorescein diacetate staining), dead cells are presented in red (propium iodide). 10× primary magnification using confocal laser scanning microscopy, bar: 50μm.
There was a significant influence of CPA on the metabolic activity of the HUVEC (see Fig. 5). In control cells a clear signal increase occurred, while after adding CPA a dose-dependent decrease of the metabolic activity was found.

Metabolic activity of HUVEC 2, 6, and 8 after CPA addition in comparison to control cells or cells activated with IL-1β (arithmetic mean±standard deviation). ***: p < 0.001 versus HUVEC, +++: p < 0.001 versus 5 mM CPA.
LDH release was highest in the control (Fig. 6). Presumably, this was due to the fact that an increased detachment of cells in the supernatant occurred, since the cell number increased most significantly here. As detached HUVEC do not survive in the supernatant, it can be assumed that the dead cells released lactate dehydrogenase.
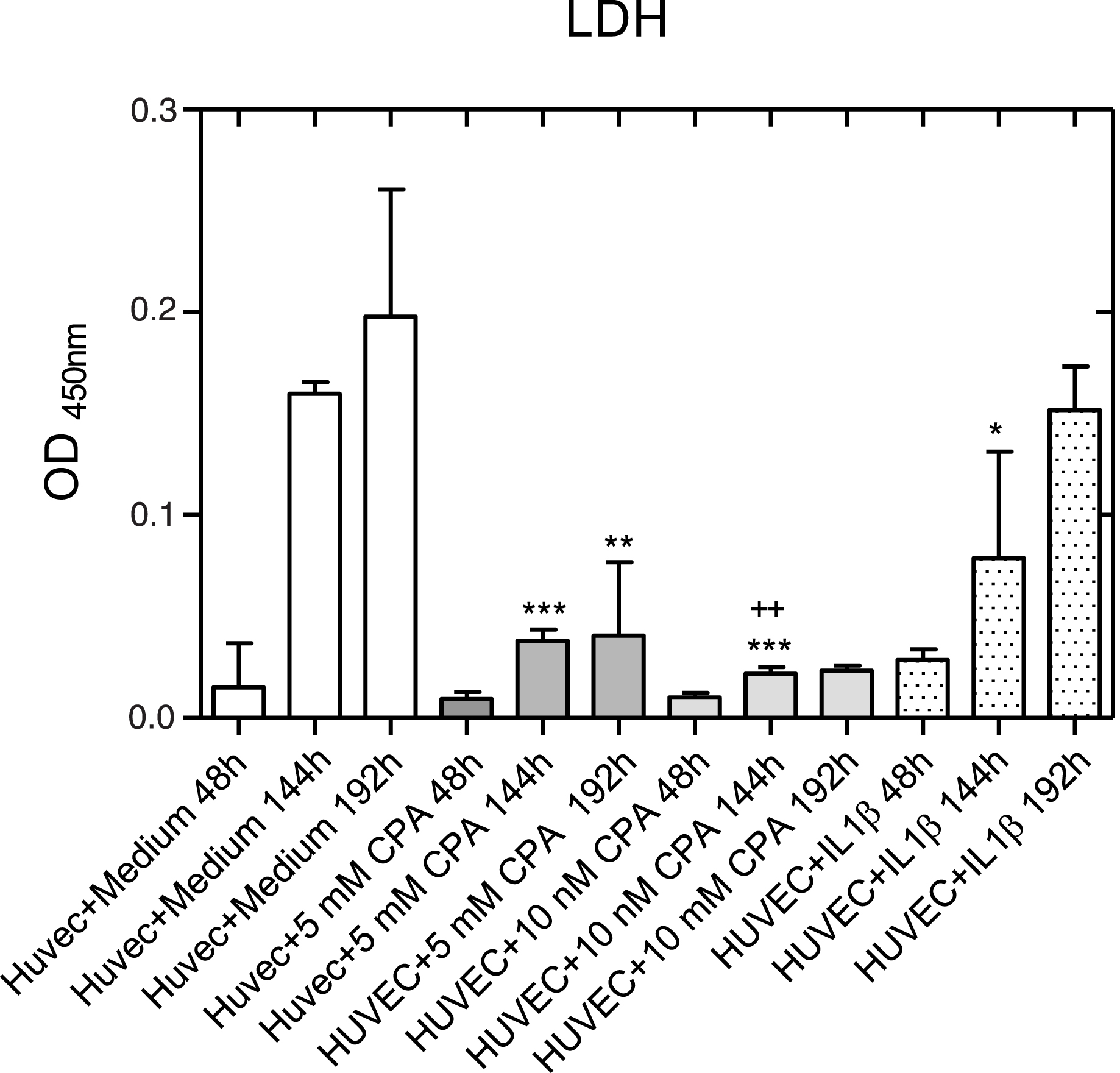
Integrity of the membrane of adherent endothelial cells on days 2, 6, and 8 after CPA addition in comparison to control cells or cells activated with IL-1β (arithmetic mean±standard deviation). *: p < 0.05 versus HUVEC, **: p < 0.01 versus HUVEC, ***: p < 0.001 versus HUVEC, ++: p < 0.01 versus 5 mM CPA.
Cyclophosphamide had a marked and significant influence on human vein endothelial cells: The number of HUVEC per area decreased in a dose-dependent manner after adding HUVEC to the culture medium (5 or 10 mM, respectively) in comparison to control cultures. The viability of the adherent HUVEC was not influenced and did not differ between the four groups. In addition, concomitant with the activation of the HUVEC monolayer, increased migratory activity occurred.
At a first glance, one might conclude that CPA already acts as a prodrug on HUVEC. But it is also conceivable that CYP enzymes endogenously expressed in endothelial cells might cause production of cytostatic CPA metabolites. It is certain that endothelial cells have a CYP system, as they are e.g. able to metabolize arachidonic acid from omega 3 fatty acids. They thus clearly have CYP450 enzymes that could in principle metabolize CPA (such as CYP2C9 [10]). In addition, human endothelial cells are known to express also CYP2C8, 2C9, 2J2, 3A, and 2B1 enzymes [10, 11]. Whether they metabolize xenobiotics, is still unclear and remains to be explored. Not only parenchymatous cells are involved in the liver, but also non-parenchymatous cells such as endothelial cells are involved in the metabolism of drugs [12]. Very recently it was reported that human capillary endothelial cells of patients with epilepsy possess a nuclear receptor, the pregnane X receptor, which could be responsible for a local drug metabolism by endothelial cells [13].
Since endothelial cells differ morphologically and functionally, depending on the localization in a vessel as well between organs, it is unclear whether the entire endothelium is metabolically active. Due to its size and water solubility, CPA is unable to pass cell membranes. 70% of CPA are rapidly metabolized to 4-hydroxycyclophosphamide (4-OHCP) [14], 25% are excreted unchanged [14]. 4-OHCP can pass cell membranes and enter the cell interior [15], where it is then cleaved into phosphoramide mustard (the ultimate genotoxic metabolite) and acrolein, which is also toxic [16].
In cell culture, the ability of CYP expression of endothelial cells decreases within hours [17], so that the results obtained could be due to CPA itself acting as a toxic prodrug.
Interestingly, the plasma levels of CPA differ significantly in oral and parenteral administration, but not those of the active metabolite [18]. These authors showed that while the AUC (area under the curve) of CPA differs significantly according to the mode of administration (i.v.: 7.14±3.77μg/h/ml, oral: 1.85±2.58μg/h/ml; p < 0.05), this does not apply to the values of the active metabolite 4-OHCP (iv: 1.66±0.36μg/h/ml, oral: 1.42±0.64; ns). These data also indicate that most of the CPA biotransformation takes place in the liver. If the endothelial cells were to contribute to the metabolism of CPA, the significantly higher plasma concentration of CPA when administered parenterally should also lead to higher plasma levels of the active metabolite, but this is not the case. A toxic effect of CPA as a prodrug on endothelial cells would be a new finding that could possibly have an impact on the application of the therapeutic in the clinic.
Clinical implication
In approximately 15% of cancer patients thromboembolic events can be expected, in patients with bronchial carcinoma up to 28% [18–20]. The exact cause is mostly unknown. In fact, the percentage of thromboembolic events in cancer patients is significantly higher; postmortem examinations in 1505 autopsies with malignant disease showed either venous thrombosis, pulmonary embolism, or both (40.3%) in 607 of the deceased [20].
From a pathophysiological point of view, the Virchowian triad naturally also applies to malignant diseases. In tumor patients, obviously increased procoagulant conditions also contribute to the high rate of thrombosis. The in vitro preliminary experiments of this pilot study suggest that CPA may contribute to this. CPA led to a considerable influence on the endothelial cell monolayer in cell culture: there was a significant and relevant reduction of adherent endothelial cells (as has already been described in vivo by the detection of shed endothelial cells for a number of cytostatics [21]). In addition, initial clinical studies on endothelial dysfunction (flow mediated dilatation [22]) revealed marked disturbances of the dilation capacity of the brachial artery under various chemotherapies [23, 24].
However, even the detachment of endothelial cells would be prothrombotic in vivo, because in this case the exposed subendothelium and the adhesive potential of platelets becomes evident, then performing their main function, which is primary hemostasis. This process involves the very rapid adhesion of platelets to the exposed subendothelium followed by platelet-to-platelet adherence. This ultimately culminates in the formation of a platelet plug, which temporarily seals the damaged vessel wall [25]. In the case the surrounding endothelium is activated releasing prothrombotic mediators (such as tissue factor, endothelin, thromboxane, ROS, thrombospondin-1, etc.), the risk of thrombus formation is markedly enhanced. The “worst case scenario” would then be the metabolic effect of the activated vascular endothelium, which provides prothrombotic metabolites on-site that could sustain and enhance this prothrombotic state. In this case, 30–50% thrombocytosis [26], which promotes thrombus formation due to the significantly increased presence of thrombocytes, especially hyperaggregable platelets [27], must be considered.
Catheter-mediated endothelial damage in central venous administration can also trigger such a condition. With concomitant chemotherapy, especially high-dose chemotherapy, the incidence of thromboembolic events is disproportionally higher [28].
Some of these factors speak therefore rather for the oral than for the parenteral application; i) no catheter injury that could be the source of thrombus formation; ii) no high intravascular CPA concentration that would result in endothelial cell detachment with activation of the endothelium.
Limitations of the study
First of all, the sample size of the study was too low to generalize the results. Furthermore, the activation of HUVEC after exposition with CPA has to be confirmed by examining e.g. E-selectin, ICAM-1, VCAM-1, MCP-1, IL-1 or IL-8, PAI-1 or tissue factor in further studies. In addition, the mechanism of endothelial cell detachment would have to be investigated. There are indications that this could occur via a reduction in proliferation, or an induction of apoptosis. At the same time, it must be investigated whether cell-cell or cell-substrate binding may also be resolved, and whether submembrane actin filaments or intermediate filaments may be affected.
Last but not least, it has to be analyzed whether the formation of toxic metabolites due to metabolism in HUVEC occurs, as this has very recently been shown in human capillary endothelial cells of patients with epilepsy [13].
Conclusion
The study revealed that CPA in vitro induced the detachment of HUVEC together with a disruption of the integrity of the plasmatic membrane compared to non-treated cells.
Footnotes
Acknowledgments
The work was supported by the Ministry for Science, Research and Cultural Affairs of Brandenburg through the grant of the joint project “Konsequenzen der altersassoziierten Zell- und Organfunktionen” of the Gesundheitscampus Brandenburg and by the Helmholtz-Association through programme-oriented funding.
This paper is dedicated to the 70th birthday of Prof. Friedrich Alfons Jung.