
Abstract
It has been reported that the beta-adrenergic receptor blocker (propranolol) and the a-adrenergic receptor (AR) blocker (phentolamine) both can inhibit human endothelial cell (EC) angiogenesis in vitro. However, it is unknown whether this inhibition also acts on pericytes. The present study aimed to determine how pericytes react to treatment with an a-/β- AR blocker. In the study, cell proliferation assays and scratch assay were performed to assess the effect of phentolamine or propranolol on cell proliferation and migration. Western blot and ELISA were employed to determine changes in VEGF-A and Ang-1 expression levels. The results indicated that the nonselective a-/β- AR blocker inhibited the proliferation, migration, and secretion of pericytes. The use of the nonselective a-/β- AR blocker might have an impact on vascularization and vascular maturation. Our research suggests the rational use of nonselective a-/β- AR blockers to treat angiogenesis-dependent diseases.
Abbreviations
AR, adrenergic receptor; EC, endothelial cell; VEGF, Vascular endothelial growth factor; ANG, angiopoietin; TIE-2, tyrosine kinase receptor-2; PTSD, posttraumatic stress disorder; BBB, blood-brain barrier
Introduction
Angiogenesis is necessary for physiological growth, such as wound healing; the menstrual cycle and reproduction; pathological conditions, such as cancers, chronic degenerative disorders; and various ischemic and inflammatory diseases [1]. Thus, physiological angiogenesis might be the key to treat many significant human diseases [2–4]. In that context, determining the effects of adrenergic receptor (AR) antagonists on the vascularization process during treatment could lead to more effective therapeutic strategies [5]. Angiogenesis begins with the proliferation, migration, and differentiation of endothelial cells (ECs) [6]. Angiogenesis is controlled by the balance of pro-angiogenic and anti-angiogenic factors. Shear stress can affect EC function, inflammatory reactions, and the development of venous disturbances [7, 8]. Decreased shear stress could lead to the upregulation of angiogenesis [9]. Vascular endothelial growth factor (VEGF) is an important pro-angiogenic factor that induces EC proliferation, migration and new vessel formation [10]. VEGFA is a dimeric disulfide-bound glycoprotein that attracts circulating neutrophils and monocytes, and induces microvascular hyperpermeability by disorganizing the endothelial junction proteins [11]. VEGFA and its receptors on ECs (VEGFR1 and VEGFR2) can also increase lumen formation and increase vessel length [12–14].
Meanwhile, angiopoietin (ANG) growth factors can bind to endothelial tyrosine kinase receptor-2 (TIE2) receptor tyrosine kinase to form the ANG-TIE2 signaling complex, which acts on the vasculature to control blood vessel development and vascular remodeling. It was reported ANG1 participates in the stability of newly formed vessels, the normalization of vessel function in various murine disease models, and alleviates oxygen-induced pathological changes (such as retinopathy, choroidal neovascularization, and diabetic nephropathy) [15]. Furthermore, ANG binds to integrin family members and induces focal adhesion kinase phosphorylation, which promotes cell migration and sprouting angiogenesis [4]. Thus, ANG acts as a direct pro-angiogenic factors. Moreover, ANG1 increases EC survival and stabilizes them against pathological insults. In addition, TIE2 expressed by ECs can be activated by pericyte-expressed ANG1.
ARs play an important role in angiogenesis. Propranolol is a nonselective β-adrenoreceptor antagonist used to treat hypertension, pheochromocytoma, infantile hemangioma, and posttraumatic stress disorder (PTSD) [16–19]. In addition. the non-selective a-AR blocker, phentolamine, was first investigated as a potential treatment for hypertension. Recently, phentolamine has also been used to treat many vascular diseases, such as impotence, erectile dysfunction, neurogenic pulmonary edema, and Raynaud’s syndrome [20–23]. Activation of the β-adrenoreceptor increased the synthesis and release of VEGF, interleukin (IL)8, and IL6, and leading to a significant increase in tumor vascularization [24]. Propranolol was reported to inhibit EC proliferation, migration, and lumen formation. Recently, microvascular ECs were found to express a-ARs, and phentolamine, as an a-AR blocker, could also inhibit EC angiogenesis in vitro, similar to the non-selective beta-AR blocker [25]. However, most studies of angiogenesis related to ARs have focused on EC, and ignored the importance of pericytes.
Pericytes are described based on their prominent location at the periphery of microvessels and are required for the formation of stable vascular networks. They can communicate with other cells and regulate microcirculatory function [26, 27]. By penetrating the basement membrane to contact ECs and release growth factors, pericytes contribute to the regulation of angiogenesis, EC survival, and the clearance and phagocytosis of cellular debris. Moreover, the extent of pericyte coverage is an important marker of vessel maturation. They also help to maintain the blood-brain barrier (BBB) and regulate microvascular flow [28–30]. This study aimed to determine the effects of a-/β- AR blockers on pericytes, which will help us to better understand the vascularization process.
Material and methods
Cell culture
Primary pericytes (Human Brain Vascular Pericytes, HBVPs) were purchased from Sciencell Research Laboratories (Carlsbad, CA, USA) and maintained in Pericyte Medium (Sciencell Research) supplemented with 2% fetal calf serum (FCS), 1% of the corresponding pericyte growth supplement, and 1% penicillin/streptomycin. Cells were cultured at 37°C in a humidified atmosphere containing 5% CO2.
Cell proliferation assays
HBVPs were seeded into 96-well plates at a density of 5000 cells/well in 100 μL of pericyte medium, and then incubated at 37°C under a humidified atmosphere containing 5% CO2. The old medium was removed after 24 h and replaced with 100 μL of fresh medium containing phentolamine at different concentrations (0, 0.5, 6, 10, and 14 μg/ml) (Sigma-Aldrich, St. Louis, MO, USA) or propranolol at different concentrations (0, 50, 100, 150, and 200 μmol/L) (Sigma-Aldrich). After 48 h, 10 μL of cell counting kit 8 (CCK-8) dye and 90 μL pericyte medium was added to each well, and the cells were incubated at 37°C under a humidified atmosphere containing 5% CO2 for 2 h. The absorbance of the cells in the wells was then detected at 450 nm using a Multiskan GO microplate reader (Thermo Fisher Scientific, Waltham, MA, USA).
Scratch assay for cell migration
HBVPs were plated in triplicate on 24-well plates at a density of 1×104 cells/well. When the cells reached 100% confluence, a 1 mm-wide gap was scratched in the center of the cell layer using a 100 μL pipette tip. Dead cells were washed away using phosphate buffered saline (PBS), and new medium containing different concentrations of phentolamine (0, 0.5, 6, 10, and 14 μg/ml) or propranolol (0, 50, 100, 150, and 200 μmol/L) was added. The scratched area of each cell layer was photographed (40×) at 0, 12, 24, and 48 h under a Nikon Eclipse phase contrast microscope (Nikon, Tokyo, Japan) with a Hamamatsu digital camera (Hamamatsu-city, Japan), and Improvision software (Perkin Elmer, Waltham, MA, USA)
Western blotting
Cells were lysed in Radioimmunoprecipitation assay (RIPA) lysis buffer (G2002, Beyotime Biotechnology, Jiangsu, China). Proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and subsequently transferred to polyvinylidene fluoride (PVDF; Millipore, Billerica, MA, USA) membranes. The membranes were blocked with 5% nonfat dry milk in Tris-buffered saline with Tween 20 (TBST) at room temperature for 1 h, followed by incubation with the corresponding primary antibodies, including anti-VEGFA (PAB12284, Abnova, Taipei, Taiwan), anti-ANG1(bs-0800 R, Bioss Antibodies Inc., Woburn, MA, USA), and anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (2118, Cell Signaling Technology, Danvers, MA, USA) overnight at 4°C. The membranes were washed with TBST three times for 5 min each time, and then incubated with the corresponding secondary antibodies for 30 min (SA00001-2, GB23301, ProteinTech, Rosemont, IL, USA). Following three washes with TBST for 5 min each time, enhanced chemiluminescence A (ECLA) and ECLB were mixed in an equal volume and applied to the PVDF membrane. After 1–2 minutes, the residual liquid was removed and the PVDF membrane was placed into a C-DiGit Blot Scanner (LI-COR Biosciences, Lincoln, NE, USA). The exposure time was adjusted according to different light intensities.
Enzyme-linked immunosorbent assay (ELISA)
ELISA was used detect intracellular VEGF-A and ANG1 levels, and their levels in cell supernatants. Cells were seeded into 6-well plates at a density of 9×104 cells/well, and cultured with 2 mL of complete medium at 37°C under a humidified atmosphere containing 5% CO2. When the cells reached a density of 70%, the old medium was replaced with 2 mL of fresh complete medium containing different concentrations of propranolol (100 μM) and phentolamine (6 μg/ml). After incubation at 37°C under a humidified atmosphere containing 5% CO2 for 48 h, a human VEGF-A ELISA kit (SEA143Hu, Cloud-Clone Corp., Katy, TX, USA) and an ANG1 ELISA kit (SEA007Hu, Cloud-Clone Corp.) were used according to the manufacturer’s instructions.
Statistical analysis
Statistical analysis was performed using one-way analysis of variance (ANOVA) with Dunnett’s post hoc test to compare the experimental groups with the control group. All statistical analyses were performed using SPSS software (IBM Corp., Armonk, NY, USA). P < 0.05 was considered statistically significant.
Results
Phentolamine and propranolol inhibit the proliferation of pericytes
Our previous experiments showed that the phentolamine could significantly inhibit the proliferation of human dermal microvascular endothelial cells (HDMECs) and human brain microvascular endothelial cells (HBMECs) in a dose-dependent manner, with a half maximal inhibitory concentration (IC50) of 30 μg/mL for HBMECs and 50 μg/mL for HDMECs.
In the present study, we observed the effect of phentolamine and propranolol on the proliferation of HBVPs (pericytes). The CCK-8 cell proliferation assay showed that phentolamine inhibited HBVP proliferation at very low drug concentrations and acted in a dose-dependent manner. At 6 μg/mL, phentolamine inhibited HBVP proliferation significantly. However, although propranolol also inhibited HBVP proliferation, a higher concentration was required to achieve significant inhibition (13 μg/mL (100 μM) (propranolol, Mw = 259.343) vs. 6 μg/mL of phentolamine; Fig. 1).
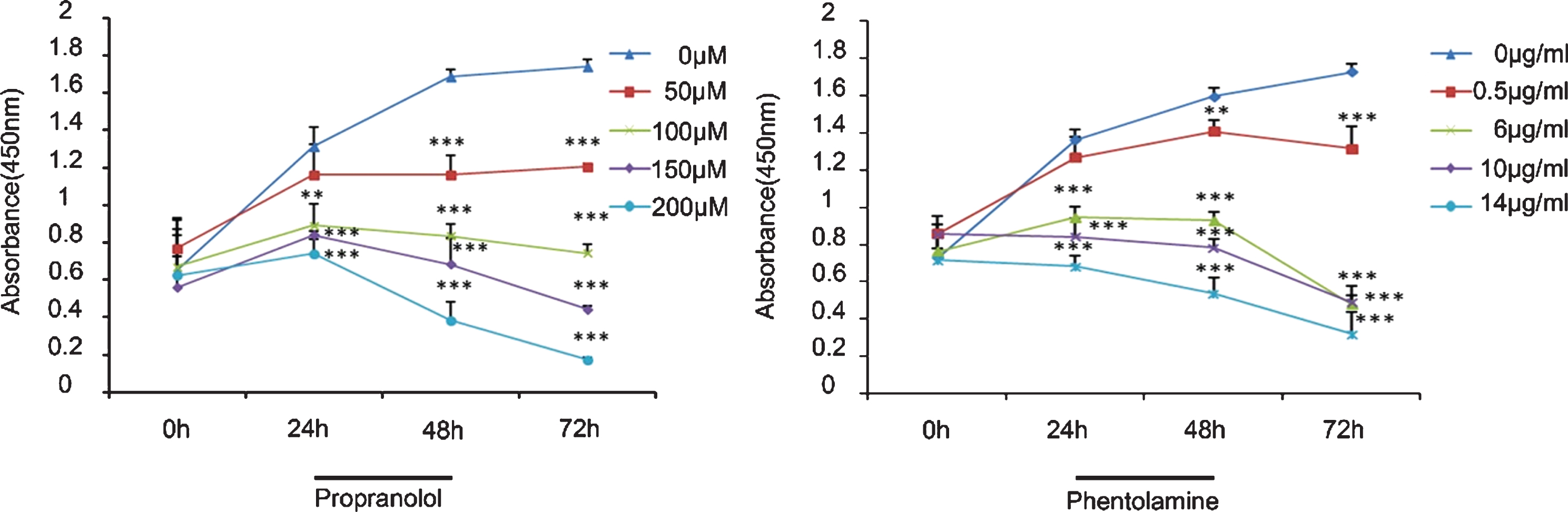
Effect of phentolamine and propranolol on pericyte proliferation. Optical density (OD) values were measured at 450 nm at 0, 24, 48, and 72 h. Both phentolamine and propranolol inhibited cell proliferation. The asterisks (*) on the figure show the difference in proliferation between the groups at each time point. One-way analysis of variance (ANOVA) with Dunnett’s post hoc test was used. For propranolol, the difference between 0 μM vs. 50 μM was not significant (P > 0.05), whereas the differences were significant for 0 μM vs. 100 μM (P < 0.05), 150 μM (P < 0.01), and 200 μM (P < 0.001). For phentolamine, the difference between 0 μg/mL vs. 0.5 μg/mL was not significant, but was significant for 0 μg/mL vs. 6 μg/mL (P < 0.05), 10 μg/mL (P < 0.05), and 14 μg/mL (P < 0.01). The error in this analysis was most likely caused by differences in the number of plated cells and cell viability. However, the inhibition induced by both chemicals was dose-dependent. Phentolamine seemed to have a stronger inhibitory effect. *P < 0.05; **P < 0.01; ***P < 0.001).
The scratch wound-healing assay was used to observe the migration of the HBVPs from a monolayer under treatment with different concentrations of phentolamine and propranolol. A low concentration of phentolamine (0.5 μg/ml) induced non-significant inhibition of HBPV migration. HPBV migration was significantly inhibited by phentolamine at 6 μg/ml, which was far lower than the concentration required to significantly inhibit the migration of ECs (10 μg/mL or 20 μg/mL). However, propranolol showed a significant inhibitory effect on HPBV migration only at concentrations of 39 μg/mL (150 μM) and above (Fig. 2).
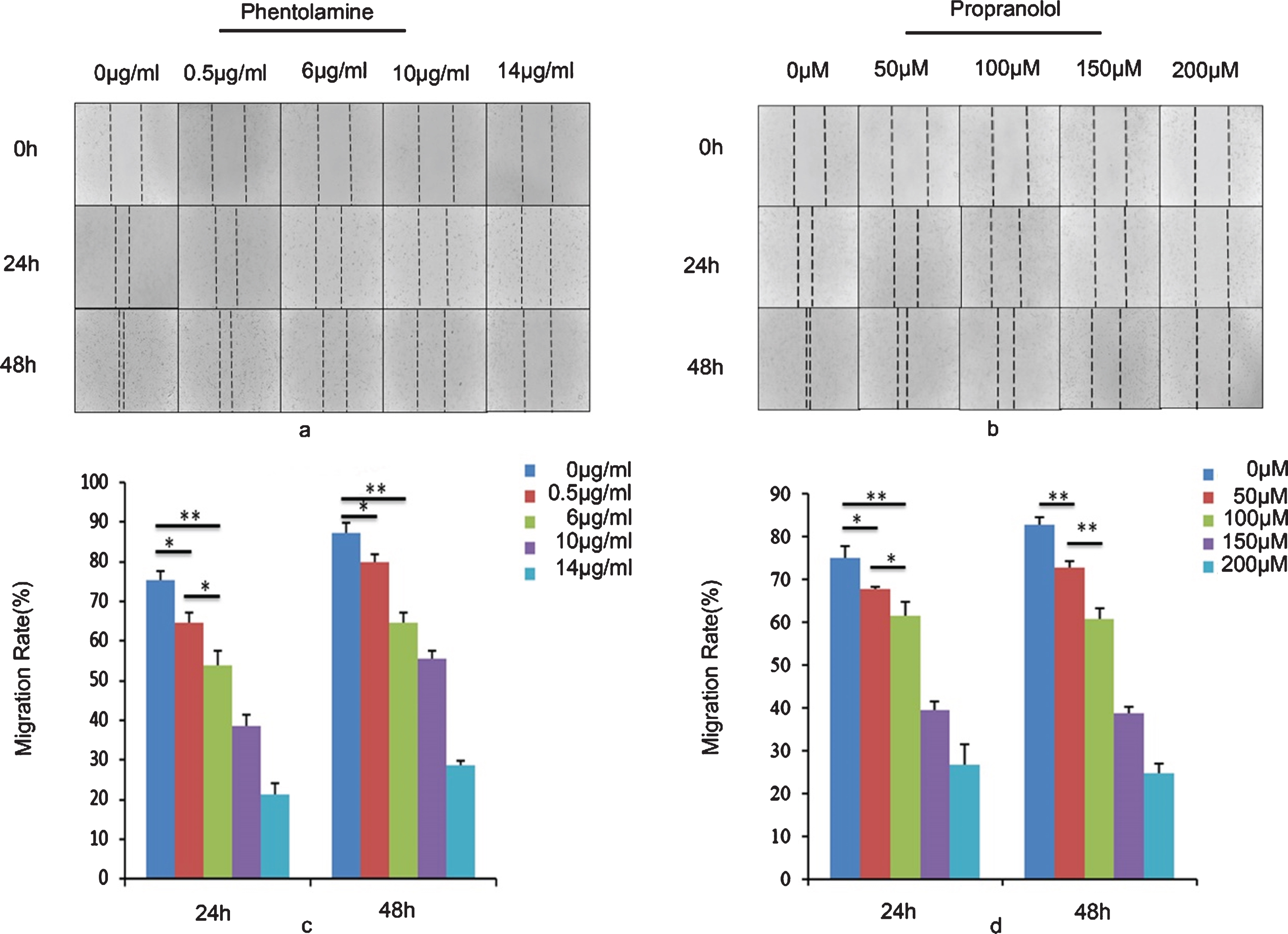
Effect of phentolamine and propranolol on cell migration. Both phentolamine and propranolol could delay the scratch wound closure of pericytes in a dose-dependent manner. (a) and (b) are images of the control (0 μM and 0 μg/mL), and phentolamine and propranolol treated cells, respectively. (c) and (d) show the percentage of the wound closure area of pericytes treated with phentolamine and propranolol at different concentrations at 24 h and 48 h. The mean of three identical and independent experiments±the standard deviation (SD) is shown *P < 0.05; **P < 0.01; ***P < 0.001). According to the result at 48 h, phentolamine seemed to have a stronger inhibitory effect.
Western blotting and ELISA were used to determine the effects of phentolamine and propranolol on the levels of VEGFA in HBVPs in vitro. As shown in Fig. 3, treatment with phentolamine and propranolol could reduce VEGFA levels. Moreover, compared with the control group (0 μg/mL), when the HBVPs were treated with phentolamine at 6 μg/mL for 48 h, the level of VEGF-A decreased dramatically (by 43% compared with that of the control group: 19.53 pg/mL for the control group vs. 11.11 pg/mL for the phentolamine group). Treatment with propranolol also decreased the VEGFA level; however, treatment with propranolol at 100 μM for 48 h did not produce such a dramatic decrease in VEGFA compared with that induced by phentolamine at 6 μg/mL for 48 h (decrease of 24%; 19.93 pg/ml for the control group vs. 15.10 pg/mL for the propranolol group; Fig. 3 (a–e)).

Vascular endothelial growth factor A (VEGFA) and angiopoietin 1 (ANG1) levels in pericytes. (a,b) VEGFA levels in pericytes as assessed using western blotting. The gray values were standardized to allow comparisons between the groups. (c,d) The decrease in VEGFA levels of pericytes under 100 μM propranolol and 6 μg/mL phentolamine at 0, 24, 48 and 72 h (standardized). (e) Effect of propranolol and phentolamine on VEGFA levels in the cells. (f) ANG1 levels in pericytes. Pericytes were treated with 100 μM propranolol and 6 μg/mL phentolamine for 24, 48, and 72 h, and the levels of VEGFA and ANG1 were detected using western blotting. The gray values were standardized to allow comparisons between the groups. (g) The decrease in ANG1 levels of pericytes under 100 μM propranolol and 6 μg/mL phentolamine at 0, 24, 48, and 72 h (standardized). (h) Effect of propranolol and phentolamine on ANG1 levels in the cells.
The western blotting and ELISA were used to determine the effects of phentolamine and propranolol on the levels of ANG1 in HBVPs in vitro. The ANG1 level decreased significantly under phentolamine and propranolol treatment. However, at 48 h, propranolol at 100 μM could significantly inhibit the level of ANG1, whereas, at the same time point, phentolamine at 6 μg/mL could not (Fig. 3 (f–h).
Discussion
Pericytes are a type of perivascular supporting cell, being present at intervals along the walls of pre-capillary arterioles, capillaries, and post-capillary venules [31, 32]. They directly contact with ECs, cover 30–90% of the micro vessel wall with their processes, and utilize a common basement membrane with ECs [33, 34]. Pericytes can control blood flow physiologically, and the relaxation of contractile pericytes is the main reason for the dilation of capillaries to increase flow [35, 36]. Pericytes can promote endothelial sprouting, and when pericytes are impaired or lose their function, the formation and function of vessels is inhibited [37].
Vascular development is a relatively complicated process, which begins with angiogenesis and ends up with functionally mature vessels. For many vascular-related diseases, providing operational vessels or maintaining normal vessel function are the goals of therapy. Many studies have examined the effects of receptor agonists on ECs [38–40]. However, relatively few have evaluated the relationship between AR-antagonists and pericytes. It is believed any aberrant or failed interactions between pericytes and proliferating ECs will result the death of ECs and pericytes, and the inhibition of angiogenesis. Pan’s study found that the non-selective a-AR antagonist phentolamine could inhibit EC angiogenesis in vitro, as could propranolol [25]. In the present study, we investigated the effects of AR antagonists on pericytes. We treated pericytes with the AR antagonists phentolamine and propranolol and detected the levels of VEGFA and ANG levels after treatment. The results showed that phentolamine had a stronger inhibitory effect on the proliferation and migration of pericytes, and propranolol had a stronger inhibitory effect on the secretion of pericyte-derived ANG1.
Pericytes can help to stabilize the vessel wall and prevent vascular leakage, and their loss or detachment is implicated in several diseases [33, 41]. A previous study revealed a correlation between pericyte numbers and vessel stability [42]. Moreover, pericytes can suppress EC proliferation and migration [43, 44]. However, vessels lacking pericytes were prone to regression in vivo [45]. The inhibition of pericyte proliferation would lead to low numbers of pericytes at their sites of activity and might lead to neovascular regression, which should be investigated in a future study. Pericyte migration might delay of the maturation of vessels; however, this speculation also requires further study.
The formation of the endothelial plexus occurs earlier than the recruitment of pericytes. When the new vasculature is formed it lacks a pericyte coating, which provides a window of opportunity for the vasculature to remodel and adjust to the physiological needs of the tissue [14]. In the present study, phentolamine showed stronger inhibition of pericyte proliferation and migration than propranolol; therefore, application of the most suitable drug at the appropriate time might help to gain a demand vascular structure.
VEGFA acts on the pericytes in an autocrine and paracrine manner to stimulate their proliferation and migration [14]. In addition, the VEGFA released by pericytes might promote EC survival during angiogenesis [46]. Evidence suggests that immature and developing vessels depend on tissue-derived VEGF, and exogenous VEGF can prevent the apoptotic death of ECs [47]. ANG1 is an essential vascular regulatory molecule that mediates the interactions of ECs and pericytes, and might be involved in promoting the maturation and stabilization of the micro-vasculature by tightening endothelial junctions and recruiting pericytes. It also was reported the ANG1 can reduce the permeability of vessels via the endothelial receptor (TIE2) [48, 49].
In this study, we found both the nonselective a-/β- AR blocker could reduce VEG-A levels, with phentolamine having a stronger inhibitory effect. Pericytes might play an important role in vessel stability via pericyte-derived factor VEGF, because a decrease in pericyte-derived VEGF resulted in selective EC death [45–47]. The use of nonselective a-/β- AR blockers, especially phentolamine, might not only inhibit the proliferation, migration, and lumen of ECs, but also could act on pericytes to decrease neo-capillary stability and cause ECs death, which would further increase phentolamine-induced inhibition of ECs.
We also found that both nonselective a-/β- AR blockers could decrease pericytes’ ANG1 levels, with propranolol having a more obvious effect. The wound-healing experiment indicated the ANG1 might be involved in GM-CSF modulation of endothelial intracellular junctions and is a key regulator of vascular integrity that mediates GM-CSF function in micro-vascular leakage. It is believed that ANG1 is a potent anti-leakage cytokine that can restore poorly remodeled and leaky vessels [50]. Furthermore, VEGFA was reported to elevate ANG1 expression in pericytes or smooth muscle cells [51, 52]. Meanwhile, ANG1 was suggested to inhibit the effects of multiple permeability-enhancing agents, including VEGF [50, 53]. How the interaction between VEGFA and ANG1 affects the vascular-related diseases requires further research.
In short, the intercellular interaction between ECs and pericytes is very important for blood vessel maturation. We believe that the nonselective a-/β- AR blockers affect angiogenesis by acting directly on ECs and indirectly on pericytes.
Conclusion
The present study found that the AR antagonists could also act on pericytes, which are involve in regulating EC function, and might affect angiogenesis and regulating vessel mature and normal EC function. However, further experiments are required to better understand the comprehensive effects of the AR receptor antagonists on ECs and pericytes in vivo and in vitro.
Conflict of interest
The authors declare and certify that they have no financial or non-financial conflicts of interests.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81671924 and 81272105); the Science and Technology key Project of Guangdong province (No. 2014B020212010); the Health and Medical Treatment Collaborative Innovation Major Special Projects of Guangzhou (No. 201508020253); the Science and Technology Planning Project of Guangdong Province of China (No. 2015B020233012); the National Key Research and Development Plan of China (No. 2017YFC1103301); and Military Medical Innovation Special Projects (No. 18CXZ029).