
Abstract
BACKGROUND AND AIM:
Both chronic intermittent hypoxia (CIH) and chronic continuous hypoxia (CCH) are risk factors for cardiovascular disease, which are associated with cardiac systolic function and associated with dysfunction of endothelia and coagulation-fibrinolysis system in the vasculature. However, the different effects of these two hypoxic models are not fully understood. In our study, we systemically compared the effects of CIH and CCH on cardiac function and related factor levels in serum using rat model.
METHODS:
Forty-five male Sprague-Dawley rats were randomly divided into the normoxia control (NC), CIH and CCH groups. The rat CIH and CCH models were established, then the blood and tissue samples were collected to analyze the function of endothelium and the coagulation-fibrinolysis system. Also, the ultrasound cardiogram was performed to directly assess myocardial contractility.
RESULTS:
Both CIH and CCH significantly decreased the NO, eNOS, P-eNOS and AT-III levels in the rat serum but significantly increased the levels of ET-1, vWF, COX-2, NF-κB, FIB, FVIII and PAI-1 in the rat serum (P < 0.05). The expression of ET-1, VWF and ICAM-1 in CIH group were higher than CCH group (P < 0.05), however, the expression of CD62p was increased in CCH group but not in CIH group. The expression of t-PA in CIH group were lower than CCH group (P < 0.05), but there were no significant differences in CCH group and NC group (P > 0.05). Using transmission electron microscope, we found that the mitochondrial ultrastructure of thoracic aorta endothelial cells in CIH and CCH group were damaged. Moreover, the myocardial contractility in CIH and CCH group were significantly decreased compared with NC group.
CONCLUSION:
Our results suggested that CIH and CCH could cause endothelial dysfunction, dysfunction of the coagulation-fibrinolysis system and decreasing of myocardial contractility. Compared with CCH, CIH has greater effect on vasoconstriction and adhesion of vascular endothelial cells, and stronger procoagulant effect.
Keywords
Introduction
Hypoxia is the situation when the body exposed to low oxygen environments and the oxygen supply does not meet the tissue demand. Depending on the length of exposure, hypoxia can be either acute or chronic. According to the pattern of exposure, hypoxia also can be continuous or intermittent [1]. In humans, chronic continuous hypoxia (CCH) occurs in high altitude and in patients with chronic lung diseases such as chronic obstructive pulmonary disease and cystic fibrosis [2]. Chronic intermittent hypoxia (CIH) has been shown to occur during sleep apnea [3, 4], which always resulted in a number of pathological responses including oxidative stress, inflammation, and sympathetic activation. All of these responses may contribute to endothelial dysfunction [5, 6], which are involved in the disease such as inflammatory diseases, tumorigenesis, and the micro vascular damage. The physiological and pathological responses to these different types of hypoxia are different, but the underlying mechanisms are not fully understood.
In clinic, the commonly happened hypoxia are resulted from ischemia, which is insufficient blood flow to provide adequate oxygenation. Accumulated studies indicated that ischemia or hypoxia can induce metabolic and micro-rheological changes, activate the platelet and induce the failure of microvascular circulation [7–11]. Another commonly happened hypoxia is obstructive sleep apnea syndrome (OSAS), which is a type of disordered breathing during sleep and is basically characterized by CIH [12, 13]. It has been demonstrated that OSAS was associated with an increased risk of hypertension, type II diabetes, angina, myocardial infarction, congestive heart failure, stroke, and fatal cardiovascular events [14–16]. Especially, repetitive hypoxia-reoxygenation in OSAS can cause systemic damage to the vascular endothelium, which leads to endothelial dysfunction, platelet activation, coagulation and imbalance of fibrinolysis systems, and subsequent dysfunction of the coagulation and fibrinolysis system, and increases the tendency of thrombosis [12, 17]. A long term of intermittent hypoxia can lead to a decrease in cardiac systolic function. Patients with severe chronic obstructive pulmonary disease may have changes of hemorheology, endothelial function, coagulation and fibrinolytic system [18, 19] and myocardial contractility, which probably caused by CCH [20]. Although CIH has been believed to be the primary mediator of increased risk of vascular disease in OSA [21], both CIH and CCH can cause cardiovascular events, which may be related to abnormal hemorheology, endothelial dysfunction, dysfunction of coagulation and fibrinolytic system, and changes in myocardial contractility function [22, 23].
However, it is very important to differentiate whether chronic hypoxia occurs intermittently or continuously. Cardiovascular risk is markedly increased in patients with CIH compared with patients with CCH [24]. The frequent change between oxygen desaturation and reoxygenation in patients with CIH is associated with increased hypoxic stress, increased systemic inflammation, and enhanced adrenergic activation followed by endothelial dysfunction and increased arteriosclerosis [25, 26]. Compared to potentially investigated effects of CIH, the pathophysiologic consequences of CCH are less well understood. Up to date, there’s little study testing the hypothesis that exposure to CIH versus CCH will yield a differential pattern of response using CCH and CIH rat models. In our present study, we systemically compared the different effects of CCH and CIH on cardiac function using rat model, which provided experimental evidence to well understand the consequences of CIH and CCH on cardiac vascular system.
Material and methods
Animals and models of hypoxia exposures
The adult male Sprague-Dawley rats (180–200g body weight) were used for all experiments and randomly divided into the normoxic control (NC) group (n = 15), CIH group (n = 15) and CCH group (n = 15). All rats were fed with water and standard chow ad libitum. Room temperature was maintained at 22–24°C. All experimental protocols were approved by the Committee on the Use of Animals for Teaching and Research of the Medical School of Wuhan University (WDRM20171020) and conformed to the National Institutes of Health guide for the care and use of laboratory animals.
Rats were placed chambers under one of three experimental conditions: normoxia, intermittent hypoxia, or continuous hypoxia for consecutive 5 weeks. All chambers had the same size and dimensions, and the rat numbers were equal for each chamber. The NC rats were housed in the normoxia animal chambers (FiO2 = 21%). The IH exposure procedure was shown as follows: Rats were placed in chambers, which were alternately given 30s of normobaric hypoxia (10% FiO2) and 60s of normoxia (FiO2 = 21%) for 8 h daily. Normobaric hypoxia was generated by flushing the chambers with alternating mixtures of 100% oxygen and 100% nitrogen, under a time-concentration cycled mode in a 90-second-long cycle, at a stable and sufficient rate controlled by the gas control delivery system. The animals were kept in the chambers by flushing into air after the intermittent hypoxia exposure. The CCH rats were housed in identical chambers which were kept the 10% FiO2 by flushing into nitrogen with stable flow continuously for 24 h every day. The accurate oxygen concentration in the chamber was monitored using an CY12-C oxygen analyzer (Meicheng electrochemical analyzer factory, China) (Fig. 1).
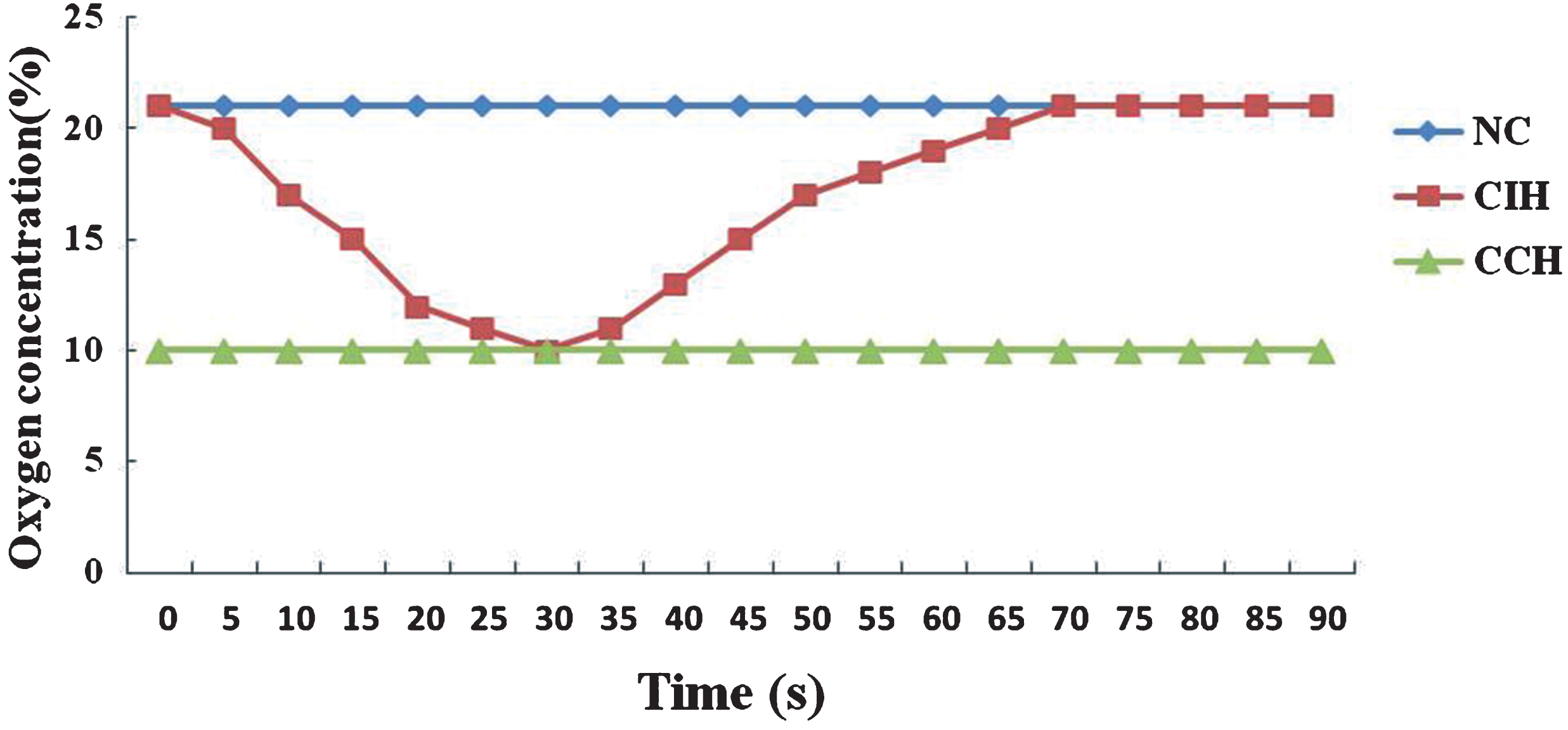
The traces of oxygen in chambers of different hypoxia groups.
The rats were taken out of the chambers when hypoxia exposure was finished. The rats were anesthetized with 20% urethane (5 ml/kg) by intraperitoneal injection. The blood samples were collected with a carotid arterial catheter. 2 ml blood was placed in an anticoagulant tube with 3.8% sodium citrate, and then centrifuged at low temperature to collect platelet-rich plasma for flow cytometry. The serum was obtained by centrifugation of remain blood sample and stored in the –80°C refrigerator for subsequent ELISA analysis. The fresh thoracic aorta was separated from rats used to investigate the ultrastructural changes of endothelial cells by transmission electron microscopy. Endothelial cells were isolated from the thoracic aorta and the protein was extracted for West Blot analysis.
ELISA test
The protein concentration of ET-1, ICAM-1, VWF, tPA and PAI1 in serum were measured using ELISA Kit following the manufacturer’s instructions (Elabscience Biotechnology, Wuhan, China). All samples were measured in duplicate. The NO concentration was measured using NO test kit (Jiancheng Bioengineering institute, Nanjing, China) following the manufacturer’s instruction.
Western blot analysis
The lysate proteins from endothelial cells were separated on a 6% or 12% Tris-glycline SDS-PAGE and electrophoretically transferred onto nitrocellulose membranes. The membranes were blocked with 5% milk in Tris-buffered saline–Tween at room temperature for 2 h and then incubated with primary antibodies against NF-κBp65 (1:1000; Proteintech, Rosemont, USA), COX-2 (1:200; Proteintech, Rosemont USA), eNOS (1:1000; Cell Signaling Technology, Danvers, MA, USA), P-eNOS(1:1000; Cell Signaling Technology, Danvers, MA, USA) and β-actin (1:200; Boster Biological Technology, Wuhan, China) or GAPDH (1:1000; Cell Signaling Technology, Danvers, MA, USA) overnight at 4°C. The protein bands were detected by an immuno-chemiluminescence method. The density was quantitated by Bio-Rad Quantity One Software.
Transmission electron microscopy
About 1 mm square tissue from fresh thoracic aorta were primarily fixed in cold 3% glutaraldehyde for 2–4 h and then washed with Sorenson’s phosphate buffer solution (pH 7.4), then the tissue was post-fixed in 1% osmium tetroxide for 1 h. After three times washes with Sorenson’s phosphate buffer solution (pH 7.4), the samples were dehydrated in a series of acetone solutions and embedded in Epon 812. About 70 nm ultrathin sections were cut using ultramicrotome. These ultrathin sections were stained with uranyl acetate and lead citrate and they were assessed using a HT7700 (Hitachi, Tokyo, Japan) transmission electron microscope.
Flow cytometer
Platelet-rich plasma (25ul) was placed in four centrifuge tubes, 10 μl fluorescein isothiocyanate (FITC) labeled anti-rat CD63 (Ebioscience, San Diego, CA), 5 μl APC labeled anti-rat CD62P (Abcam, Cambridge, MA), FITC-IgG antibody and APC-IgG antibody (BD Biosciences, Franklin Lakes, NJ) as negative isotype control were added to the tubes respectively. 200 μL of phosphate buffer saline (PBS) were added to each tube and incubated at 4°C for 30 minutes in the dark. The cells were centrifuged and washed two times with PBS. Finally, the platelets were suspended with PBS and 1% paraformaldehyde was added. The expressions of CD62P or CD63 were detected by a flow cytometer (CytoFLEX, Beckman Coulter, Inc, Chaska, MN, USA). 100,000 cells were collected and the percentage of platelets expressing CD62p or CD63 was calculated.
Echocardiography
A few indicators including cardiac output(CO), cardiac index(CI), left ventricular ejection fraction (LVEF), left ventricular systolic diameter (LVDs), left ventricular diastolic diameter (LVDs) and left ventricular fraction shortening(LVFS), which indicate the myocardial contractility of rats before and after the model was setup, were detected by echocardiography (Model number: GEvivid7, probe: 10MS, GE, Inc, USA).
Statistical analysis
All data were presented as arithmetic means±standard deviation (SD). Differences in data for the three groups were analyzed by one-way analysis of variance. The t-test was used for comparison between two groups. p < 0.05 was considered significant of the difference. All analyses were tested using 19.0 SPSS statistical software.
Results
Effect of hypoxia on serum levels of NO, ET-1, ICAM-1 and vWF
In order to confirm our hypoxia models were successfully setup, firstly some of the vascular contraction-related factors were measured. As shown in Table 1, hypoxia significantly decreased the level of NO but significantly increased the levels of ET-1 and vWF in serum (p < 0.05). It did not show any difference of the serum NO levels between the CIH group and the CCH group (p > 0.05). However, the serum levels of ET-1, ICAM-1 and vWF in CIH group were significantly higher than that in CCH group (p < 0.05). There was no difference in ICAM-1 between the CCH group and the NC group (p > 0.05).
The serum levels of NO, ET-1, ICAM-1 and vWF in three groups
The serum levels of NO, ET-1, ICAM-1 and vWF in three groups
All values are presented as arithmetic means±standard deviation (M±SD). n = 15. NC = normoxic control; CIH = chronic intermittent hypoxia; CCH = chronic continuous hypoxia. Compare to NC group, *P < 0.05, §P < 0.01; Compare to CIH group, #P < 0.05, &P < 0.01.
To evaluate the effect of two hypoxia models on endothelial function in rats, as showed in Fig. 2, the expression of COX-2, NF-κBp65, eNOS and P-eNOS in endothelial cells were detected by Western Blot. The expression of COX-2, NF-κBp65 were significantly increased in both hypoxia models compared to NC control group, but the expression of eNOS and P-eNOS were significantly decreased (p < 0.01) in hypoxia groups. However, there was no significant difference between CIH group and CCH group (p > 0.05).
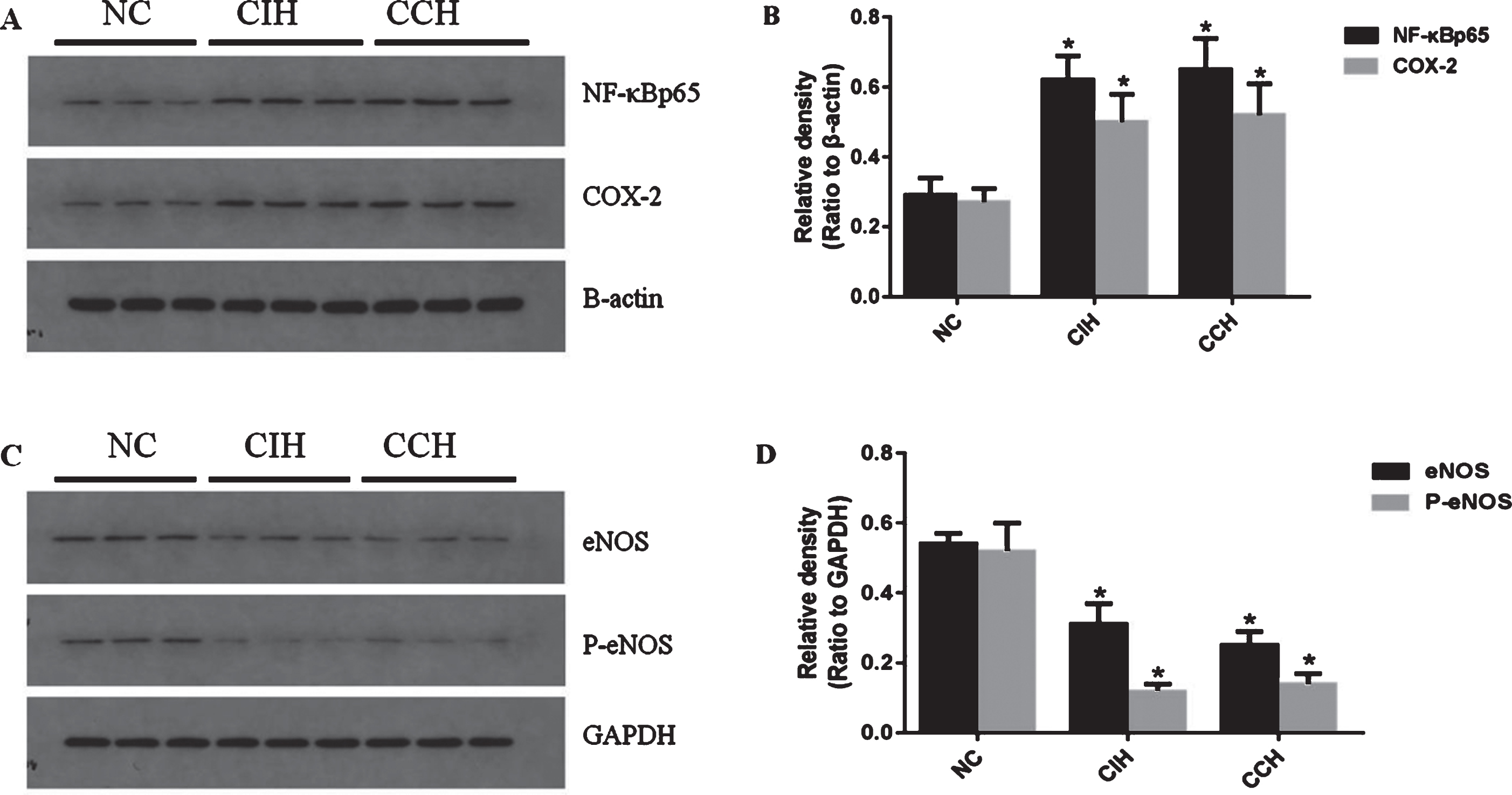
Different effects of CIH and CCH on endothelial function of rats. (A, C) Representative figures of immunoblots for NF-κBp65, COX-2, eNOS and P-eNOS. (B, D) Quantification of the protein levels of NF-κBp65, COX-2, eNOS and P-eNOS. Results are presented as M±SD; n = 15. *P < 0.01, compared to NC.
Transmission electron microscopy showed that the mitochondrial matrix particles decreased and partially disappeared, the mitochondrial edema, the mitochondrial crest become sparse, the cristae was expanded, and the cristae was swollen, some cristae was broken or disintegrated, and even some mitochondria were dissolved in the endothelial cells of thoracic aorta in CIH and CCH groups. However, the mitochondrial morphology and structure of endothelial cells in NC group were normal (Fig. 3).
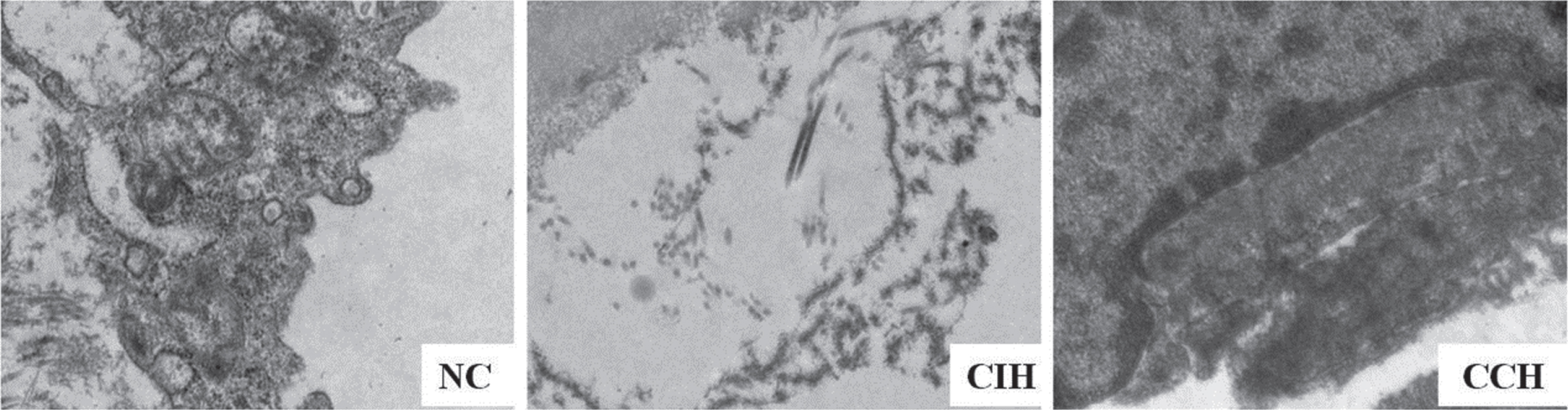
Representative pictures of transmission electron microscopy. Ultrastructural changes of endothelial cells in thoracic aorta were found the CIH and CCH groups (×10000, n = 5).
To assess the activation function of platelet, flow cytometry was performed to count the number of platelets expressing CD62p or CD63 in rats of the three groups. As showed in Fig. 4, CD62p positive platelets in the CCH group was significantly higher than in the CIH group and NC group (1.09±0.51% vs 0.42±0.12%, p < 0.05), but there was no significant difference between CIH group and NC group (p > 0.05). It did not show any statistical difference in platelets expressing CD63 among the three groups (p > 0.05).
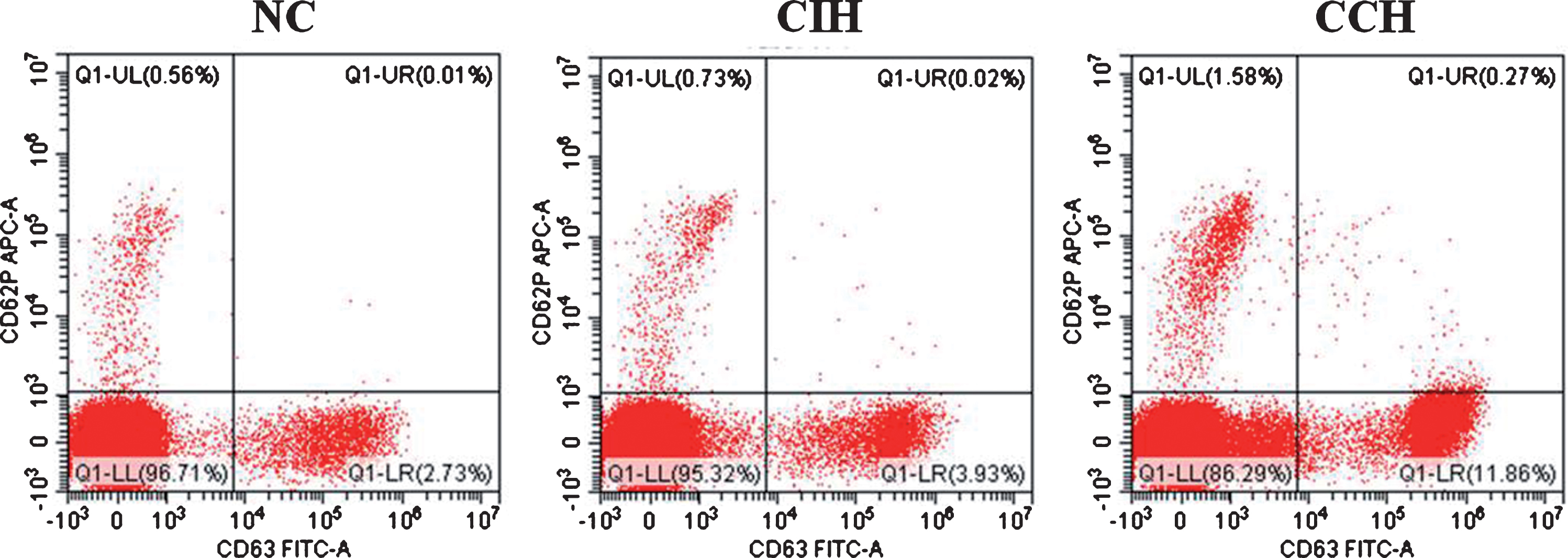
Effects of two hypoxia exposure on platelet activation of rats. The number of platelets expressing CD62p or CD63 were analyzed by flow cytometry analysis. The platelets was stained with monoclonals (anti-CD62p or anti-CD63). Data are representative of four independent experiments in each group.
As shown in Fig. 5A, both CIH group and CCH group had lower level of AT-III and higher levels of FIB and FVIII in serum than that of NC group (p < 0.01). However, it did not show any difference between the CIH group and the CCH group (p > 0.05) (Table 2). The serum level of PAI-1 in the CIH group and CCH group was significantly higher than that in NC group (p < 0.01). The serum level of t-PA in the CIH group was significant lower compared with the NC group and CCH group (p < 0.01), but there was no difference between CCH group and NC group (p > 0.05) (Fig. 5).
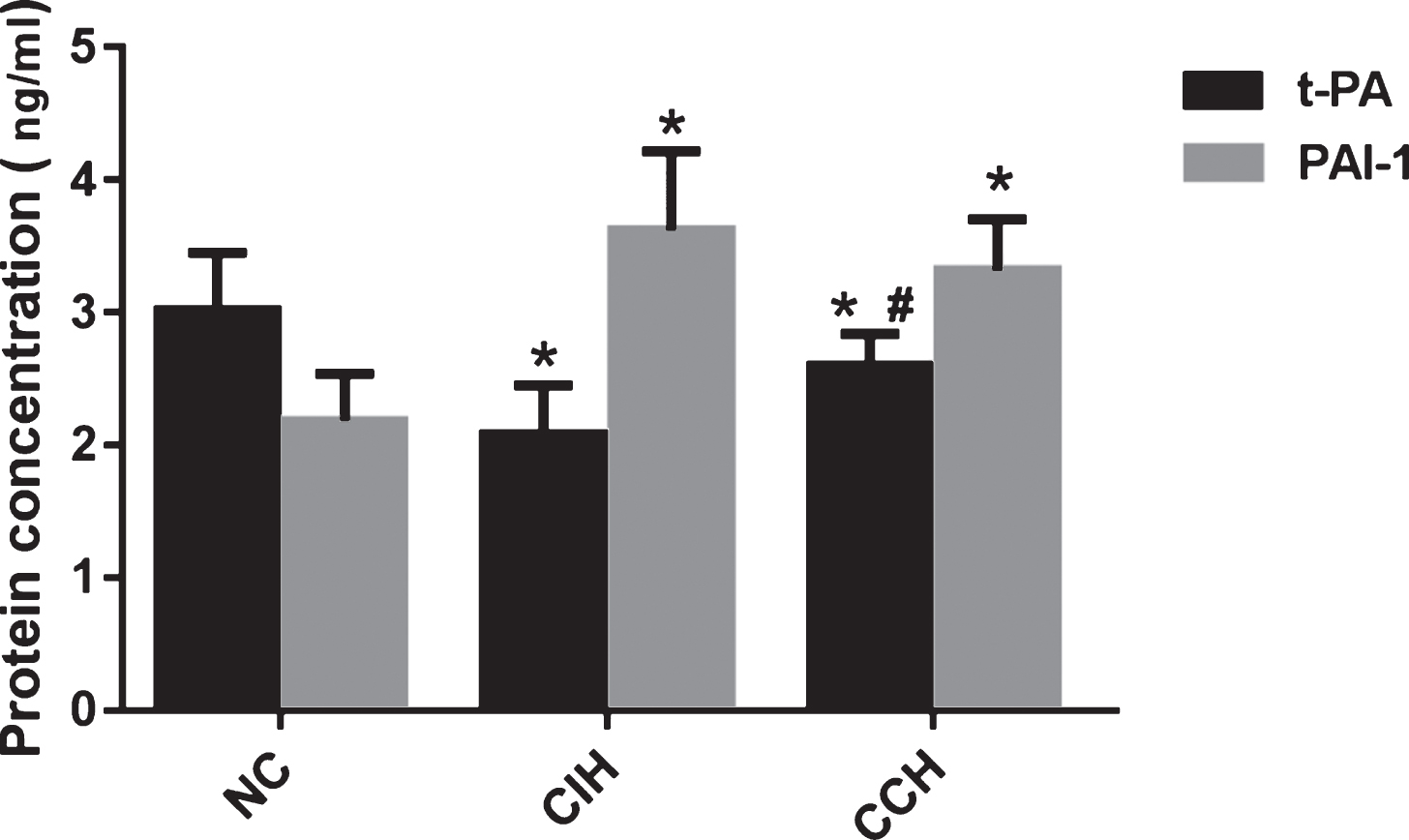
Effects of CIH and CCH on the t-PA and PAI-1 levels in serum. Results are presented as M±SD; n = 15. *P < 0.01 compared to NC. #P < 0.05, compared to CIH.
The plasma levels of FIB, FVIII and AT-III in three groups
All values are presented as arithmetic means±standard deviation (M±SD). n = 15. The plasma level of FIB and FVIII were determined with coagulation method. The plasma AT-III activity was determined using method of chromogenic substrate. Compare to NC group, *P < 0.01.
It has been directly evaluated that the myocardial contractile force changes of rats before and after the hypoxia models were setup in three groups using echocardiography. In NC group, left ventricular diastolic dimension (Dd) was significantly increased after modeling, but there were no significant differences between CIH group and CCH group (Data not shown). As expected, myocardial contractility indexes including CO, CI, LVEF and LVFS were significantly decreased after hypoxia exposure for 5 weeks (p < 0.01). However, there were no difference between CIH group and CCH group (p > 0.05) (Table 3).
Changes of myocardial contractility force before and after modeling in three groups
Changes of myocardial contractility force before and after modeling in three groups
All values are presented as arithmetic means±standard deviation (M±SD). n = 15. BM = before modeling; AM = after modeling; Compare to NC group, *P < 0.05, §P < 0.01; Compare BM with AM in each group, #P < 0.05, &P < 0.01.
The rat model of chronic exposure to CIH has been intensively used to mimics many aspects of the arterial hypoxemia that accompanies sleep apnea [27, 28]. The study by M McGuire et al using a rat model of CIH showed that chronic intermittent hypercapnic hypoxia caused systemic arterial and pulmonary hypertension, increased haematocrit and right ventricular hypertrophy, which mimics the nocturnal pattern of hypercapnic hypoxia of sleep-disordered breathing in humans [29]. In our present study, the rat models of CIH, CCH have been successfully established and the symptoms of rat models in the CIH and CCH groups were similar to the manifestation of patients with OSA and COPD. Consistent with the results of Marc M. Berger’s study that hypoxia markedly impairs vascular endothelial function in the systemic circulation in HAPE-S subjects due to a decreased bioavailability of NO [30], we found that the serum levels of NO, eNOS, P-eNOS and AT-III were significantly decreased when exposure to CIH and CCH, but the serum levels of ET-1, vWF, COX-2, NF-κB, FIB, FVIII and PAI-1 were significantly increased. Moreover, myocardial contractility indexes including CO, CI, LVEF and LVFS were significantly decreased after hypoxia exposure for 5 weeks. All these factors indicated that hypoxia, both CIH and CCH, induced the endothelia and cardiac system dysfunction. Furthermore, CIH and CCH caused ultrastructural damage to endothelial cells.
There is no significant difference between the effects of CIH and CCH on the endothelial eNOS and its synthesis product NO, which indicates that no difference in the effects of the two hypoxia models on endothelial cell diastolic function was existed. There is no significant difference in the effect of CIH and CCH on NF-κB inflammation pathway and its downstream product COX-2. However, CIH has a greater effect on vasoconstriction and vascular endothelial cell adhesion, and a stronger procoagulant effect, which indicated by the higher levels of ET-1, ICAM-1 and vWF in CIH group than CCH group. ET-1 is synthesized and secreted predominantly by vascular endothelial cells which has potent vasoconstrictor properties. Hypoxia induced the higher expression of ET-1 that occur during myocardial ischemia, which may contribute to the progression of heart failure [31]. Both types of hypoxia can cause endothelial dysfunction, but their severity are not the same. Hypoxia causes endothelial dysfunction and is likely to be responsible for one of those cardiovascular events.
Both CIH and CCH can induce hypercoagulability and reduce the function of fibrinolytic system, but the effect on platelet activation is not clear, which needs to be further studied. It has been shown that microcirculation are regulated by hypoxia and hypoxia affects all other parts of the blood, including white blood cells and thrombocytes as well as the non-cellular components [32]. Our previous study also showed that intermittent hypoxia and continuous hypoxia increase whole blood viscosity, impair the functions of red blood cells and promote the platelet aggregation in both CIH and CCH rat models. Moreover, CCH had a greater effect on blood rheology than CIH [33]. Hypoxia causes dysfunction of coagulation and fibrinolysis system, leading to thrombosis and early changes in atherosclerosis, which may be one of the mechanisms that cause cardiovascular events. Our results also indicated that CIH and CCH can cause the reduction of myocardial contractile function, suggesting hypoxia may be an independent risk factor for the decrease of myocardial contractility function, which is consistent with others studies [20, 23].
Compared with CCH group, CIH group had lower levels of t-AP and CD62P. t-PA has been known as a double-edged sword because it prevents HI-induced thrombosis in blood vessels, but unleashes a multitude of deleterious effects in the parenchyma of immature brains [34]. As a Ca2+-dependent receptor, CD62P stored in the alpha-granules of platelets and Weibel-Palade bodies of endothelial cells [35] and translocated to the plasma membrane and was released into the plasma in soluble form during platelet activation [36, 37], mediating the interaction of activated endothelial cells or platelets with leukocytes upon inflammatory and thrombogenic challenges. The different effects of CIH and CCH on t-AP and CD62 will be extensively studied in the future.
Conclusion
Our results suggested that CIH and CCH could cause endothelial dysfunction, dysfunction of the coagulation-fibrinolysis system and decreasing of myocardial contractility. Compared with CCH, CIH has greater effect on vasoconstriction and adhesion of vascular endothelial cells, and stronger procoagulant effect which were associated with cardiovascular events of OSA.
Conflict of interests
The authors have declared that there are no conflicts of interest.
Footnotes
Acknowledgments
This work was supported by research grant from the National Natural Science Foundation of China (81770089).