
Abstract
Non-swelling hydrophobic poly(n-butyl acrylate) network (cPnBA) is a candidate material for synthetic vascular grafts owing to its low toxicity and tailorable mechanical properties. Mesenchymal stem cells (MSCs) are an attractive cell type for accelerating endothelialization because of their superior anti-thrombosis and immune modulatory function. Further, they can differentiate into smooth muscle cells or endothelial-like cells and secret pro-angiogenic factors such as vascular endothelial growth factor (VEGF). MSCs are sensitive to the substrate mechanical properties, with the alteration of their major cellular behavior and functions as a response to substrate elasticity. Here, we cultured human adipose-derived mesenchymal stem cells (hADSCs) on cPnBAs with different mechanical properties (cPnBA250, Young’s modulus (E) = 250 kPa; cPnBA1100, E = 1100 kPa) matching the elasticity of native arteries, and investigated their cellular response to the materials including cell attachment, proliferation, viability, apoptosis, senescence and secretion. The cPnBA allowed high cell attachment and showed negligible cytotoxicity. F-actin assembly of hADSCs decreased on cPnBA films compared to classical tissue culture plate. The difference of cPnBA elasticity did not show dramatic effects on cell attachment, morphology, cytoskeleton assembly, apoptosis and senescence. Cells on cPnBA250, with lower proliferation rate, had significantly higher VEGF secretion activity. These results demonstrated that tuning polymer elasticity to regulate human stem cells might be a potential strategy for constructing stem cell-based artificial blood vessels.
Introduction
Artificial blood vessels are promising alternatives of allogeneic and autologous grafts for treating vascular diseases. The functionality of artificial blood vessels is highly dependent on the scaffold materials as well as their interacting cells [1–3]. The scaffold is required to be non-toxic and be able to support favorable cell attachment, differentiation and secretion of functional proteins. In addition, appropriate mechanical strength of the scaffold material is necessary to withstand the in vivo hemodynamic forces. The reported poly(n-butyl acrylate) networks (cPnBAs) have demonstrated a high potential as synthetic vascular grafts [4]. The cPnBA polymers showed negligible cytotoxicity and could support the survival of cell lines as well as primary endothelial cells [4, 5]. They also presented a high immuno-compatibility as demonstrated via the in vitro immune activation test [6]. Importantly, by varying the crosslinker content the elasticity of the cPnBA could be easily tailored to match the elasticity of native arteries, which varies in the range between 100 kPa and 1000 kPa [7–9].
Native blood vessels, aside from capillaries, consist of three layers. The endothelial cells (ECs) form the luminal side of the tunica intima and play a role to resist thrombosis and facilitate laminar flow of blood, and smooth muscle cells (SMCs) in the tunica media are responsible for remodeling of the vessel wall as well as stabilizing the elasticity and radial compliance of the vessel [1, 10]. In order to mimic the structure and function of native vessels, several types of cells have been used for preparing artificial blood vessels, including ECs, SMCs, endothelial progenitor cells (EPCs), mesenchymal stem cells (MSCs) and embryonic stem cells (ESCs) [1, 11]. Notably, MSCs have shown a high potential due to their intrinsic advantages such as abundance, self-renewal capacity, high proliferative potential and long-term viability. They could accelerate the process of endothelialization as they own the superior anti-thrombosis and immune modulatory functions. Importantly, they could differentiate into SMCs or endothelial-like cells and secret pro-angiogenic factors such as vascular endothelial growth factor (VEGF) to promote vascularization. Further, MSCs share the similar origin as pericytes, which are contractile cells abundant in blood microvessels. Pericytes can interact with endothelial cells and modulate the stabilization and hemodynamics of vessels [12, 13]. Therefore, pre-seeding of MSCs on a vascular graft in vitro may improve the function of the artificial blood vessels after implantation. In this case, the graft material should allow the favorable behaviors of MSCs including the high cell attachment, proliferation, survival and secretion of functional factors.
Accumulated evidences have proven the sensitivity of MSCs to their surrounding microenvironment. The cellular behavior and function of MSCs could be regulated by various physical cues such as substrate mechanical property [14], surface topography [15] and external force [16] via the process termed mechanotransduction [17]. Among those, a lot of attention has been drawn to the cellular response of MSCs to substrate elasticity [18]. For example, the cell fate of MSCs has been shown to be influenced by matrix elasticity at the tissue level. The soft matrices (E = 0.1–1 kPa) mimicking brain tissue, stiffer matrices (8–17 kPa) mimicking muscle and more stiffer matrices (25–40 kPa) mimicking the crosslinked collagen of osteoids could lead to MSC neurogenesis, myogenesis and osteogenesis [19]. However, currently little is known about the MSC response to scaffold elasticity in the range matching the native arteries. In this study, we hypothesized that the favorable cellular behaviors of MSCs for constructing the stem cell-based artificial blood vessels could be achieved via tuning the scaffold elasticity in the range of arteries. We investigate two types of cPnBA films with different elastic moduli (E = 250 kPa for cPnBA250; 1100 kPa for cPnBA1100 at ambient temperature) as substrate materials for human adipose-derived mesenchymal stem cells (hADSCs). Tissue culture plate (TCP) was included as a reference material. The major cellular behaviors were assessed, including the early stage attachment, proliferation, actin cytoskeleton assembly, viability, apoptosis, cellular senescence, secretion of cytokines and cell migration.
Materials and methods
cPnBA films
The cPnBA polymer networks were synthesized via bulk radical polymerization as described previously [4]. The solution containing n-butyl acrylate (nBA), poly(propylene glycol) dimethacrylate (PPGDMA) as crosslinker, and AIBN as initiator was filled into the molds formed by two glass plates and a 1.0 mm thick Teflon spacer for polymerization. The obtained polymer films were purified by swelling–deswelling in mixtures of n-hexane and chloroform, dried at 40°C under vacuum, and sterilized using ethylene oxide (gas phase: 10 wt% ethylene oxide, 54°C, 65% relative humidity, 0.17 MPa, 3 h gas exposure time and 21 h aeration phase). The amount of crosslinker was changed to tailor the properties of the cPnBAs, which have been characterized in our previous study, including the polymer wettability evaluated via dynamic contact angle measurements, the surface profiles and roughness measured via optical profilometry as well as the mechanical properties determined by tensile tests at ambient temperature [4]. In this work, the substrates with Young’s modulus of 250±20 kPa (cPnBA250, 0.4 wt% PPGDMA) and 1110±20 kPa (cPnBA1100, 7.3 wt% PPGDMA) were tested. To perform experiments with cells, the round-shaped cPnBA films with a diameter of 13 mm (Fig. 1A) were put into the commercial standard 24-well TCP (Greiner Bio-One, Frickenhausen, Germany), which was made of polystyrene with modified surface for adherent cell culture. In parallel, the TCP without cPnBA films was used as a reference material.
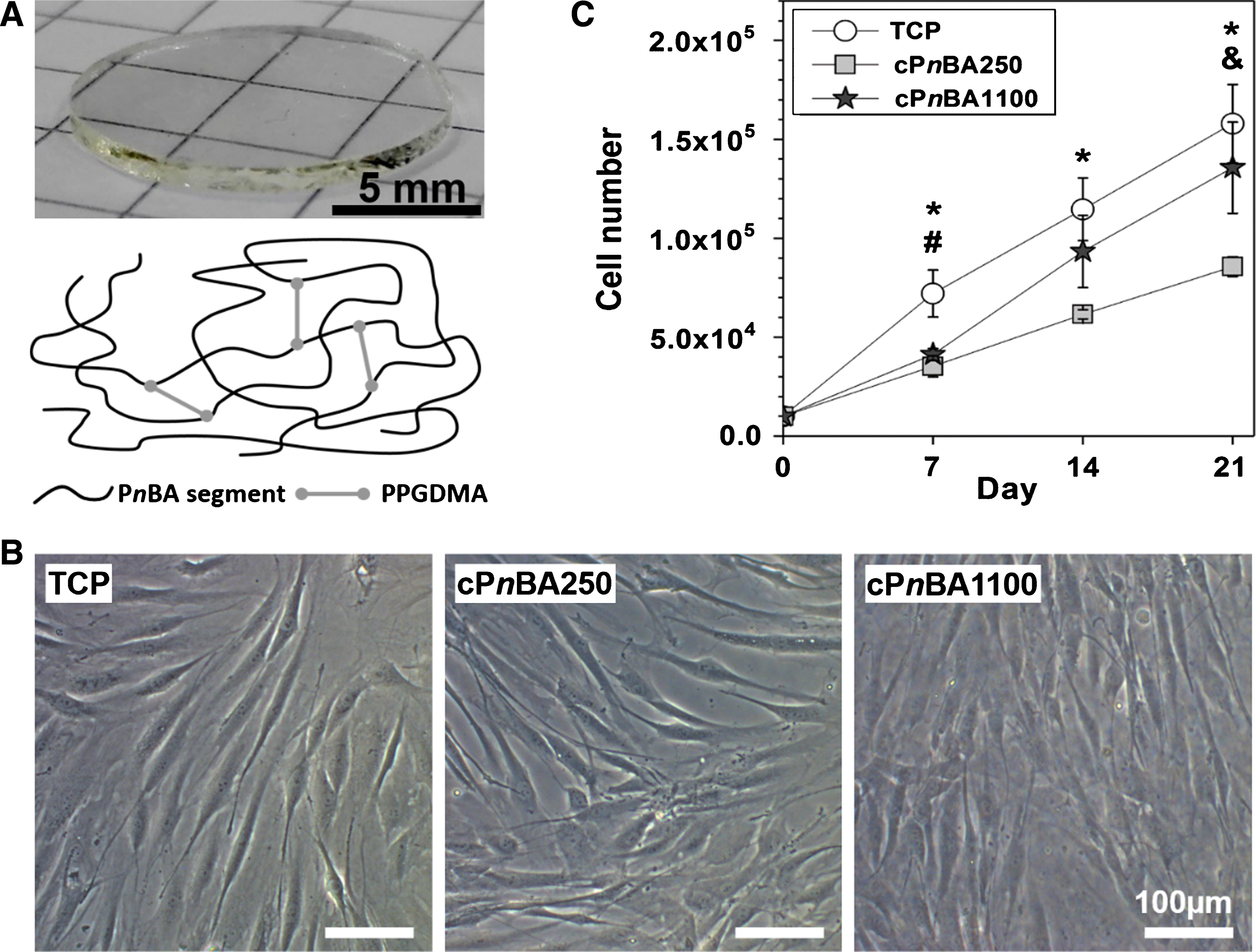
Adhesion and proliferation of hADSCs on TCP and cPnBA films. (A) Round-shaped cPnBA films with a diameter of 13 mm could be put into the commercial standard 24-well tissue culture plates for hADSC culture. The polymer contains PnBA segments crosslinked by PPGDMA. The mechanical properties of the films were tailored by changing the crosslinker amount. (B) Representative images showing early stage (24 hours) cell adhesion on cPnBA films and TCP. (C) Proliferation of hADSCs. The cell number was determined at indicated time points (n = 3, p < 0.05 (*: TCP vs cPnBA250, #: TCP vs cPnBA1100, &: cPnBA250 vs cPnBA1100)).
The hADSCs were isolated from adipose tissue obtained by abdominal liposuction from a female donor after informed consent (No.: EA2/127/07; Ethics Committee of the Charité - Universitätsmedizin Berlin, approval from 17.10.2008) [20]. The fat tissue was dissociated for 60 minutes at 37°C with collagenase NB4® (Serva GmbH, Heidelberg, Germany), followed by passing through a 100 μm cell strainer to remove the undissociated tissue. Then, the obtained cells were washed and cultured in a humidified atmosphere containing 5 vol% CO2 at 37°C. The non-attached cells were removed after 2 days and the attached cells were cultured in stem cell medium (MSCGMTM, Lonza, Walkersville, MD, USA) without additional differentiation promoting factors. To maintain cell growth, the medium was changed every 3 days and the cells were passaged at a ratio of 1:3 when reaching ∼90% confluence.
Cell adhesion and proliferation
The cells were seeded at a density of 1.0×104 cells/well. The early stage cell adhesion was evaluated via phase-contrast microscope after 24 hours. The cell proliferation rate was determined using a Cell Counting Kit-8 (CCK-8, Dojindo Molecular Technologies, Munich, Germany). At the indicated time points, the old medium was replaced with 400 μl of fresh medium for each well, followed by addition of 40 μl of CCK-8 solution. After 2 hours of incubation at 37°C, 110 μl of medium/CCK-8 mixture was transferred from each well into a transparent 96-well TCP. A microplate reader (Infinite 200 PRO®, Tecan Group Ltd., Männedorf, Switzerland) was used to measure the absorbance of the mixture at 450 nm and at a reference wavelength of 600 nm. Finally, the cell number was calculated using a standard curve, which was generated by measuring the absorbance value of a series of samples with known cell number.
Fluorescent staining
The cytoskeleton organization of the cells was studied after 2 weeks of culture on different materials. The fibrous actin (F-actin) stress fibers were stained with rhodamine conjugated phalloidin (Invitrogen, Paisley, UK) according to the given protocol. The cell nuclei were counterstained with 4’,6-diamidino-2-phenylindole (DAPI). The cell viability was evaluated via a live/dead staining. After 2 weeks of culture on the materials, the cells were stained with fluorescein diacetate (FDA, 25 μg/ml) and propidium iodide (PI, 2 μg/mL) to identify the living and dead cells, respectively. After staining, the cells were washed with phosphate-buffered saline (PBS) buffer for three times, and the fluorescence images were obtained with a confocal laser scanning microscope (LSM 510 META, Carl Zeiss, Jena, Germany).
Apoptosis assay
The activation level of caspase-3/7 was detected to assess the cell apoptosis using an apoptosis kit (Caspase-Glo®, Promega, Madison, WI, USA) at day 1 and 7 after cell seeding. The old medium was first replaced with 240 μl of fresh culture medium for each well. Then, 200 μl of Caspase-Glo® 3/7 reagent was added and the mixture was shaken orbitally at 300 rpm for 30 seconds. Following an incubation at room temperature for 90 minutes, 200 μl of mixture was transferred from each well into a 96-well opaque (white) plate, and the luminescence intensity was measured using a microplate reader (Infinite 200 PRO®, Tecan Group Ltd., Männedorf, Switzerland). The result was expressed as the relative light units (RLU) of the measured samples.
Cellular senescence assay
To measure the level of cellular senescence, the activity of the acidic senescence-associated ß-galactosidase (SA-ß-Gal) was examined at day 7 and 21 after cell seeding using a cellular senescence assay kit (Cell Biolabs, Inc., San Diego, CA, USA). The culture medium was carefully removed and the cells were washed 2 times with cold PBS. Then, 400 μl of lysis buffer was applied for each well and incubated at 4°C for 5 minutes to lyse the cells. The obtained cell lysate was centrifuged for 10 minutes at 4°C to collect the supernatant. Finally, the activity of SA-ß-Gal in the supernatant was determined with the fluorometric substrate according to the provided protocol. In addition, the total protein in the cell extract was quantified using a BCA protein assay kit (Thermo Fisher Scientific, Bonn, Germany). The result was expressed as the fluorescence intensity of the measured samples normalized with the amount of the total protein.
Cytokine secretion assay
The concentration of secreted inflammatory cytokines in the conditioned medium was measured. The conditioned media, derived from the fresh media added at day 0, 4, 11 and 18 with the volume of 700 μl per well, were collected and stored at –20°C at day 1, 7, 14 and 21. The concentration of the cytokines in the conditioned media were measured using a Bio-Plex® 200 system (BioRad, Munich, Germany) according to the manufacturer’s instructions.
ELISA
The F-actin amount in cells on different materials was determined after 2 weeks of culture. Cells were first lysed and the F-actin concentration in the cell extract was measured using a human F-actin ELISA kit (Cusabio, Wuhan, Hubei, P. R. China). The amount of total protein in the cell extract was measured using the BCA protein assay kit (Thermo Fisher Scientific, Bonn, Germany). The result was expressed as the F-actin level normalized with the total protein amount in the cell extract. The vascular endothelial growth factor (VEGF) protein in the conditioned medium was quantified using a human VEGFA ELISA kit (Thermo Fisher Scientific, Bonn, Germany) at day 1, 7, 14 and 21 respectively after cell seeding. The conditioned medium was collected as described above, and the cell number on the materials was quantified with CCK-8 (Dojindo Molecular Technologies, Munich, Germany). The result was expressed as VEGF level normalized with the cell number.
Cell migration
After 2 days of culture on different materials, the cell nuclei were stained with Hoechst 33342. Then, the cells were tracked for 24 hours using a time-lapse imaging microscope (IX81 motorized inverted microscope, Olympus, Hamburg, Germany), equipped with a bold line cage incubator to provide the humidified atmosphere (37°C, 5 vol% CO2) for supporting cell survival. To analyze the migration parameters, the obtained images were processed using ImageJ software (National Institutes of Health, USA) with the plug-ins “manual tracking” and “chemotaxis and migration tool” (ibidi GmbH, Martinsried, Germany).
Statistics
The number of replicates for experiments was indicated in the figure legend of the respective assays, and the quantification data were expressed as arithmetic mean±standard deviation unless indicated otherwise. Statistical analysis was performed using one-way analysis of variance (ANOVA) with post hoc Tukey’s test. A p value less than 0.05 was considered to be statistically significant.
Results and discussion
In this study, we prepared cPnBA films with different mechanical properties as substrate materials for hADSCs. In order to solely study the influence of the materials’ elasticity on cells, the used cPnBA films should have similar surface properties to avoid the influence of other aspects. Our previous study has demonstrated that the surface properties are similar between these two types of cPnBA films, and remain constant after sterilization with ethylene oxide. The root mean square roughness Rq of sterilized samples was 1.0±0.3 μm for cPnBA250 and 0.7±0.2 μm for cPnBA1100, and the advancing contact angle was 116±9° and 104±5° respectively [4].
Cell adhesion and proliferation
A high cell adhesion rate onto the graft materials at the early stage is necessary for constructing artificial blood vessels with MSCs. This is of especial importance in the case that the cells were pre-cultured on the materials before implantation. After implantation, a tight attachment on the implants is required for cells to withstand the physiologic blood flow. In general, the cell adhesion could be improved by modifying the material surface such as coating with extra-cellular matrix (ECM) proteins. Here, the early stage cell adhesion was examined 24 hours after cell seeding. The cells could attach and spread on both cPnBA films even without ECM coating, showing typical spindle-shaped morphology similar to the cells on TCP (Fig. 1B). The integrin is the primary cell membrane receptor to mediate the cell-material interaction and perceive various signals in the process of mechanotransduction [21]. Integrin activation could trigger a series of cellular responses including cell adhesion, cytoskeletal formation, gene expression and proliferation [22]. Therefore, allowing a high cell adhesion rate is one of the most important features of the materials used as vascular grafts.
The proliferation of MSCs could be regulated by material elasticity [18]. For example, mouse MSCs cultured on hydrogels with different elasticity (Young’s moduli from 13 to 68 kPa) were found to proliferate faster on stiffer surface [23]. Similar results have also been observed in human MSCs cultured on hydrogels with different stiffness (storage moduli from 0.6–8.2 kPa) [24]. Here, our results suggested that the cells could even sense the difference of materials at the higher range of elasticity. The cells on cPnBA250 presented slower cell proliferation rate than on cPnBA1100. After 3 weeks, a fold change of 13.6±2.3 in cell number compared to the seeding number was observed on cPnBA1100, whereas the fold change was 8.6±0.5 on cPnBA250 (Fig. 1C). Among the tested materials, TCP allowed the highest cell proliferation rate.
Cell morphology and cytoskeleton
The morphology of MSCs and the structural organization of cytoskeletal filaments have been shown to be affected by elastic properties of the culturing materials. In general, MSCs contacted with a stiffer surface appeared more spread out than the cells on a softer material. Additionally, the F-actin stress fibers were stronger and better organized in the cells cultured on stiffer surface. Such a difference in cell morphology and cytoskeleton have been reported on softer gels with different stiffness (1–15 kPa) [25] and on polymers with higher elastic moduli (0.1–310 MPa) [26]. Here, to study the structure and organization of the cytoskeleton, the F-actin stress fibers and cell nuclei were stained after 2 weeks of culture on different materials (Fig. 2A). Cells growing on TCP showed well orientated strong stress fibers. In contrast, relatively weak stress fibers were observed in cells on cPnBA250 and cPnBA1100 films. Further, the F-actin level was determined with an ELISA assay. In consistence with the fluorescence images in Fig. 2A, the F-actin expression level on TCP was more than 2 times higher than on cPnBA films. In addition to the material surfaces, the different cell density on TCP and cPnBAs might lead to the different F-actin expression. It has been reported that the expression of integrin was dependent on the degree of cell confluence, although the patterns of change varied for some subunits [27]. Therefore, the cells on TCP, with a higher cell density as the result of higher proliferation rate, might present different integrin expression patterns from the cells on cPnBAs. This could further regulate the downstream F-actin expression. By comparing the cells on cPnBA250 and cPnBA1100, we did not observe a significant difference with respect to the structure, organization and expression level of F-actin (Fig. 2B). This result might be due to that the difference of elastic modulus of these two cPnBA materials was not large enough to induce the changes of morphology and cytoskeleton in hADSCs. This observation after 2 weeks of culture was in consistence with the cell morphology 24 hours after cell seeding (Fig. 1B), in which the cells presented the similar shape and size on cPnBA250 and cPnBA1100.
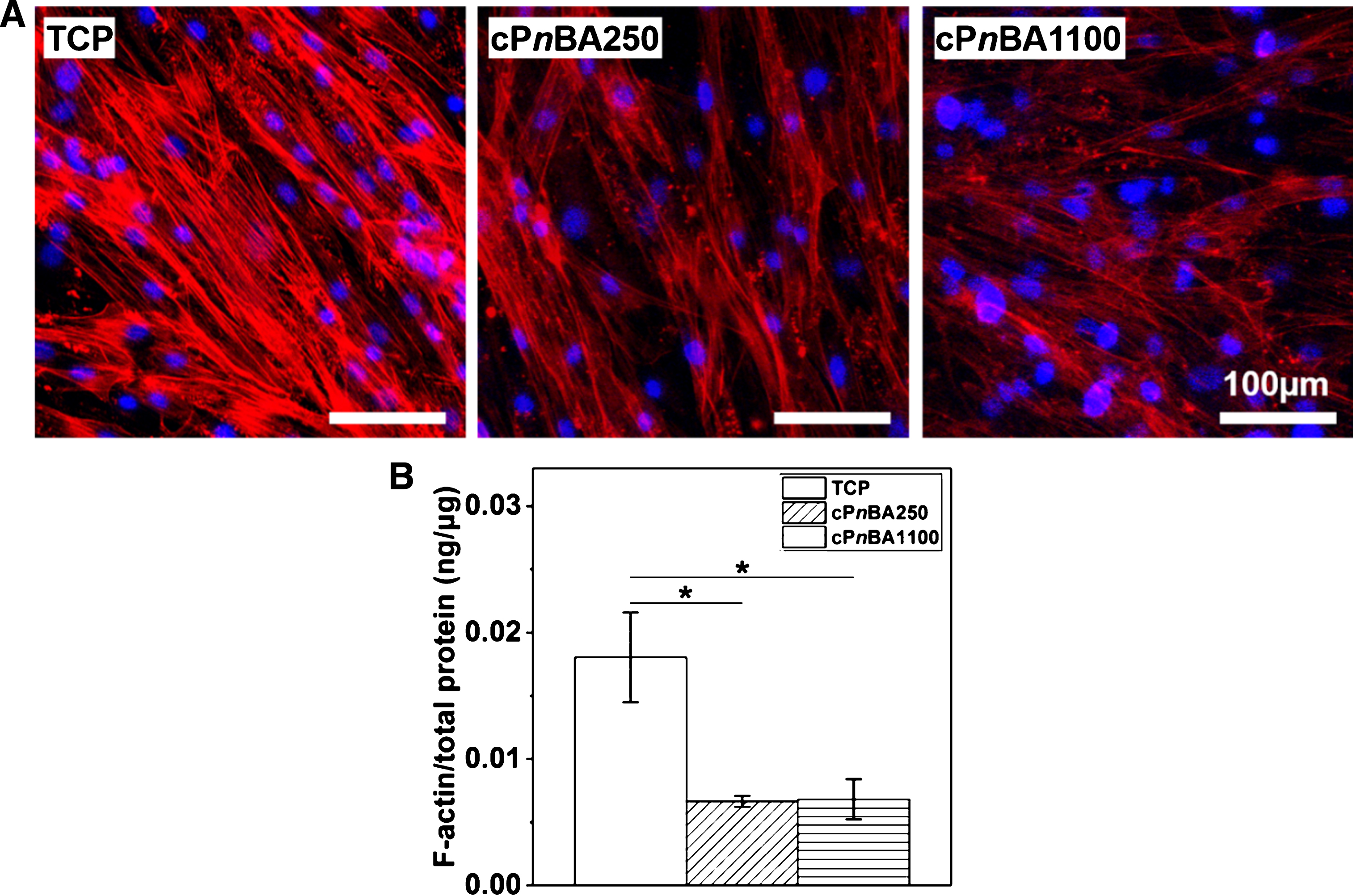
F-actin cytoskeletal structure and expression of hADSCs on TCP and cPnBA films. (A) Representative images of F-actin stress fibers (red) and cell nuclei (blue) staining after 2 weeks of culture. (B) F-actin amount in cells cultured on different materials for 2 weeks was determined with ELISA, and normalized with the level of total cell protein. (n = 3, *p < 0.05).
In order to evaluate the cytotoxicity of the materials, the live/dead staining experiment was performed (Fig. 3A). After 2 weeks of culture, the cells were stained with FDA and PI to identify the living and dead cells respectively. Similar to the cells on TCP, the cells on both cPnBA films presented high viability and no dead cells were found, indicating a negligible cytotoxicity of the cPnBA polymers. This result is in line with our previous tests using different cell types, in which high cell viability was observed for L929 cells cultured in the extracts of cPnBAs [4] and primary endothelial cells cultured directly on cPnBA surface [5].

(A) Representative images of live/dead staining of hADSCs with FDA (green) and PI (red) after 2 weeks of culture. (B) The cell apoptosis was evaluated by detecting the activation of caspase-3/7. (C) The level of cellular senescence was assessed by measuring the activity of the acidic senescence associated β-galactosidase. (n = 3; *p < 0.05).
In addition to the induced cell death by cytotoxicity, the cells could undergo the programmed cell death, which is an intentional suicide termed apoptosis [28]. The activation of caspase-3/7, a key biomarker of apoptosis, was determined to assess the cell apoptosis on different cPnBA films (Fig. 3B). At day 1, the apoptosis was at a similar level in cells on different materials. After 1 week of culture, a higher level of apoptosis was observed in cells growing on cPnBA films, as compared to cells on TCP (36% (cPnBA250) and 33% (cPnBA1100) increase to TCP). The difference of mechanical properties between cPnBA250 and cPnBA1100 did not affect the cell apoptosis.
The function of MSCs could be also affected by cellular senescence, as a result of intrinsic events or external stimuli. Senescence may lead to a permanent cell cycle arrest, making MSCs lose their self-renewal capacity. In addition, the alteration of major functions of MSCs could occur with cellular senescence, including the differentiation potential, immunoregulatory capacity, migratory and homing ability [29, 30]. The SA-ß-Gal, a biochemical marker of senescent cells, was therefore measured to evaluate the cellular senescence of hADSCs on different materials (Fig. 3C). After 1 week of culture, a significantly lower senescence level was observed on cPnBA1100, as compared to TCP. After 3 weeks, the cellular senescence was at a similar level in all tested groups. Taken together, our results demonstrated the high cell compatibility of cPnBAs for hADSCs, with respect to the cell viability, apoptosis and cellular senescence.
As the sensor and switcher of inflammation, MSCs own the function of immune-modulation. They could exert either a pro-inflammatory or an anti-inflammatory effect through the direct cell-cell contacts and/or secretion of multiple inflammatory cytokines [31, 32]. Previous studies have suggested that the secretion profile of MSCs could be regulated by the properties of cell culturing materials, such as the surface structure [33] and the substrate elasticity (ranging from 2 to 20 kPa) [34]. Therefore, the levels of secreted inflammatory cytokines were recorded up to 3 weeks in this study to investigate the influence of cPnBA elasticity on immunomodulatory capability of hADSCs (Fig. 4).
At day 21, cells on cPnBA1100 expressed a lower level of interleukin-6 (IL-6) than on TCP. Similar results were observed for pro-inflammatory interleukin-8 (IL-8) at day 7 and 14, monocyte chemoattractant protein 1 (MCP-1) at day 1 and 7, as well as anti-inflammatory interleukin-1 receptor antagonist (IL-1Ra) at day 7. No huge difference was observed for pro-inflammatory cytokines tumor necrosis factor alpha (TNFα) and interferon gamma (IFNγ). Taking all the data together, our results demonstrated that the difference of elasticity between cPnBA250 and cPnBA1100 did not show dramatic influence on secretion of inflammatory cytokines in hADSCs. The cPnBA polymers did not induce marked alteration in hADSC secretion profile as compared to TCP.
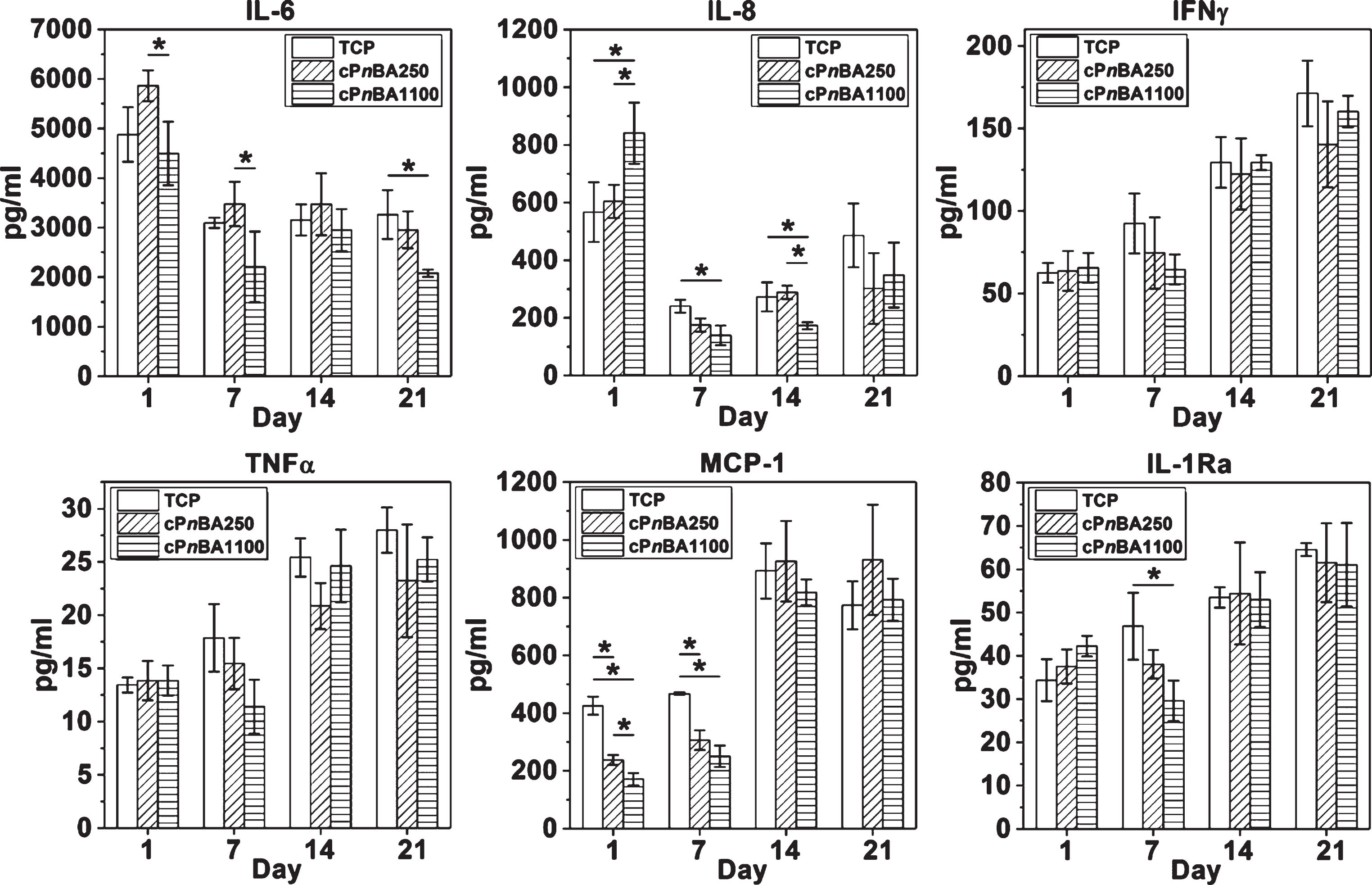
Secretion of inflammatory cytokines of hADSCs growing on TCP and cPnBA films. The concentration of cytokines in the conditioned medium was measured using a Bio-Plex 200 system. (n = 3, *p < 0.05).
The secretome of MSCs has emerged as an effective and safe therapeutic for treatment of a variety of diseases. As a cell free therapeutic, MSC secretome can promote angiogenesis and stimulate tissue adjacent cells and endogenous stem cells [35, 36]. Among the complex constituents of MSC secretome, VEGF is the major growth factor contributing to angiogenesis, which serves as the predominant inducer and mediator during angiogenic processes [37–39]. VEGF can stimulate proliferation and migration of endothelial cells located in pre-existing blood vessels to form new blood vessel sprouts, which is then anastomosis with each other to form the tubular-like vascular structures [40]. Further, VEGF can mobilize endothelial progenitor cells to the injury site, where they differentiate into mature endothelial cells and contribute to neovessel formation. The MSCs have been considered as a stable source of VEGF production, which is of great importance to improve the endothelialization and long-term patency of MSC-based artificial blood vessels. The VEGF secretion profile of hADSCs on different materials was assessed up to 3 weeks in this study (Fig. 5). At day 1, cells on TCP and cPnBA250 exhibited the higher level of VEGF expression than on cPnBA1100. Markedly, from day 7 to day 21, the VEGF secretion was induced by cPnBA250 to a significant level, which was around 2 times as high as that in cPnBA1100 and TCP cultures. This result suggested that cPnBA250 might be superior to cPnBA1100 for promoting pro-angiogenic capacity of MSCs. The mechanism for enhanced VEGF secretion on cPnBA250 might involve the integrin-mediated signaling pathways [15] as well as the stimulation of pro-inflammatory cytokines such as IL-6 [41], which need to be clarified in the future studies.

VEGF secretion of hADSCs on TCP and cPnBA films. The VEGF in the conditioned medium was quantified with ELISA assay, and was normalized with cell number. (n = 3; *p < 0.05).
Previous reports have shown that the migration of MSCs could be promoted by VEGF [42, 43]. The process of VEGF-induced MSC migration involved the promotion of focal adhesion formation and cytoskeletal rearrangement [44, 45]. Based on the significantly enhanced VEGF secretion of hADSCs on cPnBA250, we then evaluated the motility of cells on different materials after 2 days of culture, via recording the random cell movement for 24 hours using a time-lapse microscope. As expected, the hADSCs could freely move on all tested materials, with the fastest migration on cPnBA250 surface (Fig. 6A). The migration velocity of hADSCs on cPnBA250 (0.70±0.03 μm/min) was 78% and 157% higher than on TCP (0.39±0.02 μm/min) and on cPnBA1100 (0.27±0.02 μm/min), respectively (Fig. 6B). This observation was in consistence with the increased VEGF secretion of hADSCs on cPnBA250 film.
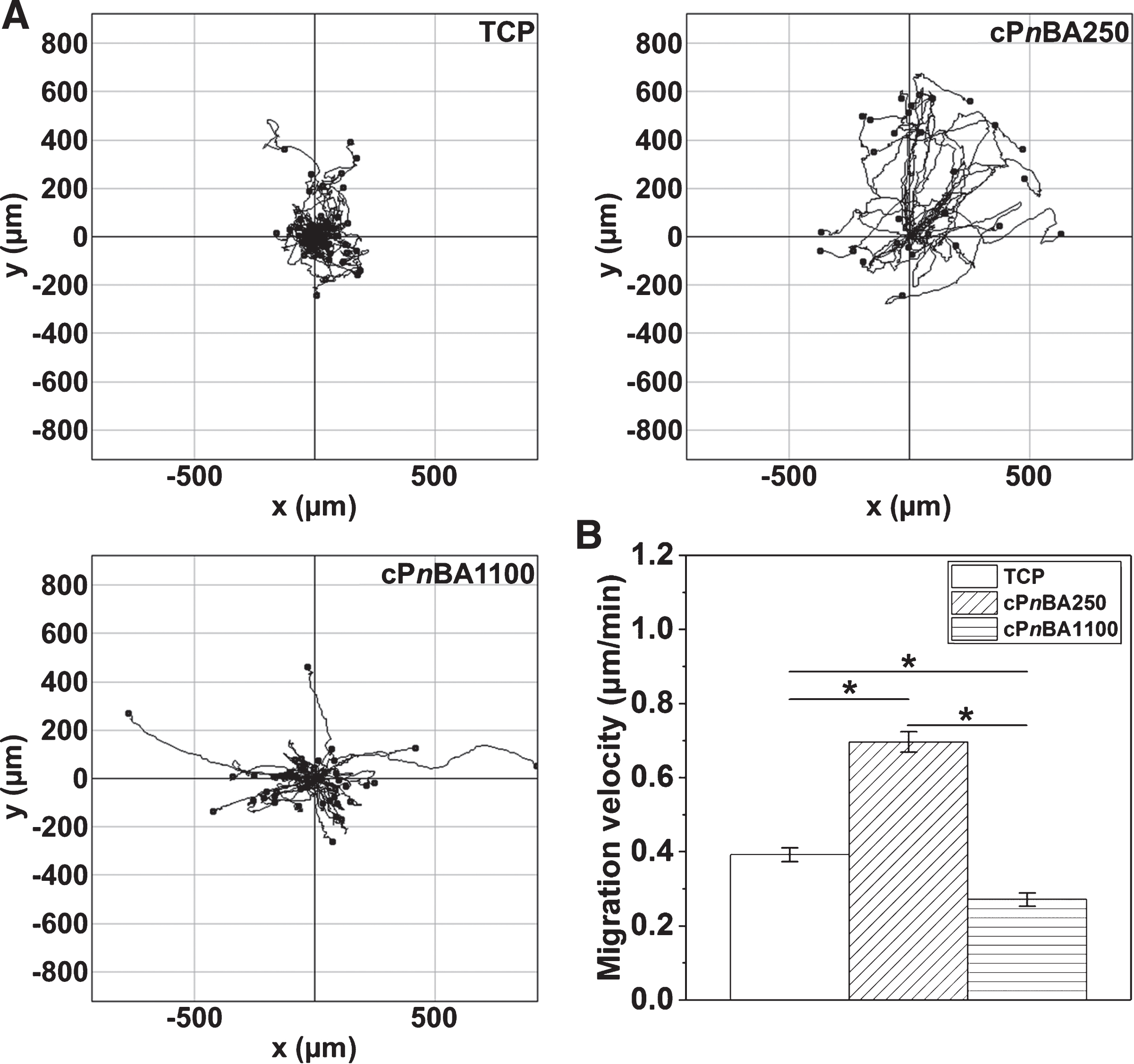
Migration of hADSCs on TCP and cPnBA films. After 2 days of culture, the cell nuclei were stained with Hoechst 33342 and were tracked up to 24 hours with a time-lapse microscope to generate the trajectories (A) and quantify the migration velocity (B). (Mean±SEM; n≥29; *p < 0.05).
Aiming to investigate the influence of scaffold elasticity in the range of native arteries on the cellular behavior and function of MSCs, we cultured hADSCs on cPnBA films with Young’s modulus of 250 kPa and 1100 kPa. Both cPnBA polymer networks presented high cell compatibility for hADSC culture. They did not show cytotoxicity to the cells, and allowed high cell attachment as well as long-term proliferation. The cell morphology, cytoskeleton assembly, apoptosis and cellular senescence of hADSCs were not markedly affected by the elasticity of tested cPnBA polymers. Compared to cPnBA250, a higher cell proliferation rate was observed on cPnBA1100. Notably, the pro-angiogenic capacity of hADSCs was enhanced by cPnBA250 substrate. The VEGF secretion activity of cells on cPnBA250 was significantly higher than on cPnBA1100, suggesting that culture of MSCs on cPnBA250 might promote the endothelialization process. Taken together, our results demonstrated the high potential of cPnBA polymer networks as synthetic vascular grafts for constructing stem cell-based artificial blood vessels. Tuning the elastic profile of the polymer scaffolds to regulate stem cells might be an effective strategy to improve the endothelialization and long-term engraftment of vascular prosthesis, which are of great importance to the function and clinical benefit of artificial blood vessels.
Footnotes
Acknowledgments
This work was financially supported by the Helmholtz Association of German Research Centers (Helmholtz-Portfolio Topic “Technology and Medicine”, Helmholtz Virtual Institute “Multifunctional Biomaterials for Medicine” (VH-VI-423), and programme-oriented funding) and the German Federal Ministry of Education and Research (BMBF project number 0315696A “Poly4Bio BB”).