
Abstract
Endothelialization of cardiovascular implants is regarded as a promising strategy for long-term compatibility. While umbilical vein endothelial cells are typically applied in research, human arterial endothelial cells (HAEC) from elderly donors would be the obvious source for autologous cellularization strategies.
In our approach, HAEC from 16 donors of varying age (16–63 years) were divided into two groups (<30 years and >30 years) and analyzed regarding morphology, viability, proliferation, function and senescence status.
No age-related differences were found regarding morphology, viability, density, prostacyclin and nitrite secretion or collagen and laminin production. However, the metabolic activity was slightly decreased (p = 0.0374) and the membrane integrity marginally impaired (p = 0.0404) in cells from older donors. Two out of three senescence assays detected more senescence markers in cells from older donors.
According to the assays applied here, HAEC from young and elderly donors up to the age of 63 years could be judged equally suitable for autologous cellularization strategies. However, this finding should be regarded with caution due to the extremely large variability between individual donors. Further studies comprising a larger sample size are necessary to investigate this issue more thoroughly.
Introduction
Atherosclerosis and resulting cardiovascular diseases (CVD) are the world’s number one cause of death [1]. CVD are characterized by stenosis of vessels restricting blood flow and thus oxygen and nutrient supply to adjacent tissues. In severe cases, total artery occlusion leads to death of the patient. Such advanced stages of CVD require vascular surgery to re-canalize, replace or bypass occluded arteries using vascular grafts. The gold standard for vascular grafts are autologous vessels due to their naturally high compatibility [2, 3]. However, in many cases their availability is limited due to varicosis, atherosclerosis, previous amputation or harvest for previous procedures in up to 30% of the patients [4]. In these cases, prosthetic vascular grafts are implanted as substitutes. These are manufactured from polymers such as poly(ethylene terephthalate) or expanded poly(tetra fluoroethylene) (ePTFE). They exhibit a high stability and blood and soft tissue compatibility, are easy to handle for surgeons and have a good shelf-life stability. However, in ePTFE femoropopliteal bypasses above the knee, patency rates of 39% after 5 years are significantly lower than patency rates of 74% using vein bypasses [2]. This failure rate is mainly due to insufficient endothelialization of the graft leading to thrombosis. Thus, hybrid grafts composed of both a polymer and endothelial cells are considered as a promising alternative with the potential to prevent thrombosis. Multiple studies working on cellularization strategies have been performed (reviewed in [5]). In most of these in vitro studies, human umbilical vein endothelial cells (HUVEC) were used, given their commercial availability or their relatively simple isolation procedure. In addition, they are functionally intact and thus in an anti-thrombotic state and exhibit a high proliferation capacity due to their low biological age. However, these cells are of no relevance for clinical application in cellularized vascular grafts due to their allogeneic origin, which is likely to evoke immunological reactions after transplantation. In fact, the usage of autologous endothelial cells from the respective patient would be desirable. However, the isolation of endothelial cells would require the removal of an intact vessel or large amounts of blood to isolate endothelial progenitor cells. Moreover, a large population of patients requiring vascular grafts are typically older than 70 years [6], which raises the question whether endothelial cells from elderly people are comparable to cells from young people in morphology and function. Moreover, it is unclear whether endothelial cell aging impedes autologous cellularization strategies. Cell aging occurs not only in vitro (reviewed in [7]) but also in vivo (reviewed in [8]). The World Health Organization defines aging as a natural, inevitable and multifactorial process that occurs within an organism at genetic, molecular, cellular, organ and system levels [9]. Cell aging is often accompanied by cell dysfunction, which in case of endothelial cells may lead to disturbances in blood flow and vascular hemostasis [10, 11]. Endothelial cells that have undergone replicative senescence in culture show unique changes in morphology such as an increase in size, flattening, polymorphic nuclei and vacuolization [12, 13]. In addition, they have a reduced proliferation potential and generate less of the key vasodilator and antithrombotic molecules nitric oxide and prostacyclin. In contrast, the production of reactive oxygen species, plasminogen activator inhibitor-1, inflammatory mediators, lysosomal enzymes such as senescence-associated β-galactosidase (β-gal) and matrix degrading enzymes is increased (reviewed in [8]). On the molecular level, senescence is characterized by telomerase uncapping and shortening [14, 15] explaining the decreased proliferation potential of affected cells. In particular, telomere shortening to a critical threshold triggers the expression of the cyclin-dependent kinase inhibitor (CDKI) p16 and activates p53, which is known as “guardian of the genome” [16]. p53 transactivates expression of CDKI p21, which in concert with CDKI p16 arrest the cell cycle by preventing G1/S progression (reviewed in [17]). In addition, the proliferation marker Ki67 is downregulated in senescent cells [18]. These changes indicate that senescent endothelial cells change from their anti-thrombotic phenotype towards a pro-inflammatory and pro-thrombotic state [19, 20]. When considering cellularization of vascular grafts, it is essential to exclude that endothelial cells taken from a specific donor are senescent, as this would provoke a rapid occlusion of the graft.
Here we hypothesized that there are age-dependent differences in function of arterial endothelial cells but also morphological changes might be indicative for differences in the state of senescence. To confirm this hypothesis, we analyzed arterial endothelial cells from 16 different donors of varying age with regard to morphology and function as well as their state of senescence, providing a framework to evaluate their potential as therapeutically relevant cell sources.
Materials and methods
Study design
Human arterial endothelial cells (HAEC) from 16 donors of different ages (16-63 years; 3 female, 13 male) were cultivated for 6 days. At day 2, day 4 and day 6 of cultivation cells were analyzed regarding cell morphology (actin staining), viability, density, the secretion of prostacyclin and nitrite and expression of von Willebrand factor. In addition, the capacity for the synthesis of extracellular matrix proteins such as collagen and laminin over time was investigated, as endothelial cells gradually build up an extracellular matrix after adherence in order to anchor themselves firmly to the substrate. Moreover, the metabolic activity and membrane integrity as well as the state of senescence was assessed using three different assays. For an age-related comparison, donors were divided into two groups (< 30 years: n = 6; mean age 22±4 years and > 30 years: n = 10; mean age 50±11).
Cell culture
HAEC (provided by PeloBiotech, Planegg/Martinsried, Germany) were cultivated in endothelial cell culture medium supplemented with 5% (v/v) fetal bovine serum (upcyte technologies GmbH, Hamburg, Germany) in a standard humidified incubator at 37°C with 5% (v/v) CO2. For the assessment of cell viability, density, fluorescence staining and quantification of prostacyclin and nitrite, cells were seeded on glass coverslips (Th. Geyer GmbH, Hamburg, Germany) in 24-well plates (TPP, Techno Plastic Products AG, Trasadingen, Switzerland) with a cell density of 15,000 cells/well for 2 days, 4 days and 6 days. The metabolic activity and membrane integrity were assessed in 96-well plates (Greiner Bio One, Leipzig, Germany) with a cell density of 5,000 cells/well after 2 days of cultivation. For determining the state of senescence cells were seeded in 24-well plates with 11,000 cells/well for 3 days. Cells were used at passages 4-9 for these experiments [21].
Cell viability
The analysis of the cell viability was performed using fluorescein diacetate (FDA, 12.5 μg·mL-1, Invitrogen, Carlsbad, CA, USA) to stain vital cells in green and propidium iodide (PI, 1 μg·mL-1, Sigma-Aldrich, Taufkirchen, Germany) to stain dead cells in red. For this, FDA and PI were added to the cell culture medium and images of HAEC were immediately taken in a 20-fold primary magnification (Laser scanning microscope Axiovert 200M, Zeiss, Oberkochen, Germany). The numbers of live and dead cells were counted using ImageJ and expressed in percent of the total cell number. The number of vital adherent cells was referred to the growth area in mm2 (sample size n = 9 (3 wells per donor with 3 images taken per well)).
Senescence assays
(a) Semi-quantitative determination of β-Galactosidase activity
To qualitatively assess the β-galactosidase activity, the Senescence β-Galactosidase Staining Kit (Cell Signaling Technology, Frankfurt am Main, Germany) was used according to the manufacturer’s instructions. Briefly, HAEC were fixed with fixative solution and incubated overnight at 37 °C in staining solution containing a substrate for β-galactosidase. To evaluate the staining in a semi-quantitative manner, cell nuclei were stained using 4′,6-diamidino-2-phenylindole (DAPI, Roth, Karlsruhe, Germany, 1:50) to detect all cells. For analysis, 5 images were taken per well in 10-fold primary magnification (Axio Imager.Z2m, Zeiss, Oberkochen, Germany) and the amount of β-galactosidase-positive cells was expressed as percent of the total cell number (sample size n ≥ 630 (3 wells per donor with at least 70 images taken per well in three independent experiments)).
(b) Quantification of β-galactosidase activity
Quantification of the senescence-associated β-galactosidase activity was performed using the Cellular Senescence Activity Assay (Enzo Life Sciences, Lörrach, Germany) according to the manufacturer’s instructions. Briefly, HAEC were lysed and the cell lysate was incubated with a fluorometric substrate for β-galactosidase. After 1 hour incubation at 37°C, the activity of β-galactosidase was measured at 360 nm (Excitation) and 465 nm (Emission) using a photometer (Tecan infinite M200 pro, Crailsheim, Germany). To compensate for differences in cell numbers between wells, the amount of β-galactosidase was normalized to the amount of protein per well. For this, the Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Berlin, Germany) was used according to the manufacturer’s instructions. Briefly, 10 μL of bovine serum albumin standard or sample was added per well to a 96-well plate (Greiner Bio One, Leipzig, Germany). BCA Reagents A and B were mixed in a ratio of 50 : 1 and 200 μL of this mixture was added per well. After 30 min incubation at 37°C the absorbance was measured at 562 nm using a photometer (Tecan infinite M200 pro, Crailsheim, Germany). Protein concentrations per well were calculated based on a standard curve and used for normalization of the β-galactosidase activity (sample size n = 18 (3 wells per donor measured in duplicates in three independent experiments)).
(c) Fluorescence staining of p16INK4A, p21 and Ki67
To visualize senescence markers p16INK4A (hereinafter called p16) and p21 as well as the proliferation marker Ki67 an immunocytological staining was performed. Briefly, HAEC were fixed with 4% (w/v) paraformaldehyde (Sigma-Aldrich, Taufkirchen, Germany) for 30 min, permeabilized using 0.5% (v/v) Triton-X-100 for 10 min (Sigma-Aldrich, Taufkirchen, Germany) and unspecific binding sites were blocked with 5% (w/v) bovine serum albumin (Roth, Karlsruhe, Germany) for 20 min at room temperature. As primary antibodies mouse anti-human p16 (Abcam, Cambridge, United Kingdom, ab54210, 1 : 500), rabbit anti-human p21 (Abcam, Cambridge, United Kingdom, ab109520, 1 : 1000) and rat anti-human Ki67 (Thermo Fisher Scientific, Berlin, Germany, 14-5698-82, 1 : 200) were incubated for 1 h at room temperature. After three washing steps with phosphate buffered saline (Thermo Fisher Scientific, Berlin, Germany), cells were incubated with the secondary antibodies goat anti-mouse IgG2a conjugated to Alexa Fluor 647 (Abcam, Cambridge, United Kingdom, 1 : 200, for p16), donkey anti-rabbit IgG conjugated to DyLight 550 (Thermo Fisher Scientific, Berlin, Germany, SA5-10039, 1 : 500, for p21), and goat anti-rat IgG conjugated to Alexa Fluor 488 (Thermo Fisher Scientific, Berlin, Germany, 1 : 200, for Ki67), respectively, as well as 4′,6-diamidino-2-phenylindole (DAPI, Roth, Karlsruhe, Germany, 1 : 50) to counterstain cell nuclei for 1 h at room temperature in the dark. Finally, cells were washed and covered with mowiol® 4-88 as mounting medium (Roth, Karlsruhe, Germany). For analysis, 5 images were taken per well in 20-fold primary magnification (confocal laser scanning microscope DMi8, Leica, Wetzlar, Germany) and the amount of stained cells was expressed as percent of the total cell number (sample size n = 45 (3 wells per donor with 5 images taken per well in three independent experiments)).
Metabolic activity
The mitochondrial activity was measured using the MTS cell Titer 96 Aqueous Non-radioactive Cell Proliferation Assay (MTS assay, Promega, Mannheim, Germany) according to the manufacturer’s instructions. Briefly, 20 μL of MTS-mixture was added per well and incubated for 3 hours at 37°C before the absorbance was measured at 492 nm using a photometer (Tecan infinite M200 pro, Crailsheim, Germany). The metabolic activity is presented as absorbance at 492 nm (sample size n = 48 for donors < 30 years (3 donors measured in 8 replicates in 2 independent experiments) and n = 48 for donors > 30 years (6 donors measured in 8 replicates in 1 experiment)).
Cell membrane integrity
Cell membrane integrity was measured using the LDH Cytotoxicity Assay Kit II (LDH, Roche, Grenzach, Germany) according to the manufacturer’s instructions. Briefly, 10 μL of cell culture supernatant were added to 100 μL of LDH reaction mix. After 30 min incubation at room temperature in the dark the absorbance at 450 nm (reference wavelength: 650 nm) was measured using a photometer (Tecan infinite M200 pro, Crailsheim, Germany). The cell membrane integrity is presented as absorbance at 450-650 nm (sample size n = 48 for donors < 30 years (3 donors measured in 8 replicates in 2 independent experiments) and n = 56 for donors > 30 years (7 donors measured in 8 replicates in 1 experiment)).
Quantification of prostacyclin
Prostacyclin secretion was quantified using a competitive enzyme immunoassay (6-keto Prostaglandin F1α ELISA Kit, Cayman Chemical, Hamburg, Germany) according to the manufacturer’s instructions. Briefly, 50 μL cell culture supernatant were used per well and measurements were performed in triplicates. Absorbance was recorded at 410 nm using a photometer (Tecan infinite M200 pro, Crailsheim, Germany). Prostacyclin concentrations were calculated based on a standard curve and normalized to the number of adherent cells per well (n = 3; 1 experiment measured in triplicates whereas not all measurements were analyzable leading to varying numbers of data points).
Quantification of nitrite
Nitrite secretion was quantified using a commercial kit (Total nitric oxide and nitrate/nitrite Kit, R&D Systems, Wiesbaden, Germany) according to the manufacturer’s instructions. Briefly, nitrate was enzymatically converted to nitrite by nitrate reductase and total nitrite was colorimetrically detected as an azo dye product of the Griess reaction. In particular, 50 μL cell culture supernatant were used per well and measurements were performed in triplicates. Absorbance was recorded at 555 nm using a photometer (Tecan infinite M200 pro, Crailsheim, Germany). Nitrite concentrations were calculated based on a standard curve and normalized to the number of adherent cells per well (n = 3; 1 experiment measured in triplicates).
Fluorescence staining of actin filaments, von Willebrand factor, collagen and laminin
To assess the cell morphology qualitatively, cells were stained with phalloidin conjugated to Alexa Fluor 555 (Thermo Fisher Scientific, Berlin, Germany, 1 : 40) to visualize actin filaments and antibodies to detect von Willebrand factor, collagen I-V and laminin (after permeabilization of the cells) as described above. The primary antibodies were rabbit anti-human von Willebrand factor (Sigma-Aldrich, Taufkirchen, Germany, F3520, 1 : 200), rabbit anti-human collagen I-V (Abcam, Cambridge, United Kingdom, ab36064, 1 : 250) and mouse anti-human laminin (antibodies-online GmbH, Aachen, Germany, ABIN269898, 1 : 250). As secondary antibodies donkey anti-rabbit antibody conjugated to Alexa Fluor 488 (Invitrogen, Carlsbad, CA, USA, A21206, 1 : 500, for von Willebrand factor), donkey anti-rabbit IgG conjugated to DyLight 550 (Thermo Fisher Scientific, Berlin, Germany, SA5-10039, 1 : 500, for collagen), and donkey anti-mouse IgG conjugated to Alexa Fluor 488 (Jackson ImmunoResearch, Cambridge, United Kingdom, 715-545-150, 1 : 500, for laminin) were used.
Statistics
Statistical analyses were performed using Graphpad Prism 6 (Graphpad Software, San Diego, California). Gaussian distribution of the data was tested using the d’Agostino & Pearson omnibus normality test. Data are reported as arithmetic mean±standard error of the mean. Comparisons between two groups were performed by Student’s t-test for normally distributed data and by Mann-Whitney test for non-parametric data. Differences were considered significant at p < 0.05.
Results
Morphology, viability and proliferation of HAEC
Actin-and von Willebrand factor staining
Endothelial cells from all donors exhibited a cobblestone-like morphology typical for endothelial cells [22]. The pattern of actin fibers was similar in cells from younger and older donors. Whereas actin fibers were located in central parts of the cells at day 2 and day 4, they translocated to the outer rim of many cells at day 6 indicating that a functionally-confluent monolayer of cells started to develop at this point of time. Von Willebrand factor as an identity marker for endothelial cells was present in nearly all of the cells regardless of the donor age. This indicates that only few contaminating cells such as fibroblasts or smooth muscle cells were present in the cell population. Representative images are shown in Fig. 1.

Endothelial cell morphology and identity. Arterial endothelial cells from 16 different donors were cultivated and analyzed at day 2, day 4 and day 6 of cultivation regarding their morphology by actin filament staining (A-C) and identity as endothelial cells by von Willebrand Factor staining (D-F). Cell nuclei were stained blue using DAPI. Only one set of representative images is shown given that no age-related morphological and identity-related differences were obvious between donors. Images were taken using a confocal laser scanning microscope with a 20-fold primary magnification.
The majority of endothelial cells from all donors remained viable over the complete cultivation time of 6 days as shown by FDA-staining (Fig. 2). At day 2, all cells from younger donors and nearly all cells from older donors were viable. Up to day 6 of cultivation, the viability significantly decreased to 93±6% in the group with donors younger than 30 years (p = 0.0031) and 95±3% in the group with donors older than 30 years (p = 0.0186). With regard to the donor age, no significant difference between the two age groups were observed (p > 0.9999).
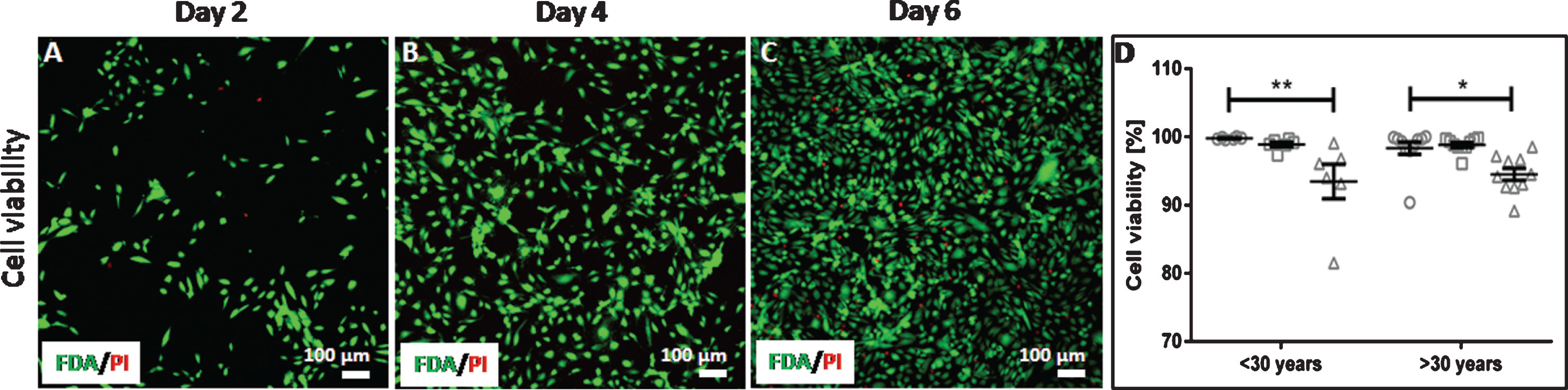
Endothelial cell viability. Arterial endothelial cells from 16 different donors were cultivated and analyzed at day 2, day 4 and day 6 of cultivation regarding their viability. Viable (green) and dead (red) cells were imaged using a confocal laser scanning microscope with a 20-fold primary magnification (A–C). Viability of cells from donors < 30 years and > 30 years was expressed as percent of viable cells (D) shown as grouped scatter plot with the arithmetic mean±standard error of the mean (n = 9; 1 experiment with 3 wells per donor and 3 images taken per well). Symbols represent the cell viability at different time points of cultivation: circles represent day 2, squares represent day 4 and triangles represent day 6.*: p < 0.05; **: p > 0.01.
The increasing cell density over the cultivation time of 6 days shows that cells were able to adhere and proliferate on the glass cover slips. No statistical difference between the two age groups could be detected (at all time points: p > 0.9999). Nevertheless, it is noteworthy that the cell density in the group with cells from donors older than 30 years is 1.7-fold higher at day 6 than in the group with cells from donors younger than 30 years (p > 0.9999). Moreover, the increase in the cell density from day 2 to day 6 was statistically significant with cells from older donors (p = 0.003). In contrast, the increase in cell density with cells from younger donors was not significant (p = 0.422; Fig. 3).

Endothelial cell density. Arterial endothelial cells from 16 different donors were cultivated and analyzed at day 2, day 4 and day 6 of cultivation regarding their cell density expressed as number of adherent cells per mm2. Donors were divided into two age groups (< 30 years and > 30 years) for an age-related comparison. Data are shown as grouped scatter plot with the arithmetic mean±standard error of the mean (n = 9; 1 experiment with 3 wells per donor and 3 images taken per well). Symbols represent the cell density at different time points of cultivation: circles represent day 2, squares represent day 4 and triangles represent day 6. **: p > 0.01.
The state of cellular senescence can be determined by different methods. Here, three methods were applied: (i) semi-quantitative determination of the β-galactosidase activity by counting of β-galactosidase-positive cells in relation to the total cell number, (ii) quantification of the senescence-associated β-galactosidase activity in the cell lysate, and (iii) immunocytological staining and counting of the senescent markers p16 and p21 in relation to the total cell number.
According to the β-galactosidase activity-staining assay (method (i), Fig. 4A), some more cells from older donors expressed β-galactosidase (40% ± 0.3%) compared to cells from younger donors (37% ± 0.3%; p < 0.0001). In contrast, based on the quantification of the β-galactosidase activity in the cell lysate (method (ii), Fig. 4B), the senescence-associated enzyme was more expressed in cells from younger donors (4664±1522 RFU* μg-1) than in older donors (3666±2786 RFU* μg-1; p = 0.003). Immunocytological staining of p16 and p21 (method (iii), Fig. 4 C) showed that these two senescence markers were expressed in more cells from older donors (p16 : 19% ± 1.8% and p21 : 28% ± 1.8%) compared to cells from younger donors (p16 : 14% ± 2.1% and p21 : 21% ± 2.5%). In case of p21, this difference was statistically significant (p = 0.026) whereas it was not significant for p16 (p = 0.1976). It is noteworthy that more cells from older donors expressed Ki67 as a marker for cell proliferation (35% ± 1.9%) than cells from younger donors (30% ± 2.6%). However, this was not statistically significant (p = 0.0741).

Endothelial cell senescence status. Arterial endothelial cells from 16 different donors were cultivated for three days and analyzed regarding their senescence status using three different types of assays. β-Galactosidase activity was either qualitatively stained followed by a semi-quantitative analysis of the amount of positively stained cells in relation to the total cell number (A) or quantified in the cell lysate and normalized to the protein content (B). In addition, the senescence markers p16 and p21 as well as the proliferation marker Ki67 were stained by immunocytochemistry (C). Donors were divided into two age groups (< 30 years and > 30 years) for an age-related comparison. Data are shown as scatter plot with the arithmetic mean±standard error of the mean. For A: n≥630; 3 independent experiments with 3 wells per donor and at least 70 images taken per well. For B: n = 18; 3 independent experiments with 3 wells per donor and measurements in duplicates. For C: n = 45; 3 independent experiments with 3 wells per donor and 5 images taken per well. Symbols represent the expression of p16 (circles), p21 (squares) and Ki67 (triangles).*: p < 0.05; **: p > 0.01; #: p < 0.0001.
Metabolic activity and cell membrane integrity
The metabolic activity was slightly but significantly reduced in cells from older donors (0.89±0.02) compared to cells from young donors (0.95±0.02; p = 0.0374; Fig. 5A). In accordance with this, the cell membrane integrity was more impaired in cells from older donors (0.043±0.002) compared to cells from younger donors (0.036±0.002; p = 0.0404; Fig. 5B).
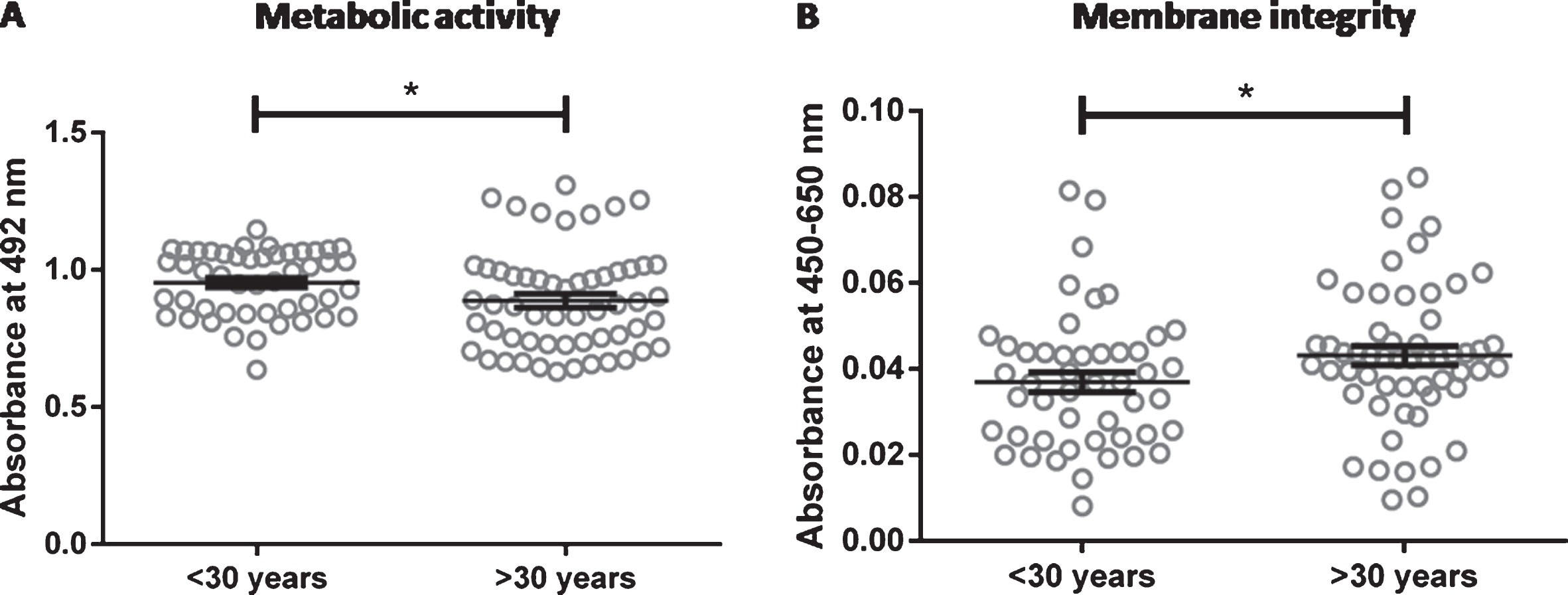
Metabolic activity and membrane integrity of endothelial cells. Arterial endothelial cells from 16 different donors were cultivated for two days and analyzed for metabolic activity (A) and membrane integrity (B). Donors were divided into two age groups (< 30 years and > 30 years) for an age-related comparison. Data are shown as scatter plot with the arithmetic mean±standard error of the mean (n = 48 for donors < 30 years (3 donors measured in 8 replicates in 2 independent experiments) and n = 48 for donors > 30 years (6 donors measured in 8 replicates in 1 experiment). *: p < 0.05.
In contrast to the parameters described above, no age-dependent differences in the secretion of prostacyclin or nitrite were detected (at all time points: p > 0.9999). The prostacyclin concentration decreased in both groups over the cultivation time of 6 days to a similar extent (Fig. 6A). Likewise, the amount of total nitrite decreased over time in cells from donors younger than 30 years. In the older group, the nitrite concentration was highest at day 4 (Fig. 6B). However, the differences between individual donors were extremely high resulting in large standard deviations and thus in no statistically significant differences between the two age groups (at all time points: p > 0.9999).
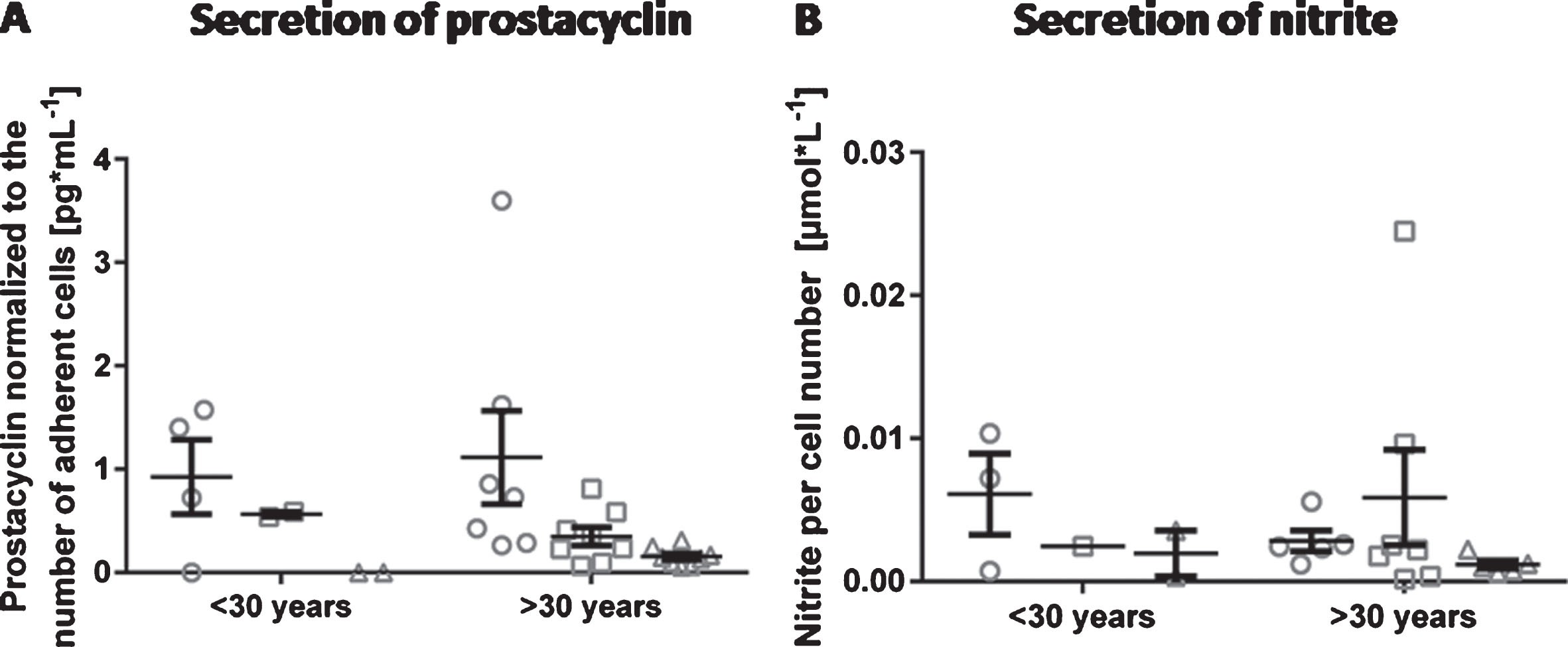
Secretion of prostacyclin and nitrite by endothelial cells. Arterial endothelial cells from 16 different donors were cultivated and analyzed at day 2, day 4 and day 6 of cultivation regarding the release of prostacyclin (A) and nitrite (B) into the cell culture medium. Donors were divided into two age groups (< 30 years and > 30 years) for an age-related comparison. Data are shown as grouped scatter plot with the arithmetic mean±standard error of the mean (n = 3; 1 experiment measured in triplicates whereas not all measurements were analyzable leading to varying numbers of data points).
Endothelial cells from all donors synthesized collagen at day 2 (Fig. 7A). Secretion of collagen into the extracellular environment was observed at day 4 or day 6 in a donor-dependent manner (Fig. 7B1 and B2 as well as C1 and C2). However, this secretion was not associated to the donor age. Laminin was detected in endothelial cells from all donors regardless of the age. In contrast to collagen, it was intracellularly located at all time points of cultivation and not secreted into the extracellular space (Fig. 7D-F).
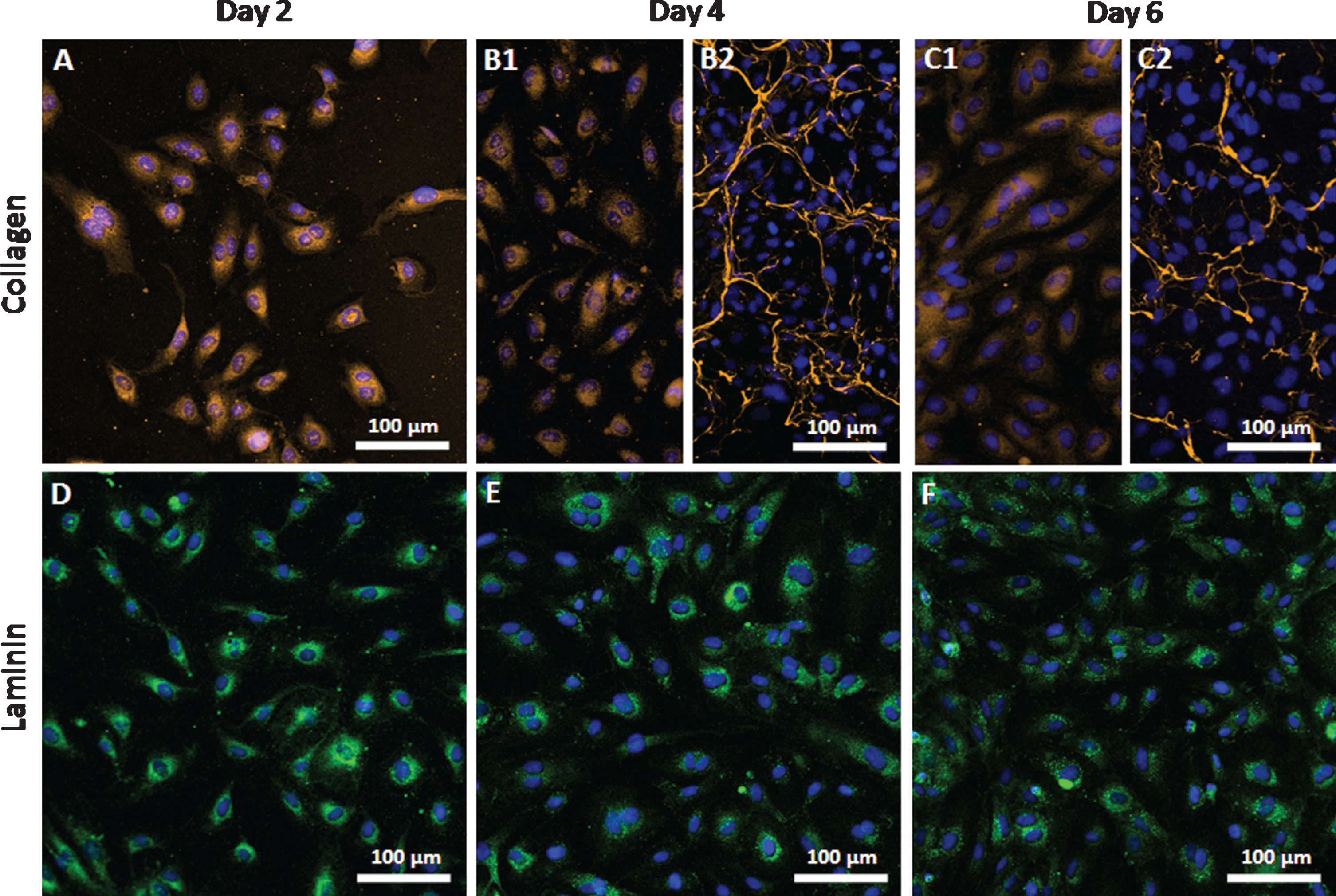
Expression of collagen and laminin by endothelial cells. Arterial endothelial cells from 16 different donors were cultivated and analyzed at day 2, day 4 and day 6 of cultivation regarding the expression of collagen (A–C) and laminin (D–F). Cell nuclei were stained blue using DAPI. Since no age-related differences were found between donors representative images are shown. B1 and C1 show cells expressing intracellular collagen and B2 and C2 show cells secreting collagen into the extracellular space. Images were taken using a confocal laser scanning microscope with a 20-fold primary magnification.
HUVEC are the most common type of endothelial cells used in cellular research. However, from a translational point of view, they are of no relevance in terms of autologous cellularization strategies for vascular implants. In contrast, arterial endothelial cells from elderly people should be investigated more thoroughly as they represent the key cell type for cellularization of vascular grafts. We therefore analyzed arterial endothelial cells from 16 different donors of varying age with regard to morphology and function as well as their state of senescence. The major finding was that extremely large variations between cells from individual donors exist and that far more donors have to be analyzed to draw reliable conclusions. The data presented here should thus be regarded as preliminary, which need to be confirmed on a broader basis.
With regard to morphology, no age-dependent difference in the expression of stress fibers as well as in the decline of these fibers during the development of a functionally-confluent monolayer was observed. Thus, cells from all donors are likely to prevent platelet adhesion in order to maintain an undisturbed blood flow. Likewise, the expression of von Willebrand factor, a unique marker of EC, was similarly expressed in both age groups. However, the differences in the viability of cells from individual donors was not statistically different. Also, the cell density at day 6 was significantly larger with cells from older donors than with cells from young donors suggesting that cells from older donors have an increased proliferation potential. Although this finding contradicts the observation that young cells of various cell types possess a higher proliferation potential compared to older cells [23–25], it is in line with the increased expression of the proliferation marker Ki67 in cells from older donors (Fig. 4 C). Ki67 was first identified in Hodgkin lymphoma cell nuclei in 1983. While it is a widely used clinical marker for proliferation of cancer cells and prognosis of cancer progression [26, 27], recent research brings up increasing doubts on its reliability. First of all, it was found that not every cell type expresses Ki67 during proliferation [28, 29]. Secondly, the amount of Ki67 expressed after entry into the cell cycle (G1/S-phase) depends on the time the cells have spent in G0-phase (quiescence). The longer the time cells have spent in G0 the lower the expression of Ki67 after re-entry into the cell cycle [30]. This indicates that not only the presence or absence of Ki67 but also quantitative information should be considered in order to make a profound prognosis. Thirdly, in some cancer cases such as node-positive breast cancer, Ki67 was found to be no valid marker at all [31]. We are not aware that such controversies exist for endothelial cells cultured in vitro. In contrast, several studies report on the relation between proliferation and Ki67 expression [32, 33]. Thus, the data presented here showing that cell density corresponds to the expression of Ki67 indicates that in vitro proliferation of the cells from these specific donors older than 30 years is indeed increased compared to cells from these specific younger donors. Nevertheless, more donors should be analyzed to confirm this result, taking into consideration that only two out of ten donors in the group older than 30 years were older than 60 years.
To determine the senescence status of the cells, three different types of assays were used. The most common assay is based on the detection of the β-galactosidase activity via staining for enzymatic activity using chromogenic substrates [34]. Here, this assay detected some more senescent cells in the group with older donors (40% ± 0.3%) compared to cells from younger donors (37% ± 0.3%; Fig. 4A), which fits to the assumption that senescent cells increase in number with increasing age. Although this difference was statistically significant it was small and thus raises the question whether it is biologically relevant. Moreover, this assay is associated with a number of concerns. The staining intensity varies considerably between cells making it difficult to distinguish faint from clearly positive cells. Thus, counting β-galactosidase-positive cells is subjective and semi-quantitative. To circumvent this issue, here an ImageJ-based semi-automatic method developed by Lozano-Gerona and Garcia-Otin was used [35]. Moreover, a high confluence leading to contact inhibition and quiescence was shown to upregulate β-galactosidase activity regardless of the senescence status leading to false-positive results when confluent areas were used for counting. Thus, a quantitative assay for β-galactosidase considering all cells was developed [36] and applied in our study. This assay led to contradictory results compared to the β-galactosidase activity-staining-method because the β-galactosidase activity was significantly higher in cells from younger donors rather than from older donors (Fig. 4B). Since β-galactosidase is assumed not to be a unique marker for senescence [37], it is recommended to use additional markers such as p16 and p21 among others [18]. Here, the expression of p16 and p21 was more frequently observed in cells from older donors compared to young donors (Fig. 4C). Thus, two of the three assays applied here have shown a slight tendency towards more senescent cells from older donors whereas one assay points towards the other direction. Generally, it has to be noted again that the observed differences were marginal and despite their statistical significance barely relevant from a biological point of view. Thus, a fundamental study comparing all three assays regarding their sensitivity would be a valuable contribution to better assess such data.
The metabolic activity was slightly but significantly decreased and the cell membrane integrity to some extent impaired in cells from older donors. Both findings are in line with several previous studies. Human corneal endothelial cells from donors older than 50 years exhibited reduced expression of proteins that support metabolism [38] and hyperpermeability was clearly associated with increasing age [39]. Although the observed differences were small, they might be a result of the senescent phenotype of cells from older donors. Thus, it is possible that cells from older donors are compromised in their function to maintain hemostasis. Senescent cells are known to be metabolically different from non-senescent cells and have the potential to contribute to metabolic diseases and cancer [40, 41]. In addition, they are associated with disrupted adherens junctions caused by impaired vascular endothelial cadherin and increased monolayer permeability [42]. Although such markers were not investigated in this study, it can be hypothesized that their expression is to some extent impaired in cells from older donors compared to cells from young donors.
Prostacyclin (PGI2) is a parameter to assess the activation state of endothelial cells. Under physiological conditions, it is expressed at basal levels to prevent activation and adhesion of platelets enabling an undisturbed blood flow [10]. At sites of inflammation or injury, endothelial cells become activated which initiates a cascade of reactions including the increased expression of PGI2 aiming to protect the endothelium from damage and resulting thrombosis. Thus, PGI2 is commonly known as an indicator of damage and a clear sign for the activation/perturbation of endothelial cells [43, 44]. Here, the concentration of PGI2 decreased over time in both age groups. This is in line with the decrease of stress fibers and so to the development of a functionally-confluent monolayer as shown in former studies [45, 46] (Fig. 1A–C). Thus, both parameters fit together and show that endothelial cells cultured in vitro need a certain time to adapt to their environment in order to develop a physiological relevant phenotype.
Nitric oxide is another vasodilating molecule essential for the regulation of vascular tone and adaptation to blood pressure changes [47]. An early study found that the production of nitric oxide is increased in proliferating aortic bovine endothelial cells and decreased in quiescent cells [48]. The same phenomenon was observed in the group with donors younger than 30 years (Fig. 6A). In the older age group, there was not such a linear trend recognizable, however, this is most likely due to the extremely large variability at day 4. The subendothelium on which the endothelial cell monolayer rests is a highly reactive surface and is synthesized by endothelial cells itself [11, 49]. The synthesis and deposition of extracellular matrix proteins are an important function of endothelial cells as they provide structural support and regulatory cues for endothelial cell proliferation, migration, morphogenesis, survival and vessel stabilization thus controlling neovascularization. The basal membrane matrix of vascular endothelial cells is mainly composed of collagen type IV variants and laminin isoforms among other molecules. Surprisingly, little is known about how this matrix is temporally and spatially assembled [50]. In Drosophila and C. elegans laminin assembly is followed by an independent self-assembly of collagen type IV networks [51]. In the present study, laminin was synthesized by cells from all donors but not released into the extracellular space up to day six. In contrast, collagen was deposited by cells from some donors, however, not in an age-dependent manner. From this it could be hypothesized that laminin deposition in vitro requires more time than collagen secretion. A recent study found that only endothelial progenitors were able to produce matrix components in vitro under normoxic conditions whereas mature endothelial cells required hypoxic conditions [52]. This would explain why laminin was not found to be deposited in our study, however, collagen was deposited despite normoxic conditions. Considering all data shown here, which could be partly related to the age of the donors even though the differences were extremely small, it remains unanswered whether differences observed in the collagen secretion are truly age-related or due to natural donor-dependent variations. Apart from that, it is important to keep in mind that the results found in this study refer to donors of a maximum age of 63 years. Clear differences might appear at a later stage of life as was previously shown for platelets from donors of up to 98 years of age [53].
Conclusion
In contrast to the hypothesis, no age-dependent morphological differences concerning the expression of actin fibers were detected between the two age groups tested in this study (22±4 years and 50±11 years). On the other hand, differences regarding endothelial cell functionality and status of senescence, while small, could be measured. These differences were, however, confounded by the high variability between cells from individual donors. Thus, the biological relevance of the observed differences is low. According to the parameters addressed here, it could be concluded that arterial endothelial cells from both young and elderly donors are equally suitable for the cellularization of vascular grafts. However, it is indispensable to conduct more studies comprising a larger sample size but also addressing graft interactions directly, in order to answer this question in a more reliable manner. In addition, cells from seniors (>80 years) should be included in future studies as this age group becomes increasingly important for vascular implants due to the rising life expectancy, especially in the western world.
Footnotes
Acknowledgments
The work was financially supported by the Helmholtz-Association through programme-oriented funding and by the Ministry for Science, Research and Cultural Affairs of Brandenburg through the grant of the joint project “Konsequenzen der altersassoziierten Zell- und Organfunktionen” of the Gesundheitscampus Brandenburg, as well as by the European Fonds of Regional Development (EFRE, Brandenburg, Germany; project “PERsonalisierte Medizin durch FUNCTIONomics in Berlin-Brandenburg: “Endothelfunktionstestung mit Medikamenten und isolierten Metaboliten”; project number: 85002925). The authors thank Dr. Manfred Gossen for his valuable support during manuscript preparation.