
Abstract
INTRODUCTION:
Mesenchymal stem cells (MSCs) have been described in breast cancer models to migrate towards carcinoma and integrate into tumor associated stroma supporting tumor growth, increasing their metastatic potency and contributing to tumor-angiogenesis. Platelet-derived growth factor (PDGF) isoforms (AA, BB, CC) stimulate growth, survival and motility of MSCs and certain other cell types. Noteworthy, breast carcinomas are known to express PDGF. We aim to further shed light on i) the relevance of the different PDGF isoforms on adipose tissue derived stem cells (ASCs) migration and ii) the underlying pathway dependent on PDGF stimulation.
MATERIALS AND METHODS:
Breast cancer cell lines were purchased and ASC’s were isolated from murine subcutaneous adipose tissue. The transmigration of ASC’s towards the PDGF-isoforms was assessed by using recombinant human PDGF-AA, PDGF-BB and PDGF-CC in a trans-well culture dish system. Transmigrated ASC’s were quantified in 5 randomly selected fields per condition using fluorescence microscopy after calcein-staining. PDGF-BB depended transmigration of ASC’s was verified by downregulation and overexpression of PDGF-BB in breast cancer cell line using lentiviral vectors. In addition, a PI3-kinase inhibitor (LY294002) and a MAP-kinase inhibitor (PD98059) were used to identify the pathway involved in the PDGF-BB mediated migration of ASC’s towards tumor.
RESULTS:
ASC’s transmigration significantly increased towards PDGF AA at 50 ng and only showed further increase by 500 ng which was similar to cell behavior when exposed to PDGF CC. In comparison, PDGF-BB significantly increased ASC’s transmigration already at a low level of 5 ng with further significant increase for 20 ng and 40 ng. Cell transmigration was blocked with PDGFR-α antibodies but only for PDGF-AA and PDGF-CC whereas PDGFR-β blockage showed a significant effect on transmigration for PDGF-BB and PDGF-CC but not for PDGF-AA. Neutralizing antibodies in combination with PDGF receptor blockage confirmed findings. In addition, only PI3-kinase inhibitor but not the MEK-1 selective inhibitor caused a significant decrease of transmigration for ASCs towards breast cancer cells.
DISCUSSION:
The transmigration of ASC’s is most significantly enhanced by PDGF-BB via the PI3-kinase pathway. This data support that PI3-kinase is an important key player for MSC migration towards malignancy which need further research to prevent tumor progression in early disease stage.
Introduction
Mesenchymal stem cells (MSCs) have been identified in various tissue which resides as pericytes within the basement membrane of small vessels with direct cellular contact to endothelial cells [1]. Moreover, MSCs have been described in breast cancer models to migrate towards carcinoma and integrate into tumor associated stroma supporting tumor growth, increasing their metastatic potency and contributing to tumor-angiogenesis [2–4]. The migration of MSCs towards malignancy is directed by a large variety of bioactive molecules (e.g. growth factors, cytokines, chemokines) but also by topographic microstructure of the environement [5] however, the type of cytokine released is dependent on the tumor cell type and its niche [6–9].
Recently, it has been reported that PDGFR-β signaling plays an important role in recruitment of MSCs towards tumor sites [10]. Noteworthy, breast cancer cells are known to express PDGF and radiation therapy has been shown to increase the release of PDGF-B by tumor cells, which not only attract MSCs to the tumor site but also induces the newly recruited MSC to differentiate into pericytes, and promote vasculogenesis and tumor growth [11]. In addition, it has been confirmed that MSCs express the corresponding receptors for PDGF ligands [8]. The essential role of PDGF-BB/PDGFR-β signaling in vascular maturation has been well-documented by genetic studies [12, 13]. During vascular development, endothelial cells (ECs) secrete PDGF-BB, which not only enhances pericyte motility but also forms a chemotaxis-like gradient to facilitate pericyte recruitment [14, 15]. Recently, several studies have correlated PDGF-BB signaling with vascular remodeling within tumors and showed that transgenic PDGF-BB expression in tumors can increase pericyte [16–18].
We have recently shown that adipose tissue contains multipotent stem cells that can differentiate into myofibroblasts when exposed to tumor cell derived TGF [19]. Amos et al. [20] demonstrated that adipose tissue-derived stem cells (ASCs) express pericyte lineage markers in vivo and in vitro, exhibit increased migration in response to PDGF-BB in vitro, reveal perivascular morphology when injected in vivo, and contribute to increases in microvascular density during angiogenesis by migrating toward vessels. We have shown that ASCs are incorporated into tumor vessels [3, 21] and are capable of inducing myofibroblast differentiation [19]. However, to date it is not clear by which pathway PDGF-BB acts on ASCs and regulates their migration.
In the present study we aim to further shed light on i) the relevance of the different PDGF isoforms on ASCs migration and ii) the underlying pathway dependent on PDGF stimulation.
Materials and methods
Cell lines and cell culture techniques
4T1 murine breast cancer cells were purchased from American Type Culture Collection (ATCC) and cultured in RPMI 1640 medium (Cellgro) supplemented with 10% fetal bovine serum (FBS) (Atlanta Biologicals), L-glutamine (Cellgro), and penicillin-streptomycin (Cellgro) at 37°C in a 5% CO2-containing chamber.
Isolation of murine ASCs
Perirenal, pelvine, and subcutaneous fat tissues were dissected from mice and processed as described previously [22].
ELISAs for PDGF-BB
Serumfree media (RPMI 1640) were conditioned by 4T1 for 48 hours, centrifuged at 1500 rpm for 5 min and passed through a 0.45 μm filter system (Millipore). Conditioned media (CM) were prepared fresh and used immediately for migration assay or stored at -20°C for ELISA. ELISAs were performed using the PDGF-BB Quantikine Kit (R&DSystems) according to the manufacturer’s instructions.
Overexpressing and silencing of PDGF-B in 4T1 cell line
A lentiviral vector overexpressing the murine PDGF-B (mPDGF-B/PLVX) was constructed by subcloning murine PDGF-B (Invivogen) into the PLVX vector (Clontech) between EcoRI and XbaI restriction. A specific Kozak sequence for the PDGF-B gene (GTCGGC) was inserted in front of the gene to achieve a higher protein expression in target cell lines. High fidelity PCR (Roche) and following primer were used:
sense 3’ CTATAAATAAGGAATTCGTCGGCATGAATCGCTGCTGGGCGC 5’;
antisense 3’ GTTTATATAATCTAGAGGCTCCGAGGGTCTCCTTCAGGGCC 5’
4T1 cell lines were infected with lentiviral shRNA constructs (Sigma) targeting PDGF-B mRNA to silence secretion of PDGF-B. An empty pLVX and non-target shRNA were used to establish control cell lines with no interference of PDGF-BB secretion (Fig. 1e).
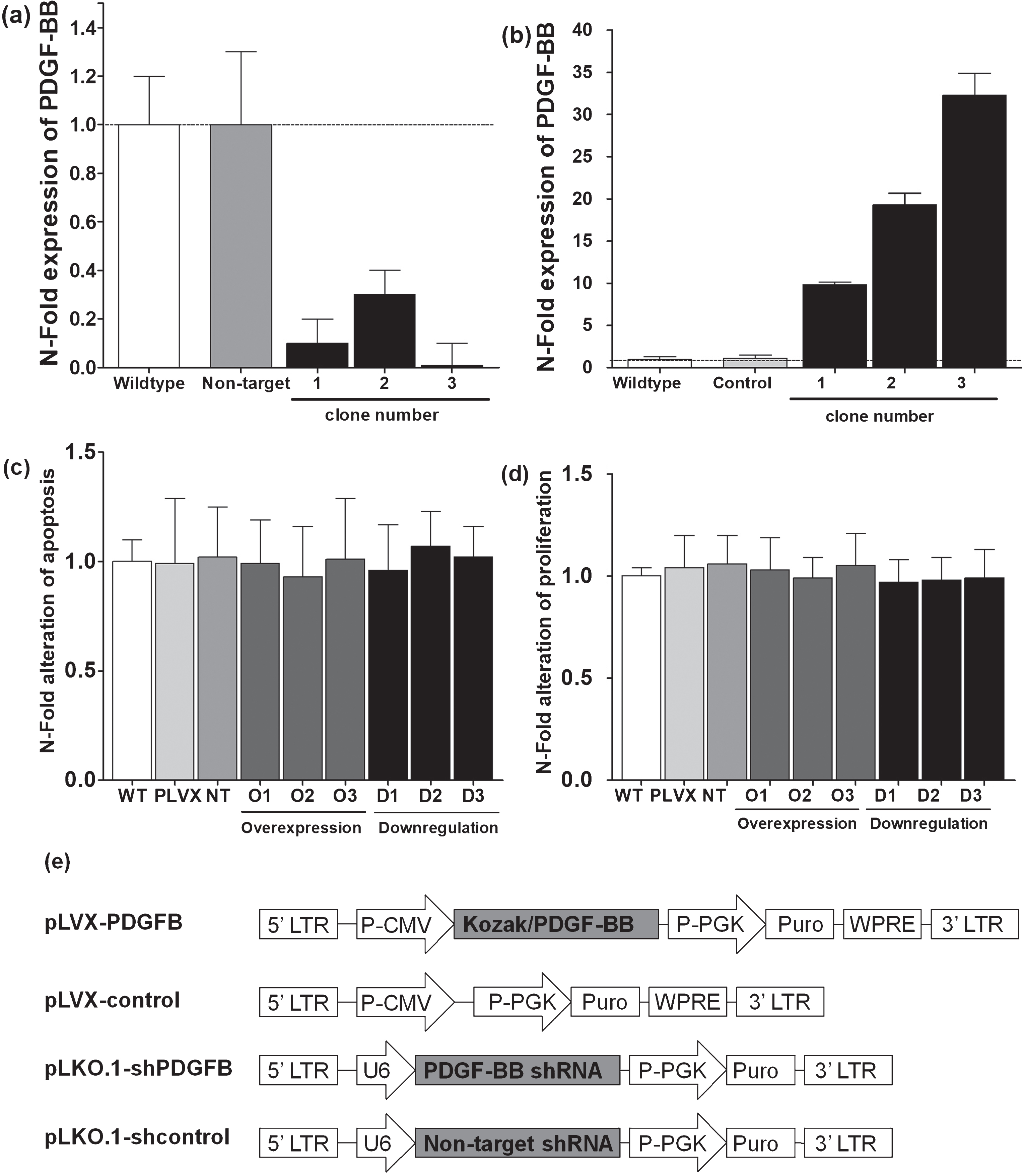
PDGF-BB protein levels for different clones after successful lentiviral transfection for downregulation (a) and overexpressing (b) of PDGF-BB. No significant difference was evident between transfected clones and appropriate controls regarding apoptosis (c) or proliferation (d). Lentiviral construct for silencing of PDGF-BB mRNA in breast cancer cells was based on a pLKO.1 vector whereas overexpression of PDGF-BB was achieved by cloning the appropriate sequence into a pLVX backbone (e).
Lentiviral vectors were generated by calcium phosphate-mediated transfection of 293T cells as described previously by our group [23].
4T1 cells were infected with the appropriate lentiviral construct and selected by puromycin for 10 days. ELISA for murine PDGF-B (RnDSystems) was used to measure the level of PDGF-B in 4T1 overexpressing or downregulation this specific protein (Fig. 1a, b).
Proliferation and apoptosis assay
Cell proliferation was monitored using Cell Proliferation Reagent Kit I (Roche).1×103 lentiviral transfected cells were seeded in quintuplicates into each well of a 96 well plate and allowed to attach for a period of 24 h at 37 °C. Afterwards, cells were incubated in fresh applied cell medium for 24 h. Analysis of proliferation was carried out following the kit instruction. Absorbance was measured immediately using a microplate reader spectrophotometer (BioTek). Measurements were performed at 492 nm (test wavelength) and at 620 nm (reference wavelength), to correct for noise.
Apoptosis was investigated by using Cell Death Detection ELISAplus (Roche), which monitors DNA fragmentation. All lentiviral infected cell clones were seeded into 24 well plates at 1×104/cm2 and grown to confluency for 72 h. Cells were harvested using trypsin-EDTA. Cell pellets were collected and apoptosis assays were performed according to the manufacture protocol. Each experimental condition was carried out with at least three samples and was repeated at least three times. The reaction products in each 96-well plate were read using a BioTek microplate reader.
Transmigration assay
The Transwell Migration System (BD Biosciences) with a 3-μm pore size was used for the transmigration experiments. Murine ASCs were plated in the upper chamber of the system. The transmigration of ACS’s towards the PDGF-isoforms was assessed by using recombinant PDGF-AA, PDGF-BB and PDGF-CC (RnDSystems) in a trans-well culture dish system. After 8 h of transmigration through the membrane, cells were fixed and stained with calcein (Sigma). Migrated cells were quantified in randomly selected fields (n = 3) per condition (triplicate, n = 3) using fluorescence microscope (Nikon TE-2000U) with a Cascade camera (Photometrics) and Image J software [24].
Inhibitor assay
Commercially available Inhibitors of PI3-kinase (PI3K) and MAP kinase (MAPK) were used to identify the pathways involved in the PDGF-BB mediated migration of ASCs towards breast cancer cells. PI3K/Akt signaling was inhibited by LY294002 (Cell Signaling) at a final concentration of 50 μM and MAPK signaling was inhibited by PD98059 (Cell Signaling) at a final concentration of 10 μM. ASCs were incubated in culture medium containing the inhibitor for 48 h prior to migration assay and the inhibitors remained in the culture medium during the subsequent transmigration assay. Untreated cells served as control.
Reverse transcription-polymerase chain reaction
Total RNA was extracted from parental and transfected cancer cell line using RNAqueous kit (Ambion) and was reverse transcribed using iScriptTM cDNA synthesis kit (Biorad). Expression level of PDGF-BB and GAPDH mRNA was determined by real-time PCR on an iCycler (Biorad) using SYBRGreen assay (Applied Biosystem) and data were analyzed as previously described [2].
Results
Dose dependent migration of ASCs
To characterize the PDGF dependent migration of ASCs, different members of the PDGF family were used for in vitro migration assay. Transmigration of ASCs significantly increased for recombinant PDGF-BB at a concentration of 5 ng/ml and further increased over a small range of protein concentration in a dose dependent manner (Fig. 2). In contrast, transmigration of ASCs only showed a 2-fold increase of migration index for PDGF-AA or PDGF-CC at the same range of protein concentration as apparent for PDDG-BB. Further increase of transmigration was only evident for protein concentration of PDGF-AA and PDGF-CC at 500 ng/ml (data not shown).

Dose dependent transmigration of ASCs was only evident for PDGF-BB and showed a significant increase for all applied concentrations (**p < 0.0001). In contrast, increased concentration of PDGF-AA and PDGF-CC did not show any variation of ASCs transmigration capacity as seen for PDGF-BB concentration (5 ng/ml to 40 ng/ml) but a significant increase of transmigration when compared to control (*p < 0.01). However, a further increase of transmigration was only apparent for concentration higher than 500 ng/ml (data not shown).
PDGF-B was over expressed or downregulated in breast cancer cell line (4T1) using lentiviral constructs (Fig. 1), and single clones were picked, grown to culture and investigated for PDGF-BB expression, proliferation and apoptosis in comparison to the parental cell line. PDGF-BB silencing or overexpression revealed no significant difference regarding apoptosis or proliferation when compared to control transfected or non-transfected cell lines (Fig. 1c, d).
PI3-Kinase pathway
Cell transmigration was blocked with PDGFR-α antibodies but only for PDGF-AA and PDGF-CC whereas PDGFR-β blockage showed a significant effect on transmigration for PDGF-BB and PDGF-CC but not for PDGF-AA. Neutralizing antibodies in combination with PDGF receptor blockage confirmed findings (Fig. 3a).
In addition, only PI3K inhibitor (LY294002) but not the MAPK inhibitor (PD98059) was associated with a significant decrease of transmigration for ASCs towards breast cancer cells (Fig. 3b). Western blot analyses of ASCs revealed strong expression of PDGFR-β and only weak expression of PDGFR-α when compared with control cell line (Fig. 3c) as previously reported by our group [25].
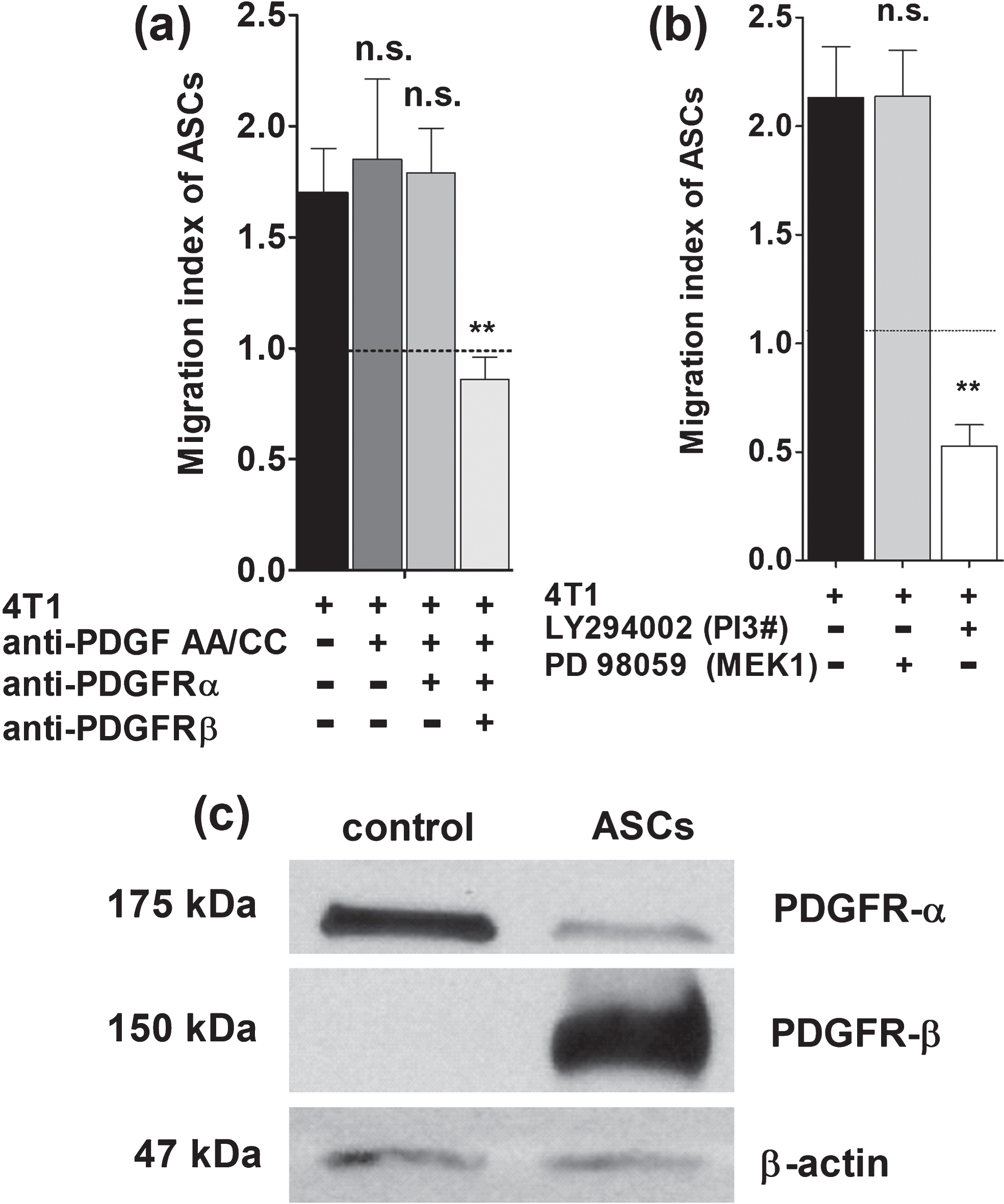
(a) Neutralizing antibodies against PDGF-AA und PDGF-CC did not affect transmigration of ASCs even when PDGFR-α was blocked (n.s., non-significant). However, significant decrease of ASCs transmigration was apparent when PDGFR-beta was blocked (**p < 0.0001). (b) LY294002 targets PI3K and resulted in a significant decrease of ASCs transmigration towards breast cancer conditioned medium (**p < 0.0001). In contrast, the selective inhibitor of MAPK (PD98059) did not interfere with transmigration of ASCs. (c) Western blot analyses of ASCs revealed strong expression of PDGFR-β and only weak expression of PDGFR-α when compared with control cell line (breast cancer cell line).
The current study shows that the transmigration of ASCs is most significantly enhanced by PDGF-BB via the PI3K pathway. Our data support that PI3K is an important key player for ASCs migration towards malignancy which need further research to prevent tumor progression in early disease stage.
The role of MSCs in breast cancer has been the focus of research for many years. It has been shown, that MSCs enhance tumor growth and are able to support tumor angiogenesis and the process of metastases [2–4]. Recent studies suggest that MSCs are attracted to tumor site and subsequently differentiate into pericytes or cancer associated fibroblasts [3]. MSCs accumulate at the sites of tumor and establish a microenvironment supporting growth by secreting various chemokines and cytokines (e. g. PDGF) [26]. We could show, that transmigration of ASCs towards breast cancer is especially dose dependent by breast cancer secreted PDGF-BB. A recent study strengthened the link between migration of MSCs and tumor by showing that the irradiation of breast cancer enhanced the release of PDGF-BB by cancer cells which caused enhanced migration of MSCs towards the tumor [27]. In addition, an animal study underline the importance of PDGF-BB mediated tropism of MSCs towards malignancy showing that MSCs localized significantly more to high expressing PDGF-BB gliomas [28].
Our results indicate that tumor-derived PDGF-BB mediates migration of ASCs through PDGFR-β which is expressed on ASCs. Pretreatment of ASCs with neutralizing antibodies reduced in vitro migration of ASCs only when PDGFR-β was targeted. This is in line with a recent report showing that blocking of PDGFR-β on progenitor cells abolished PDGF-BB induced MSCs migration [29]. Other striking evidences are provided by using a nuclease-resistant RNA aptamer (Gint4.T) that binds to the extracellular domain of the PDGFR-β and impedes the ligand-dependent cross-talk between MSCs and tumor cells both in vitro and in vivo [30–32].
We further investigated the downstream signaling of PDGFR-β pathway by using PI3K inhibitor (LY294002) and MAPK inhibitor (PD98059) whereas only LY294002 was associated with a significant decrease of ASCs migration towards breast cancer cells. This is in line with experiments using transgenic mice carrying mutation of PDGFR-β which revealed a signaling through PI3K [33]. In addition, a recent study revealed that MSCs stimulate tumor growth and vascularization within colorectal cancer xenograft model in vivo and induce activation of Akt and ERK in endothelial cells, thereby increasing their recruitment and angiogenic potential [34].
Currently there is no doubt that MSCs can have tremendous effects to the microenvironment of tumor regarding angiogenesis, tumor growth and metastasis. It is crucial to understand the tropism of MSCs for breast cancer or any other malignant tumor in order to control or target MSCs with a therapeutic approach.