
Abstract
INTRODUCTION:
Stromal-epithelial interactions are fundamental for normal organ development and there is a multitude of evidence that the different components of the microenvironment are also necessary for the maintenance and promotion of the “tumor organ”. Deregulated tumor associated extracellular matrix (tECM) is a hallmark of cancer, causing an alteration in the amount and composition of the different components (i.e. proteins, proteoglycans, glycoproteins and polysaccharids) of the ECM. As epithelial-stromal interactions are reciprocal, it is possible that tECM itself is able to initiate tumor development. We therefore established a mouse model to examine the influence of tECM of murine breast cancer on developing breast tissue in mice.
MATERIALS AND METHODS:
Breast cancer was established in 5 BALB/c mice by subcutaneous injection of 1×106 4T1 cells in 100μl PBS into the left mammary fat pad. The mammary fat pad including the primary tumor was excised after two weeks, decellularised and labelled as tumor extracellular matrix (tECM). Tumor ECM of 4T1 tumors was implanted into the 4th inguinal mammary fat pad of BALB/c mice (n = 5) aged 5 days. After 12 weeks the fourth mammary fat pad including the primary tumor was excised. Tissue was used for paraffin embedding and mouse breast cancer PCR array. Murine breast cancer tissue (BCT) and normal murine breast tissue (BT) served as control.
RESULTS:
Gene array analysis of 84 breast cancer-specific transcripts revealed that the mammary gland cells which were exposed to tumor ECM (tECM-BT) showed a similarly high overexpression for 22 genes as apparent for breast cancer tissue (BCT). The corresponding scatter plot showed a high agreement in the expression of the examined genes between the mammary gland cells which were exposed to tumor ECM and the breast cancer tissue.
DISCUSSION:
Our results clearly demonstrate that the tECM is able to shift the gene expression pattern of murine mammary epithelial cells towards that of carcinoma, indicating a role in breast cancer initiation. These data underlines that the acellular component of the tumor (ECM) can lead to a transformation of mammary gland tissue cells. These data show for the first time that the interaction of normal breast tissue cells with tumor ECM leads to an exchange of information and a consecutive overexpression of tumor-specific genes.
Introduction
Carcinoma was long seen as a disease solely caused by genetic and epigenetic alterations in proto-oncogenes and/or tumor suppressor genes of epithelial cells [1]. However, emerging research suggests that epithelial tumors are more than a mere accumulation of genetically dysregulated carcinoma cells. Both, normal and malignant epithelial cells live in a complex microenvironment, composed of stromal cells and extracellular matrix. Stromal-epithelial interactions are fundamental for normal organ development and there is a multitude of evidence that the different components of the microenvironment are also necessary for the maintenance and promotion of the “tumor organ” (reviewed in [2–4]). Cancer associated fibroblasts (CAFs), the predominant cells of the tumor stroma have an active role in various steps of carcinogenesis. In fact, there are indications that active fibroblasts even are able to initiate tumor development by triggering genetic and epigenetic alterations in epithelial cells. However, the exact nature of the factors, mediating this initiating process is mainly unknown. Besides the secretion of signal molecules CAFs are mainly responsible for the production of the tumor associated extracellular matrix (tECM). Deregulated ECM dynamics is a hallmark of cancer, causing an alteration in the amount and composition of the different components (i.e. proteins, proteoglycans, glycoproteins and polysaccharids) of the ECM. This leads to unique physical, biochemical and biomechanical properties that promote tumor development. As epithelial-stromal interactions are reciprocal, it is possible that tECM itself is not only a requirement for tumor maintenance but also able to initiate tumor development. We therefore established a mouse model to examine the influence of tECM of murine breast cancer on developing breast tissue in mice. Our results clearly demonstrate that the tECM is able to shift the gene expression pattern of murine mammary epithelial cells towards that of carcinoma, indicating a role in breast cancer initiation.
Materials and methods
Mice
Two-month-old female BALB/c mice were purchased from Charles River Laboratories (Germany) and housed in a specific pathogen-free facility at the University of Regensburg. All experiments were performed following the guidelines of Veterinary Medicine at the University of Regensburg and according to the guidelines of the Journal for Clinical Haemorheology and Microcirculation [5].
Tumor cell lines
Murine breast cancer cell line 4T1 (ATCC 2539) was cultured in RPMI 1640 medium (Invitrogen) supplemented with 10% fetal bovine serum (FBS) (Invitrogen), L-glutamine (Sigma), and penicillin-streptomycin (Sigma) at 37°C in a 5% CO2-containing incubator.
Generation of tECM of murine breast cancer
Breast cancer was established in 5 BALB/c mice by subcutaneous injection of 1×106 4T1 cells in 100μl PBS into the left mammary fat pad. Tumors reached the average diameter of 10 mm (∼100 mm3 in volume) after two weeks and mice were anaesthetized for tumor tissue harvesting using Ketamil (10 mg/mL)/Xylazil (0.9 mg/mL). The mammary fat pad including the primary tumor was excised and all mice were euthanized after the procedure.
Decellularization
Samples were frozen at – 80°C and processed following the protocol previously described [6]. Briefly, tumor tissues were re-frozen at – 80°C and thawed at room temperature for 3 cycles, and then rinsed in ultrapure water for 2 days at room temperature with agitation (120 rpm). These samples were then treated with 0.5 M NaCl for 4 h, which was followed by 1 M NaCl for 4 h, and washed in ultrapure water overnight; this saltwater wash procedure was repeated. After being treated with 0.25% trypsin/EDTA for 2 h and rinsed in deionized water for 1 h, samples were processed with isopropanol (IPA) overnight. The samples were then treated with 1% Triton X-100 for 3 days (1 change daily), washed in ultrapure water for 2 days (3 changes daily), and rinsed in phosphate-buffered saline (PBS) for 1 day. The tissue was cell free after the whole procedure and labelled as tumor extracellular matrix (tECM).
Sterilisation
The tECM samples were sterilised with a solution of 1% peracetic acid (Wofasteril; Kesla)/ 24% Ethanol (Sigma Aldrich) in aqua ad injectabilia for 4 h by vigorous shaking at a pressure below 200 mbar [7–9].
Generation of breast tissue under the influnce of tECM
Tumor ECM of 4T1 tumors was implanted into the 4th inguninal mammary fat pad of BALB/c mice (n = 5) aged 5 days. After 12 weeks mice were anaesthetized using Ketamil (10 mg/mL)/Xylazil (0.9 mg/mL) and the fourth mammary fat pad including the primary tumor was excised.
Staining
Tissue was fixed in 4% paraformaldehyde (PFA) for 1 h and then dehydrated, embedded in paraffin and sectioned (4μm sections). Sections were then dewaxed, rehydrated and processed with the following standard staining protocols: haematoxylin and eosin (H&E), Masson’s trichrome (for collagen) and actin [10, 11].
H&E staining
Sections were immersed for 4 minutes in Harris haematoxylin solution (Sigma-Aldrich), rinsed in water and counterstained for 4 minutes in eosin (Carl Roth GmbH). After rinsing with deionized water, samples were subsequently dehydrated in an ascending alcohol series. The sections were embedded with Entellan (Merck) and analyzed by microscope (Axioskop 40 FL, Carl Zeiss). Photos of the sections were taken using an Olympus camera XC30 (Olympus Soft Imaging Solutions).
Masson’s trichrome
Paraffin embedded 4-μm sections were stained with Masson’s trichrome reagent to demonstrate collagen. First sections were fixed in Bouin’s liquor overnight and washed next day in running water until the yellow color disappeared. Afterwards slides were rinsed twice with distilled water and stained with Mayer’s haematoxylin for 5 minutes. Slides were then placed in 0.5% hydrochloric acid in 70% alcohol for 5 seconds, washed in running tap water for 30 seconds and rinsed twice with distilled water. Slides were stained with acid ponceau for 10 minutes and rinsed three times with distilled water and dissolved in 1% phosphomolybdic acid aqueous solution. Staining with aniline blue was performed for 5 minutes and afterwards dissolved in 1% glacial acetic acid for 5 minutes. Slides were than dehydrated in 95% ethyl alcohol several times, followed by anhydrous alcohol, hyalinized with dimethylbenzene, and sealed with neutral balsam. Collagen fibers appeared blue and remaining nuclei after decellularization black.
Actin
Deparaffinized sections were washed with Tris-buffered saline (TBS: 0.05 M Tris, 0.015 M NaCl, pH 7.6), before being incubated with protease-free donkey serum (5% diluted in TBS with 0.1% Triton-X 100 for cell permeabilization). The primary antibody (actin: rabbit-anti-mouse 1:50, abcam) was diluted in 10 mM PBS containing 0.1% BSA, 0.3% Triton X-100 and the sections were incubated overnight with the prepared primary antibody for 1 hour at room temperature and subsequently at 4°C overnight [12]. The following day, the sections were washed 3 times in PBS containing 0.1% BSA, 0.2% gelatine and 0.05% Saponin followed by incubation with a secondary antibody (horseradish peroxidase conjugated goat anti-rabbit immunoglobulins; Dako) diluted in PBS with 0.1% BSA and 0.3% Triton X-100 for 1 h. After additional washing for 3 times with PBS containing 0.1% BSA, 0.2% gelatine and 0.05% Saponin, peroxidase activity was detected (after 10 minutes of incubation) using diaminobenzidine (DAB) in a final concentration of 1 mg/mL. Then sections were rinsed in PBS for 3×10 minutes, twice in ultrapure water and then counterstained in Mayers Hematoxylin for 2 minutes and placed in cold running tap water for 20 minutes. Dehydration was performed by 2×3 minutes incubation in 70%, 96% and 99% ethanol followed by 3×5 minutes in xylene before coverslips were finally mounted using Entellan (Merck).
RNA extraction and cDNA synthesis
RNA was extracted from (i) murine breast cancer tissue (BCT), (ii) normal murine breast tissue (BT) and (iii) breast tissue of mice exposed to tECM (tECM-BT) using RNAqueous kit (Ambion) according to the manufacturer’s instructions and reverse transcribed using iScriptTM cDNA synthesis kit (Biorad). Total RNA was standardized among the samples by concentration of 1μg/μl prior to reverse transcription and subsequently used for PCR.
Mouse breast cancer PCR array
Copy DNA of BCT, BT and tECM-BT were analyzed using the RT² Profilertrademark PCR Array System for murine breast cancer (SABiosciences). All steps were performed according to the manufacturer’s instructions.
Results
Tumor associated extracellular matrix
The inoculation of 4T1 cells into the 4th mammary fat pad of BALB/c mice (Fig. 1a) resulted in a fast growing mammary tumor with a tendency to metastasize early [13]. Tumor of around 10 mm established after 14 days (Fig. 1b) in all mice and tumors were removed (Fig. 1c and d). The excised tumor tissue was decellularized (Fig. 1e). A small proportion from each decellularized tumor was stained with H&E, Masson’s trichrome and actin in order to validate successful decellularization (Fig. 2).

The inoculation of 4T1 cells into the 4th mammary fat pad of BALB/c mice (a) resulted in a tumor of around 10 mm (b) which was excised (c, d) and subsequently decellularized (e).
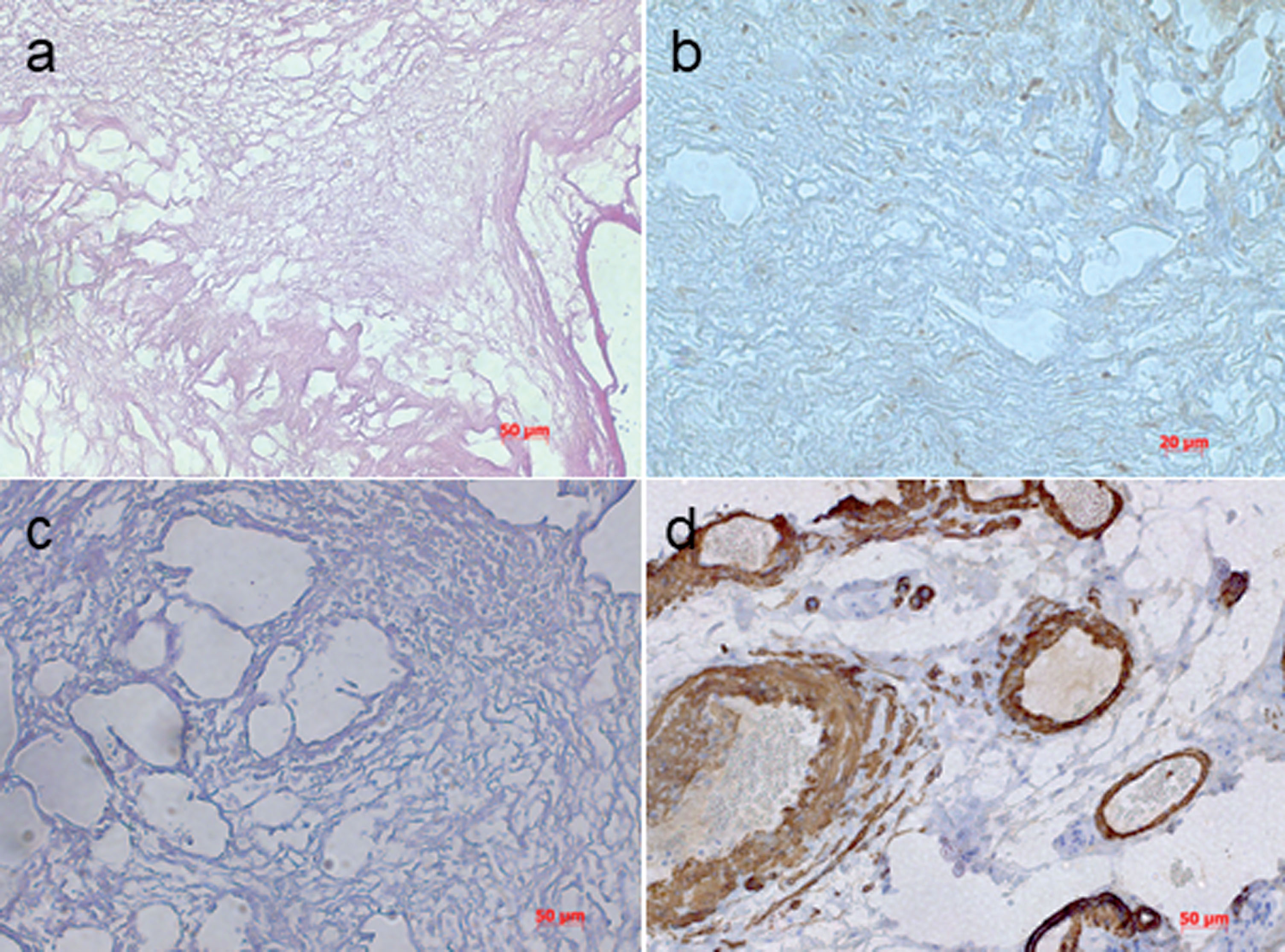
A small proportion of each decellularized tumor tissue was stained for H&E (a) which confirmed no remaining cell nuclei. Furthermore, Masson’s trichrome staining was performed to evaluate collagen in the decellularized tissue (c). In addition, immunohistochemistry for actin showed no positive signal (b) confirming a successful process of decellularization. A positive control for actin (d) confirmed quality of used antibody. In summary, decellularized murine mammary carcinoma showed no evidence of cell nuclei.
All H&E-stained sections as well as Masson’s trichrome and actin staining showed satisfactory morphology under light microscopy without appearance of any remaining nuclei.
Gene array analysis of 84 breast cancer-specific transcripts revealed that the mammary gland cells which were exposed to tumor ECM (tECM-BT) showed a similarly high overexpression for 22 genes as apparent for breast cancer tissue (BCT). The control group with normal mammary gland tissue (BT) showed no expression of breast cancer-specific genes (Fig. 3). The corresponding scatter plot (B) also shows that there is a high agreement in the expression of the examined genes between the mammary gland cells which were exposed to tumor ECM and the breast cancer tissue.
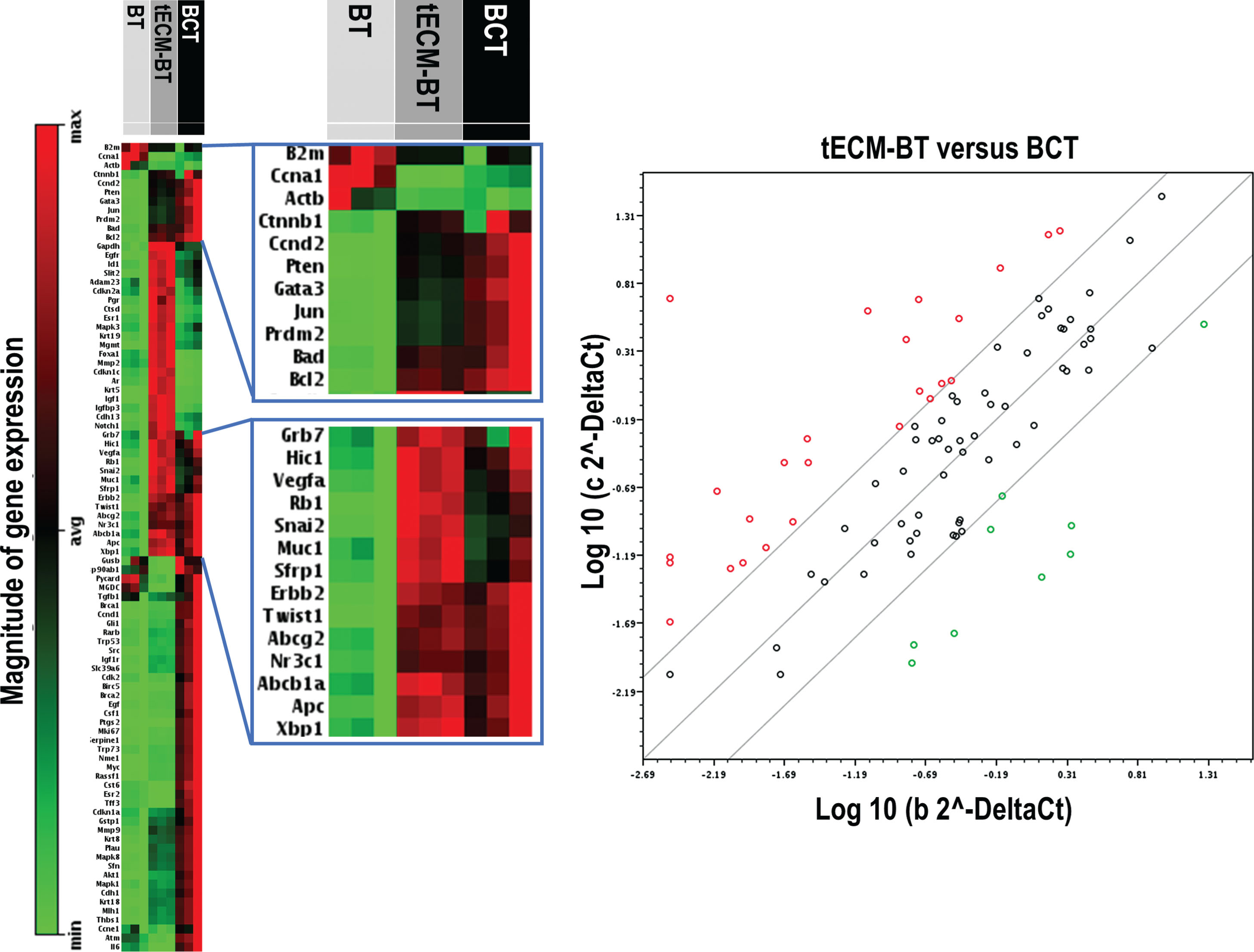
Gene array analysis of 84 breast cancer-specific transcripts compared between normal breast tissue (BT), tumor extracellular matrix breast tissue (tECM-BT) and breast cancer tissue (BCT). The corresponding scatter plot shows that there is a high agreement in the expression of the examined genes between the mammary gland cells which were exposed to tumor ECM and the breast cancer tissue.
Noteworthy, similar expression levels in BCT and tECM-BT were apparent for TWIST playing an essential role in cancer metastasis and acts as an oncogene in several cancers.
Gene expression analysis revealed that an expression shift occurs in breast cells due to contact to tumor extra-cellular matrix. These data underlines that the acellular component of the tumor (ECM) can lead to a transformation of mammary gland tissue cells. These data show for the first time that the interaction of normal breast tissue cells with tumor ECM leads to an exchange of information and a consecutive overexpression of tumor-specific genes.
There is a multitude of evidence that the different components of the microenvironment especially the matricellular proteins [14] can promote tumor growth and metastasis but the question remains if the microenvironment can promote tumor development. We therefore established a mouse model where developing breast epithelial cells grow in extracellular matrix derived from breast cancer tissue. The mammary gland of mice develops from a single duct extending from the nipple during the first 3 weeks postpartum. With the onset of ovarian function after 3-4 weeks of life the rate of ductal growth and branching increases, leading to highly arborated ductal system filling approximately two-thirds of the surrounding fat pad in the 5th week [15]. The formation of the ductal system is dependent on a highly complex interaction between the epithelial cells and the surrounding stroma [16]. During development the composition of the ECM is tightly regulated by controlling the activity of ECM remodeling enzymes at multiple levels [17, 18]. Importantly, ECM-dynamics are reciprocal. On the one hand epithelial and stromal cells are producing remodeling enzymes causing a rearrangement of the ECM, on the other hand ECM with its special chemical and mechanical characteristics influences adjacent cells and modifies their behaviors by regulating their expression patterns [19].
The present data clearly demonstrates that the microenvironment can cause a genetically different behavior of the epithelial cells since only extracellular matrix without any cellular component was implanted.
Based on the available data, a targeted modification of tumor ECM (e.g., antibody therapy) could prove beneficial in combating malignant diseases of the mammary gland tissue. Furthermore, the present data highlights that extracellular matrix of tumor might be one reason for local tumor recurrence when not excised in surgery.